
Anatomie 4
b. La racine
La structure primaire de la racine est, contrairement à celle de la tige, relativement uniforme d'une plante à l'autre. Elle est représentée schématiquement par la figure 1
?Les tissus conducteurs forment au centre de la racine un cylindre compact, appelé cylindre central , constitué de faisceaux de xylème et de phloème qui alternent régulièrement . Dans le xylème, et contrairement à ce qui passe dans la structure primaire de la tige, la différenciation des vaisseaux progresse de l'extérieur vers l'intérieur, le protoxylème est externe et les vaisseaux de plus gros calibre sont situés vers le centre de la racine.
?Autour de ces tissus conducteurs se trouvent deux assises cellulaires particulières : la plus interne, appelée péricycle, doit son importance à ce qu'elle est le départ de la formation des racines latérales et des radicelles ; la plus externe , l'endoderme, est caractérisée par la présence sur les parois radiales de ses cellules d'épaississement cutinisés représentés sur la figure 1 C , et qui jouent un rôle important dans le contrôle de l'absorption.
?L'écorce interne présente souvent, mais pas toujours une disposition régulière de ses cellules. L'écorce externe est un parenchyme banal.
Enfin la racine est limitée extérieurement par une assise comparable à un épiderme qui n'aurait pas de cuticule et dont certaines cellules se sont démesurément allongées en constituant des poils absorbants. Il y a lieu de bien remarquer qu'un poil absorbant est constitué d'une seule cellule, et non pas d'une file cellulaire ( fig 1 B). Sur une coupe transversale pratiquée à proximité du sommet de la racine , on voit l'épiderme, non encore différencié en couche pilifère, et doublé extérieurement par les tissus de la coiffe.
C Anatomie de la feuille
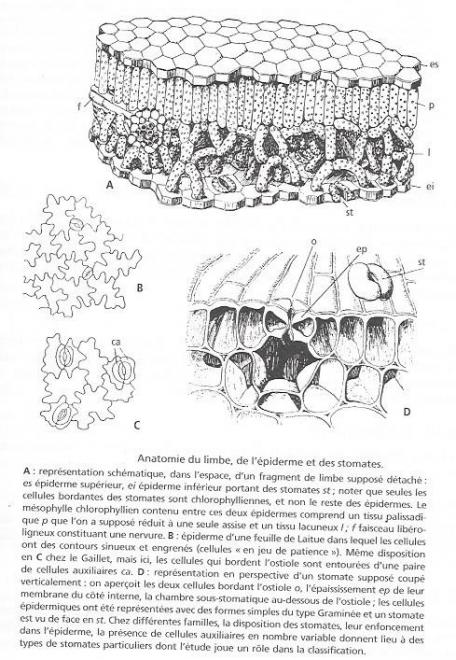
L'anatomie du limbe est présentée à la figure 2
Une coupe transversale dans le limbe ( fig 2 A) montre un épiderme supérieur et un épiderme inférieur dépourvu de chloroplastes et un tissus médian, appelé mésophylle, formé de cellules chlorophyliennes. Le mésophylle est ordinairement différencié en deux couches : la partie supérieure est constituée d'une ou plusieur assises de cellules allongées dans le sens vertical, de sorte qu'elles apparaissent en coupe comme les planches d'une palissade et, pour cette raison, ce tissus est appelé tissus palissadique ; il est risque en chloroplastes. La partie inférieure est toute différente , ses cellules laissent entre elles de larges lacunes et elles sont plus pauvres en chlorophylle : c'est le tissus lacuneux . La photosynthèse s'effectue essentiellement dans le tissus palissadique qui possède le plus de pigment assimilateur et qui reçoit la lumière frappant la face supérieure de la feuille ; les échanges gazeux entre le tissus assimilateur et l'extérieur, et notamment l'arrivée du gaz carbonique se font par circulation de l'air à travers le tissus lacuneux et l'épiderme inférieur. Ce dernier est interrompu par de nombreux orifices appelés stomates et dont la structure et dont la structure détaillée est indiquée par la figure 2 C ; les stomates existent aussi sur la face supérieure.
