

Anatomie 3
Structures primaires
III.2 Structures primaires
Les cellules issues de la division des cellules méristématiques primaires se différencient en tissus primaires, dont la disposition relative dans un organe constitue sa structure primaire.
III.2.1 LA STELE DES TRACHEOPHYTES
La présence constante de tissus conducteurs leur donne une grande importance dans la caractérisation de la structure primaire. Depuis Van Tieghem et Douliot ( 1886 ), on appelle stèle ou cylindre central, l'ensemble des tissus conducteurs primaires occupant la région axiale d'un organe. A la périphérie, la stèle est entourée par le péricycle ( une ou plusieurs assises cellulaires de nature parenchymateuse ).
L'écorce qui enveloppe la stèle est comprise entre un endoderme, assise cellulaire profonde au contact du péricycle et un épiderme ou un rhizoderme externe.
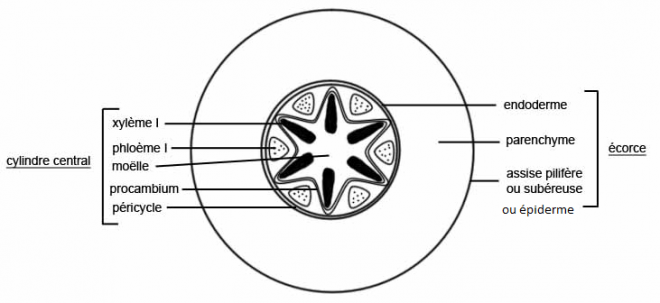 ?Fig 1 : coupe transversale dans une racine de dicotylédone.
?Fig 1 : coupe transversale dans une racine de dicotylédone.
Les stèles étant différentes d'un organe à l'autre pour un individu donné ou d'un groupe systématique à l'autre pour un organe donné, leur étude se révèle importante, sur le plan anatomique, comme sur le plan phylogénique. Les différents types de stèle seront présentés, du plus simple et du plus archaïque ( protostèle ), au plus évolué ( eustèle ).
a) Les stèles sans moëlle axiale
- La protostèle
Une protostèle ( du grec protos = premier et stêlé = colonne ) est constituée d'une colonne de sylème cylindrique, entourée d'un manchon de phloème.
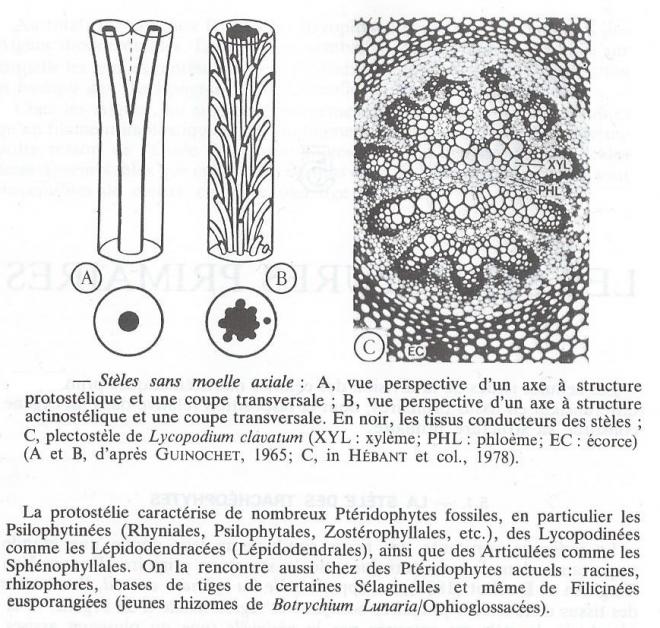 Fig 2
Fig 2
?- L'actinostèle
C'est une variante de la protostèle dont la colonne de xylème est plus ou moins cannelée. En coupe transversale, la stèle est donc étoilée, d'ou son nom ( du grec aktis, inos = rayon ) et la qualification de monarche, diarche, triarche...polyarche suivant le nombre de bras de l'étoile.
Comme les protostèles, les actinostèles sont qualifiées de monarches, diarches... polyarches et d'endarches, mésarche, exarches suivant le nombre et la position ( centrale, intermédiaire ou périphérique ) des pôles ligneux, centres d'origine de la différenciation du xylème. C'est à leur niveau qu'apparaissent donc les premiers éléments conducteurs, les plus primitifs d'ailleurs ; en particulier des trachéides annelées de faible diamètre constituant le protoxylème. Par la suite, des trachéides moins primitives et de diamètre plus important sont mises en place ( métaxylème ) .