?Un stomate ( fig 2 D) est formé d'un orifice ou ostiole , limité par deux cellules arquées, et qui fait communiquer l'extérieur avec une lacune des tissus sous-jacents appelée chambre sous-stomatique.Les parois des deux cellules bordantes qui sont tournées du côté de l'ostiole sont épaissies. Un mécanisme, qui ne peut être décrit ici, fait que lorsque les cellules qui bordent l'ostiole sont turgescentes , elles se bombent en ouvrant l'orifice ; au contraire lorsqu'elles sontpauvre en eau, c'est à dire lorsque les tissus de la feuille ont subit une fanaison partielle , ces cellules s'affaissent et l'ostiole se trouve ainsi obturée. Ce processus joue , semble - t-il , un rôle dans la régulation de la transpiration. Parfois, les cellules bordantes sont accompagnées de cellules annexes ( fig 2 C ). Souvent, les cellules de l'épiderme ont une forme particulière, comportant des sinuosités qui s'engrènent les unes dans les autres ( cellules dites en jeu de patience ).
?Fig 2
Les nervures qui parcourent le limbe se ramifirent un très grand nombre de fois et les plus fines d'entre elles forment un réseau qui s'anastomose en constituant des mailles. Les plus fines nervures ne sont plus formées que de quelques vaisseaux de bois, non accompagné de liber.
?L'anatomie du pétiole ressemble beaucoup à celle de la tige et comporte généralement un cercle de faisceaux libéro-ligneux dont le bois est tourné vers l'axe de l'organe. Toutefois, la forme fréquemment semi-cylindrique du pétiole et le peu de place laissé aux faisceaux conduisent à des altérations de cette structure théorique . Les faisceau se disposent alors d'une manière irrégulière , ou sur plusieurs rangs , ou suivant plusieurs cercles adjacent. Le pétiole qui doit supporter le poids du limbe souvent de grande taille, peut présenter un développement particulier des tissus de soutien; rarement il peut cependant être creux .
La chute des feuilles se fait par le jeu d'une différenciation particulière constituée par une assise de liège qui se développe en travers du pétiole.
II.3 Méristèmes et structures secondaires
Si les tissus primaires se constituent très précocement au cours du développement des organes , il apparaît par la suite des tissus secondaires . Les seconds se mettant en place au sein des premiers, les cellules des tissus primaires doivent subir une dédifférenciation permettant un retour à l'état méristématique . D'où l'importance de la dédifférenciation cellulaire véritable différenciation à rebours. Les méristèmes secondaires ( ou cambiums) issus de cette dédifférenciation sont à l'origine des tissus secondaires responsables de la croissance en épaisseur de certains rachéophytes . En outre c'est grâce à la dédifférenciation cellulaire que tous les organes adventifs apparaissent
III.3.1 LA DEDIFFERENCIATION CELLULAIRE
?a) Ses caractères
Au cours de ce phénomène , la cellule perd les propriétés qu'elle avait acquises, et inversément retrouve celles qu'elle avait perdues en se différenciant. Si la différenciation entraîne un vieillissement cellulaire , la dédifférenciation se traduit par un rajeunissement , les caractères de la dédifférenciation sont donc :
- Perte des fonctions physiologiques spécifiques et perte concomitante des caractères structuraux correspondants.
?- Diminution du rapport paraplasme/ protoplasme.
- Reprise de l'activité mitotique.
?- Réapparition de potentialités histogénétiques étendues. Les cellules " dévoilent alors des potentialités dont les corrélations organiques inhibaient la manifestation dans l'organisme normal" (Buvat)
b) Les phases de dédifférenciation.
?Etudiant ce phénomène chez différentes espèces d'Angiospermes, Buvat reconnaît, si la différenciation est complète, deux phases précédées, dans certains cas, d'une phase préliminaire :
mc) exemples de dédifférenciation
?Ce sont surtout les Spermatophytes, les Préspermatophytes et les Ptéridophyytes qui sont concernés, bien que des régressions structurales aient été observées aussi chez les Bryophytes.
1) Dédifférenciation et ontogenèse des trachéophytes
- Exemples de dédifférenciation ne dépassant pas la première phase.
- Les arcs cambiaux interfasciculaires de la tige des Spermatophytes se forment par dédifférenciation des cellules parenchymateuses des rayons médullaires de la structure primaire
?- De même, la dédifférenciation des cellules péricycliques est à l'origine d'une partie du cambium libéro-ligneux racinaire
-Enfin, l'installation de la zone génératrice subéro-phellodermique suppose la dédifférenciation préalable de cellules corticales ( parenchyme, collenchyme) ou de cellules du phloème.
- Exemples de dédifférenciation complète :
- Les bourgeons axillaires dans la plupart des cas et tous les bourgeons adventifs s'ébauchent à partir de cellules dédifférenciées
- L'origine endogène des racines secondaires repose sur la dédifférenciation de tissus variés ( endoderme, péricycle , phloème etc.). Pourtant il est fréquent que le péricycle soit plus spécialement impliqué chez les Spermatophytes et l'endoderme chez les Ptéridophytes.
On pourrait dire des choses comparables à propos de l'origine des racines adventives à partir de cellules corticales, péricycliques, libériennes et même cambiales.
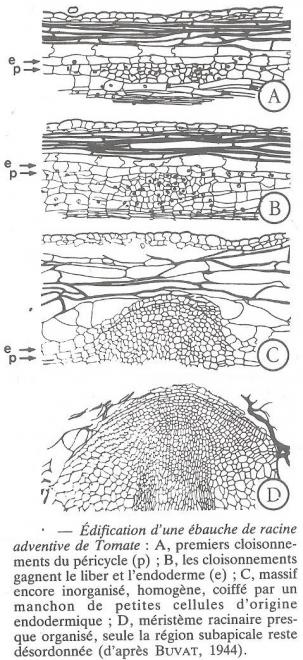
Fig 3
2) Dédifférenciation expérimentale
La dédifférenciation cellulaire peut être déclenchée expérimentalement par la perturbation de corrélations organiques ( traumatismes, action de substances chimiques, excitations parasitaires ) ou par leur supression ( fragmentation d'organes ou de tissus ).
Afin de compléter ce qui a été exposé sur la différenciation du collenchyme , nous empruntons à Buvat les résultats qu'il a rapportés à propos de la dédifférenciation expérimentale de ce tissus chez les tiges de la tomate ( Lycopersicum esculentum/ Solanacées ) :
?Dès qu'une ébauche d'une racine débute en profondeur, le diamètre des cellules de collenchyme s'accroit, tandis que leur paroi diminue d'épaisseur, d'où l'apparition à la surface de la tige d'un mamelon qui matérialise la future région de sortie de la racine adventive. puis les mitoses reprennent et les longues cellules se cloisonnent transversalement , tandis que les chloroplastes se divisent plusieurs fois. les cellules filles sont sensiblement isodiamétriques à parois minces , avec un gros noyau, de nombreux chloroplastes , un cytoplasme peu abondant et une grande vacuole.
?La dédifférenciation ne se poursuit pas plus avant ici , mais elle a pour conséquence d'avoir fait perdre aux cellules collenchymateuses leurs caractères de cellules de soutien , ce qui est capital pour le passage de la jeune racine adventive au travers de l'écorce de la tige . En effet elle ne rencontrera qu'un tissus de faible résistance qu'elle perforera facilement pour sortir . Les mitoses des cellules en dédifférenciation ont permis la détermination de leur nombre chromosomique . A ce sujet, Buvat signalait , dès 1944 l'importance du fait qu'elles sont octoploïdes, ce qui laissait présumer que les tissus d'un végétal pourraient être à des niveaux de ploïdie différents.
d) La plasticité des cellules végétales
Toutes les cellules végétales non dégénérescentes sont susceptibles , ayant perdu leurs caractères de spécialisation, de retourner à l'état méristématique et , après une période de prolifération, de former un tissus différent de celui qu'elles constituaient primitivement. C'est ainsi que Buvat a montré que chez les Angiospermes , peuvent être impliqués dans un processus de dédifférenciation-redifférenciation : des épidermes , des parenchymes chlorophylliens ou non, le collenchyme, l'endoderme, le péricycle, de jeunes tubes criblés et leurs cellules compagnes, le parenchyme libérien et le parenchyme ligneux, mais aussi les cambiums.