Tableau 1
La protostélie caractérise de nombreux Ptéridophytes fossiles, en particulier les Psilophytinées ( Rhyniales, Psilophytales, Zostérophyllales, etc...) , des Lycopodinées comme les Lépidodendracées ( Lépidodendrales ), ainsi que les Articulées comme les Sphénophyllales. On la rencontre aussi chez des Ptéridophytes actuels: racines, rhizophores, bases des tiges de certains Sélaginelleset même de Filicinées eusporangiées ( jeune rhizomes de Botrychium Lunaria/Ophioglossacées).
- La plecostèle
Actinostèle polyarche dont le xylème envahi par des rubans de phloème est disloqué, la plecostèle est bien représentée chez de nombreux Lycopodes.
?b) Les stèles avec moëlle axiale
- La siphonostèle
?Cette fois, xylème et phloème constituent deux manchons cylindriques emboîtés entourant une moelle. La colonne de xylème n'est donc pas pleine ( du grec siphôn = tube )
Si, en coupe transversale, la couronne de xylème est entourée par une couronne de phloème, la siphonostèle est ectophloique, alors qu'elle est amphiphloique quand la couronne de xylème est située entre deux couronnes de phloème. D'une manière générale, les siphonostèles sont très fréquentes chez les Ptéridophytes.
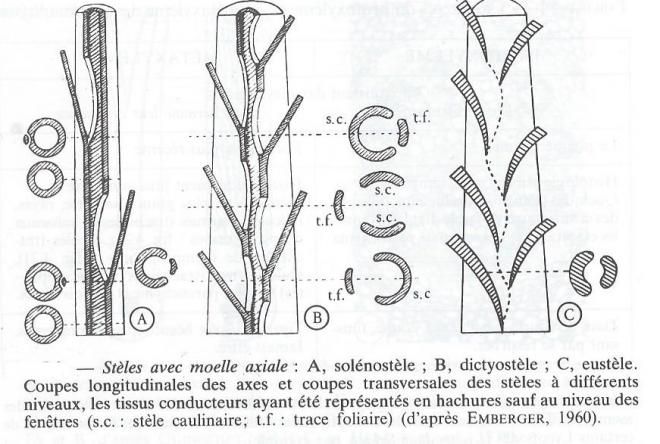
Fig 3
- Solénostèle et dictyostèle
Solénostèle et dictyostèle sont ajoutées du fait de l'existence de fenêtres ( ou brêches ), solutions de continuité intéressant à la fois le xylème et le phloème et constituées de parenchyme. Si les fenêtres sont rares, on n'en rencontre pas forcément dans une coup transversale ou, tout au plus une seule. On est alors en présence d'une solénostèle ( du grec sôlên = tuyau ). Quand le nombre des fenêtres grandit, plusieurs sont alors visibles sur une même coupe. La solénostèle est devenue une dictyostèle ( du grec dicktuon = réseau ) fragmentée en plusieurs méristèles ( du grec meris = partie ). Si la dictyostélie est fréquente chez les Ptéridophytes, surtout parmi les Filicinées, elle est très rare chez les Spermatophytes.
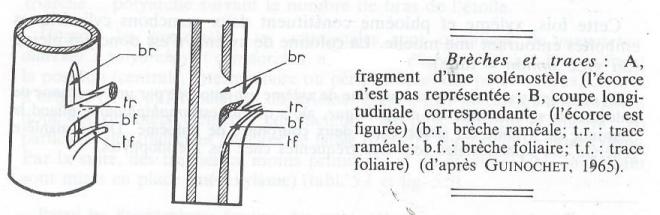
fig 4
- L'eustèle
Si le processus de fragmentation de la stèle caulinaire se développe par l'agrandissement des fenêtres foliaires, la vascularisation caulinaire disparaît et les cordons conducteurs ne sont plus représentés que par des traces foliaires. On est alors en présence d'une eustèle endarche extrêmement répandue ( Ptéridophytes évolués et Spermatophytes ). Dans ce cas, tout se passe comme si l'axe était composé d'autant d'unités que de feuilles, par concrescence de bases foliaires ou de feuilles plus ou moins modifiées.
II.I 2.2 LA STRUCTURE PRIMAIRE DES SPERMATOPHYTES
Nous dégagerons ici les caractères principaux de la structure primaire des tiges, racines et feuilles.
a) La tige
Comme notre propos est de comparer la structure primaire de la tige, de la feuille et de la racine, et que ce dernier organe ne comporte aucun noeud, nous n'envisagerons ici que la structure internodale.