Ceci ne signifie pas que n'importe laquelle de ces catégories cellulaires peut être à l'origine d'un individu nouveau. " Ainsi la dédifférenciation des cellules épidermiques des feuilles de Brimeura ou des cellules parenchymateuses de la racine de chicorée conduit à l'édification d'organismes complet tandis que les cellules péricycliques de tomate , celles du parenchyme libérien de la carotte ou du mésophylle de Brimeura ne fournissent que des racines et que les cellules endodermiques ou les cellules compagnes du liber de tomate ne fournissent que certains tissus des ébauches radiculaires " (Buvat).
III.3.2 LES MERISTEMES SECONDAIRES
Ils sont au nombre de deux :
- La zone génératrice libéro-ligneuse ou cambium libéro-ligneux, responsable de la formation de liber et de bois.
?- La zone génératrice subéro-phellodermique, ou phellogène, responsable de la formation de liège et de phelloderme.
a) Leur formation
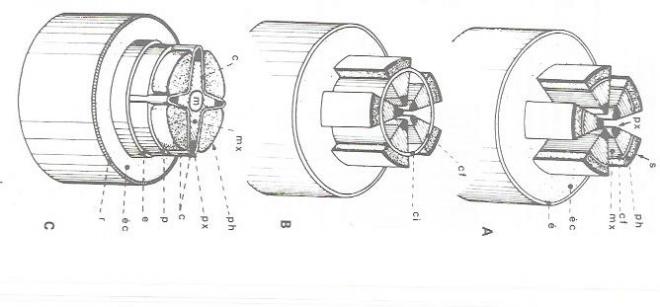
1) La zone génératrice libéro-ligneuse ( fig 4 )
?Dans la tige, au niveau de chaque faisceau criblo-vasculaire, entre le phloème et le xylème, le cambium fasciculaire est un reste du faisceau procambial qui n'a pas subi de différenciation au cours de la réalisation du faisceau primaire. Plus tard, à la suite d'une dédifférenciation des cellules parenchymateuses reliant la moëlle à l'écorce ( rayons médullaires de nature primaire ), apparaissent des arcs de cambium interfasciculaires qui, en réunissant entre eux les arcs de cambium fasciculaire, constitueront un manchon méristématique continu.
?Les faisceaux primaires des nervures de la feuille des Gymnospermes et des dicotylédones peuvent acquérir aussi du cambium fasciculaire.
Au niveau de la racine, alors que l'évolution vasculaire primaire n'est pas encore terminée, des cellules situées sur la face interne des faisceaux de phloème entrent en division, et réalisent des arcs cambiaux discontinus. Plus tard, au niveau des pôles ligneux, des cellules péricycliques dédifférenciées construisent de nouveaux arcs cambiaux se raccordant aux précédents.
Il résulte de la différence qui existe entre la structure primaire de la tige et celle de la racine que, sur des coupes transversales d'organes, le manchon méristématique secondaire apparaît d'abord circulaire dans le premier cas, sinueux et étoilé dans le second cas. Rapidement, cette différence s'estompe, dès que les formations secondaires libéro-ligneuses débutent, le manchon devenant alors circulaire dans la racine comme dans la tige.
 ?fig 4
?fig 4
2) La zone génératrice subéro-phellodermique
En principe elle n'existe pas dans la tige et la racine des dicotylédones herbacées, ni dans les feuilles d'une manière générale. Son apparition plus tardive que celle du cambium libéro-ligneux, se fait au niveau de zones privilégiées, sur l'emplacement de futures lenticelles. Dans la tige, elle occupe des positions très variées suivant les espèces , de l'assise corticale sous épidermique ( le plus fréquemment ) jusque dans le liber et, plus rarement, dans le bois et même dans la moelle . Dans tous les cas, une dédifférenciation préalable des tissus corticaux ( parenchyme, collenchyme ), des tissus conducteurs et des cellules médullaires est à son origine. il arrive que la même zone génératrice fonctionne pendant toute la vie du végétal ou presque , mais, le plus souvent, on assiste à des remplacements successifs, une zone qui en relaie une autre étant toujours en position plus profonde que la précedente.