1) Angiospermes dicotylédones et Gymnospermes
?- L'épiderme est cutinisé, stomatifère, avec ou sans poils.
- L'écorce ou cortex est parenchymateuse avec ( ou sans ) tissus de soutien ( collenchyme, sclérenchyme ) disposés en anneaux continus ou en îlots séparés.
- Les faisceaux criblo-vasculaires disposés sur un cycle unique comprennent du xylème et du phloème superposés, le xylème situé au contact de la moelle étant coiffé vers l'extérieur par le phloème. Le xylème est constitué de protoxylème ( du grec prôtos = premier et xulon = bois), et de métaxylème ( du grec meta, ce préfixe indique une succession ) . De même on distingue un protophloème et un métaphloème de structure plus complexe.
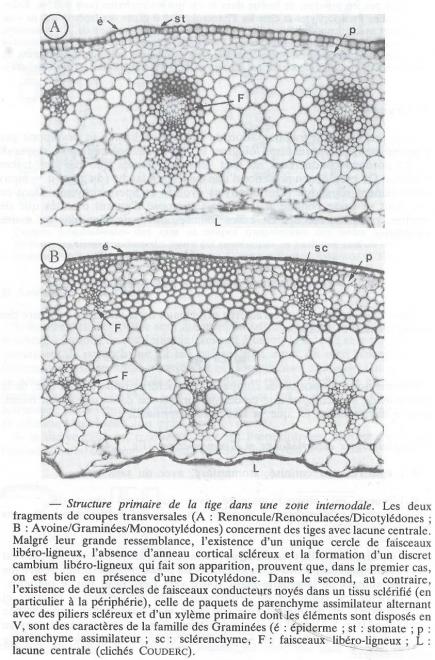
Fig 5
Un exemple typique de tige est donné par la figure 6. La tige de fenouil comporte de dedans en dehors : une moelle, un cercle de faisceaux libéro-ligneux, une écorce et un épiderme. L'écorce présente des bandes longitudinales de tissu de soutien à cellules épaissies, et des canaux sécréteurs produisant une essence qui donne à la plante son odeur caractéristique. L'examen à fort grossissement d'un fragment de cette coupe transversale ( fig 6) montre le détail des principaux tissus. La moelle et l'écorce sont constituées de cellules banales, sans différenciation particulière : c'est du parenchyme. Les côtés longitudinales de tissus de soutien apparaissent , en coupe , sous forme d'îlots elliptiques situés au dessous de l'épiderme ; les parois des cellules sont considérablement épaissiespar un dépôt de cellulose qui leur donne de la rigidité, ce tissus est du collenchyme.
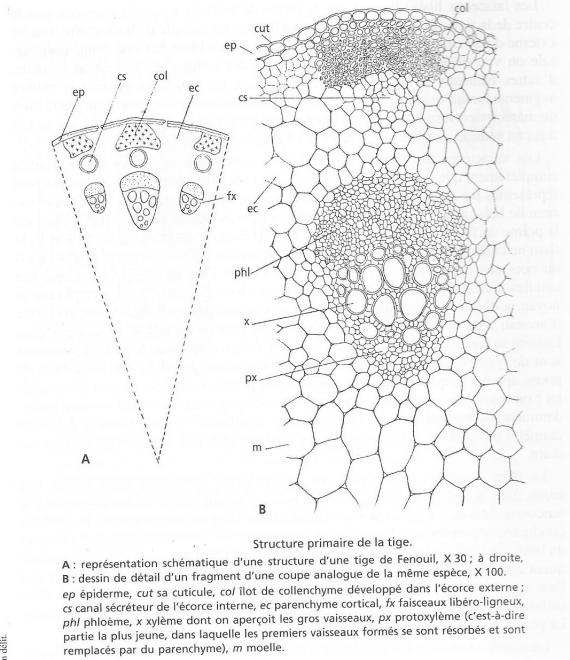
Fig 6
Les canaux sécréteurs sont formés d'un cylindre de cellules sécrétrices entourant la lumière du canal dans lequel se déverse l'essence sécrétée. De tels canaux existent d'ailleurs dans toute la plante et dans la plupart des espèces de la famille des Apiacées, à laquelle appartient le fenouil.
Enfin, l'épiderme est constitué d'une seule assise de cellules qui présentent sur leur paroi extérieure un épaississement imprégné d'une substance particulière, la cutine; cette paroi épaissie est appelée cuticule.
?Les faisceaux libero-ligneux ont la forme de secteurs à pointes tournées vers le centre de la tige ; ils comprennent du côté interne un triangle de bois , coiffé du côté externe d'une calotte sensiblement semi-circulaire de liber. Sur une coupe transversale on voit déjà que le bois contient, parmi des cellules de dimension normale, d'autres éléments beaucoup plus gros qui sont les vaisseaux et dont le calibre augmente lorsqu'on va de la pointe du bois en direction du liber : cette augmentation de diamètre correspond à une différenciation progressive des vaisseaux, qui se fait dans un sens dit centrifuge par rapport à l'axe de la tige.
?Les caractères histologiques des tissus conducteurs ne peuvent être étudiés complètement que sur une coupe longitudinale ; des fragments de telles coupes sont représentés par la figure 7. La section longitudinale passant par l'axe d'un faisceaux de bois montre la succession des éléments vasculaires qui le composent ; vers la pointe du faisceau, dans la partie qui est représentée sur la figure 7 A et B, le tissus ne contient que des éléments conducteurs et des cellules de parenchyme ; c'est du bois peu différencié, le premier formé et que l'on appelle protoxylème . Les cellules conductrices sont allongées en tubes ; leur contenu vivant, cytoplasme et noyau, a disparu et les parois sont garnies d'épaississements de lignine, en forme d'anneaux dans les premiers vaisseaux formés, puis de spirale et enfin de réseau . Ensuite se forme un bois plus différencié, dit métaxylème , dans lequel , dans lequel les vaisseaux sont de type ponctué : leur membrane est entièrement lignifiée sauf au niveau de pores appelés ponctuations et dont la structure, parfois complexe ne sera pas étudiée ici; on trouve en outre dans le métaxylème du parenchyme vivant comme précédemment mais aussi des cellules mortes et lignifiées, très allongées et de faible diamètre qui sont les fibres et qui seront étudiées plus loin à propos du bois secondaire.
?Le liber comporte pareillement un protophloème différencié dans la tige très jeune, dans la partie externe du faisceau, puis un métaphloème qui se différencie à la rencontre du bois. La partie la plus importante du liber est constituée par les cellules conductrices proprement dites, appelées tubes criblés: contrairement aux vaisseaux du bois , elles ont conservé leur cytoplasme , le noyau seul a disparu. Elles communiquent entre elles par des ponctuations spéciales formées de très petits pores et que l'on appelle cribles. Elles sont en connexion avec des cellules plus petites dites cellules compagnes, à cytoplasme et noyau persistants et dont le rôle est incertain. Le phloème contient également du parenchyme et parfois des fibres.
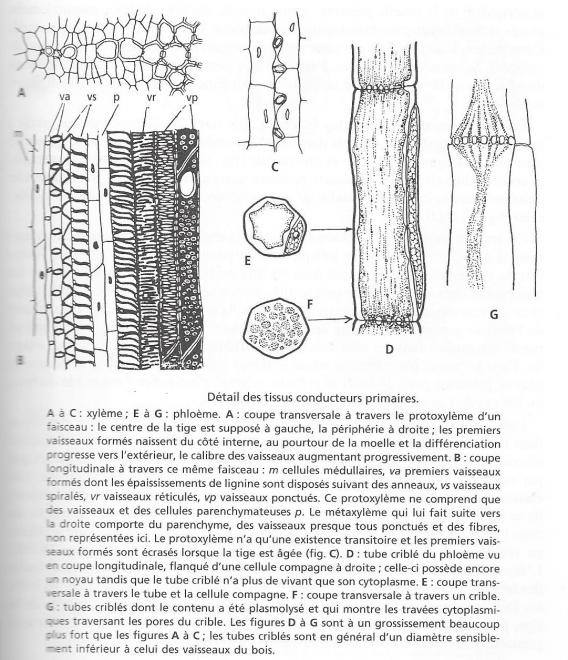
Fig 7
La nature chimique des constituants des divers tissus : cellulose et autres substances de membrane, lignine, cutine, produits de sécrétion, etc., ne sera pas faite ici ; se reporter à ce sujet au cours de biochimie.
La structure primaire des tiges peut varier considérablement d'une plante à l'autre et les exemples représentés sur la figure 8 donne une idéede ces variations . Chez le Genêt d'Espagne ( Spartium, Papilionacées ), la tige ( fig 8 A)comporte encore un cercle de faisceaux libero-ligneux de structure analogue aux précédents, mais chaque massif de phloème est coiffé d'une calotte de tissus de soutien qui, ici, a des parois cellulaires imprégnées de lignine et porte le nom de sclérenchyme . D'autres faisceaux de sclérenchyme courent dans l'écorce. Celle-ci est formée en grande partie d'un parenchyme particulier à cellules chlorophylliennes, et la tige a un rôle assimilateur qui compense dans cette espèce la réduction des feuilles. Enfin, le centre de la tige , occupe à l'état jeune par une moelle lâche, est ensuite creux , en raison de la formation d'une grande lacune par résorption de la moelle.