?dans la racine, elle peut se situer directement directement sous l'assise pilifère, dans l'écorce et pour la plupart des espèces , au niveau du péricycle. Dans ce cas, son origine est liée à celle du cambium libéro-ligneux . Après dédifférenciation et division des cellules péricycliques placées en face des pôles ligneux , parmi les cellules filles, les plus internes formeront les arcs de la zone libéro-ligneuse , les plus externes, l'amorce de la zone subéro-phellodermique . Partie de la zone des pôles ligneux , la dédifférenciation s'étend alors à tout le péricycle, d'ou réalisation d'une zone cambiale continue.
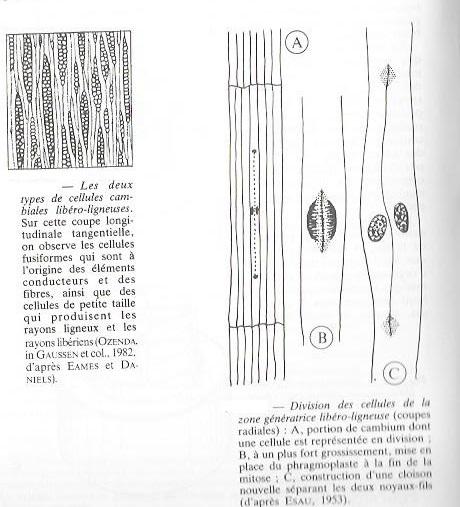
Fig 5
b Leurs caractères
Le tableau 6 résume les caractères des méristèmes secondaires. Il correspond plus particulièrement aux cellules qui engendrent le xylème et le phloème secondaires. Mais , celles qui sont à l'origine des rayons ligneux et des rayons libériens( ou parenchyme horizontal ) sont beaucoup plus petites presque isodiamétriques, groupées en paquets intercalés parmi les précédentes ( fig 5 ).
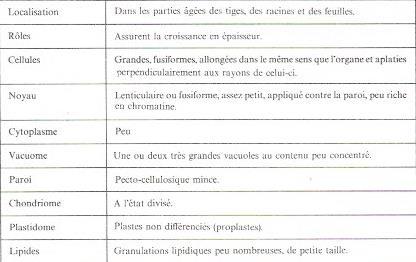 ?Fig 6
?Fig 6
c Leur activité
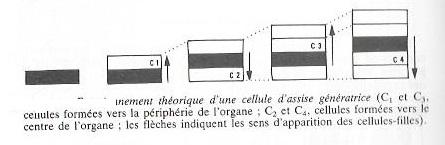
Par des cloisonnements longitudinaux périclines ( fig 5 ) , ces cellules se divisent activement sur leurs faces internes et externe, mettant en place des files radiales de cellules filles (fig 7). L'épaisseur de la zone génératrice ne grandit pas pour autant, les cellules filles se différenciant en tissus secondaires. Il en résulte une croissance en épaisseur de l'organe se manifestant de part et d'autre de la zone cambiale . Des cloisonnements anticlines de cellules méristématiques permettront à cette zone d'accompagner la croissance en épaisseur de la tige et de la racine. En principe les cellules cambiales sont susceptibles de se diviser alternativement sur leurs faces externe et interne, mais cette périodicité n'est pas aussi régulière que semble l'indiquer la figure 7. En effet, il se forme toujours plus de bois que de liber, de liège que de phelloderme.
Enfin, l'activité des cambiums est rythmique. Sous notre climat tempéré, elle débute au printemps chez les arbres, dans les rameaux dont les bourgeons débourrent et progressivement s'étend aux branches, au tronc, puis aux racines. Au début de l'automne cette activité cesse dans le même ordre qu'elle avait débuté au printemps. En fait les modalités de la réactivation cambialevarient avec l'age de l'arbre. Par exemple , la reprise d'activité du cambium libéro-ligneux est lente et basipède chez les arbres jeunes et les rameaux des grands arbres, où elle dépend de l'activité des bourgeons apicaux . Si les arbres sont agés, la réactivation du cambium se fait simultanément en plusieurs zones et se généralise très vite . Chez le Hêtre âgé, par exemple, le cambium du tronc sort de dormance peu après la reprise des mitoses à l'extrémité des rameaux, mais avant que celui de la base des branches soit réactivé: le tronc âgé manifeste donc une certaine autonomie vis à vis de la cime de l'arbre et des bourgeons terminaux .
Date de dernière mise à jour : 25/10/2017
Ajouter un commentaire