La tige de la plupart des arbustes, mais aussi celle d'un certain nombre de plantes herbacées, présente un cercle complet de faisceaux libero-ligneux; il en est ainsi dans la tige du liseron, figurée sur la figure 8 B . Mais, en outre, dans cette tige la périphérie de la moelle présente, contre le bois, des ilôts de liber anormalement placés et dont la structure histologique est identique à celle du liber externe normal. Cette présence de liber interne caractérise un certain nombre de familles : Convolvulacées, Solanacées, Myrtacées. Parfois la tige peut contenir des cordons libériens dispersés dans la moelle ( liber médullaire ) ou même des faisceaux libero-ligneux médullaires.
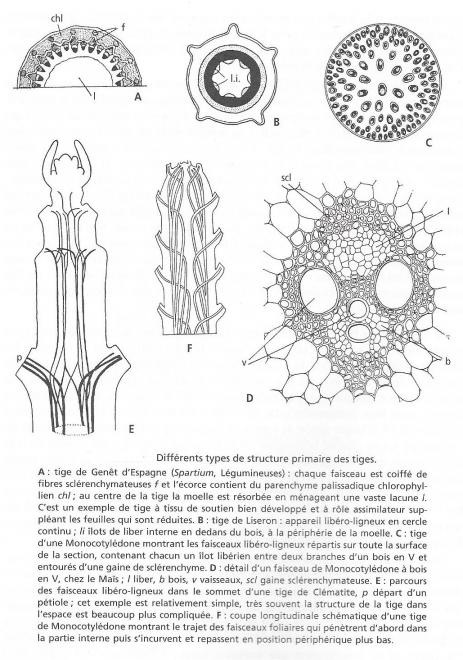 Fig 8
Fig 8
Chez les Monocotylédones, les faisceaux sont rarement disposés sur un cercle; presque toujours ils sont répartis dans tout l'ensemble de la tige de sorte qu'on ne peut plus distinguer une écorce et une moelle ( fig 8 C ). En coupe transversale, le contour du xylème des faisceaux présente souvent une forme de V enserrant le phloème dans sa concavité, tandis qu'une gaine de sclérenchyme ceinture le faisceau ( fig 8 D).
?Nous avons appelé stèle l'ensemble du système conducteur de la tige , et notamment le cylindre libéro-ligneux lorsque cette disposition simple est réalisée. Toutefois , la variabilité de cet appareil dans les différents groupes de Phanérogames ( et plus encore chez les Ptéridophytes ), la difficulté de séparer ce qui appartient à la tige et ce qui correspond déjà au départ des feuilles , la connaissance encore insuffisante de l'organogenèse de beaucoup de tiges, font que le terme stèle a été employé par divers auteurs dans des sens souvent très souvent différents . Dans la racine, par contre, le même terme de stèle a été employé avec beaucoup plus de précision pour désigner le cylindre central, c'est à dire l'ensemble du bois, du liber et de l'assise péricyclique qui les entoure.
Pour être complète, une étude anatomique doit figurer la disposition des tissus dans l'espace. Il ne faudrait pas croire que celle-ci se déduit de ce que l'on observe en coupe transversale, par une simple translation parallèle à l'axe de l'organe, et que par exemple les faisceaux libero-ligneux constituent une sorte de colonnade. Les choses sont beaucoup plus compliquées. Ces faisceaux suivent une courbe sinueuse, ils se ramifient, se fusionnent, peuvent parcourir un certain trajet dans la moelle ou dans l'écorce avant de se rendre dans les feuilles. Le " cylindre vasculaire " affecte en réalité dans l'espace une disposition grillagée, visible sur la figure 8 E. L'émergence des faisceaux qui entrent dans les pétioles découpe dans ce cylindre des brèches que nous avons appelées " fenêtres foliaires". Chez les Monocotylédones ( fig 8 F), les faisceaux qui proviennent des feuilles pénètrent d'abord dans la moelle de la tige, puis au cours de leur descente dans celle-ci ils s'incurvent vers l'extérieur et prennent place dans les cercles libéro-ligneux externes.
Date de dernière mise à jour : 25/10/2017
Ajouter un commentaire


