
5.La cellule
BIOLOGIE VI : EXPLORATION DE LA CELLULE
VI.1 L’importance des cellules
Tous les organismes sont composés de cellules, c’est le premier niveau hiérarchique du vivant capable de vivre.
Bien des organismes ne sont composés que d’une cellule
Animaux et végétaux sont composés de plusieurs cellules spécialisées qui sont incapables de vivre isolées de l’organisme.
Même associées en organismes pluricellulaires pour atteindre un niveau de complexité supérieur (tissus, organes), elles restent à la base de la structure et du fonctionnement de ceux-ci.
Tout ce qu’un être vivant réalise, il le doit d’abord à son activité cellulaire
La vie à l’échelle cellulaire naît d’un ordre structurel, ce qui confirme l’axiome structure/fonction et les propriétés émergentes d’un système complexe.
Les cellules sont capables de détecter les modifications dans leur environnement et d’y réagir.
A partir de la cellule ancestrale, les cellules ont été soumises au processus évolutif et se sont diversifiées.
VI.2 Utilisation de microscopes et d’instruments de biochimie pour étudier les cellules
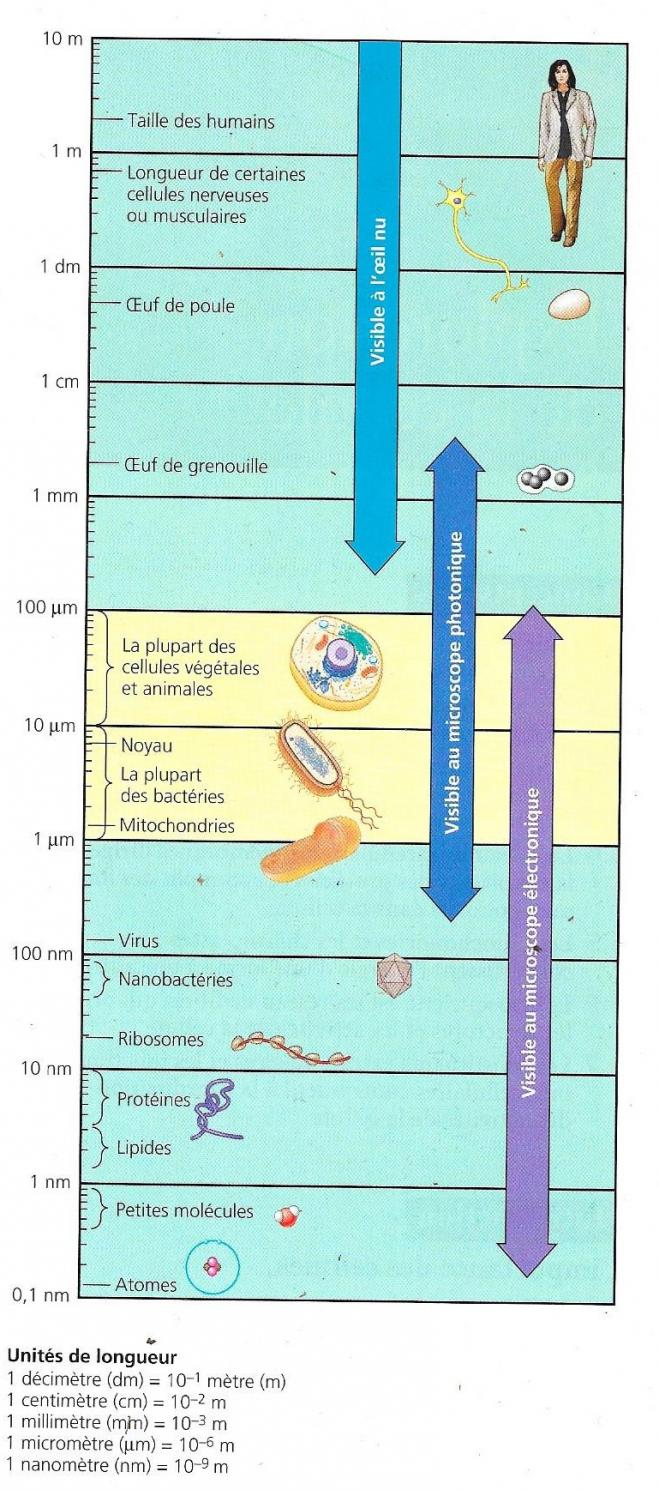
Les cellules sont de taille microscopique, il est difficile de concevoir leur degré de complexité. Des techniques existent cependant pour les observer.
2.1 La microscopie
A) INTRODUCTION
L’évolution de la science est tributaire de l’évolution de moyens techniques permettant à l’homme d’aller au-delà de ses sens. C’est le cas de la microscopie découverte au 16ème siècle et de ses évolutions. A l’heure actuelle, on ne peut explorer la cellule qu’à l’aide de diverses techniques microscopiques.
B MICROSCOPES PHOTONIQUES
Les microscopes utilisés à la renaissance, tout comme ceux que l’on retrouve classiquement dans les laboratoires sont les microscopes photoniques.
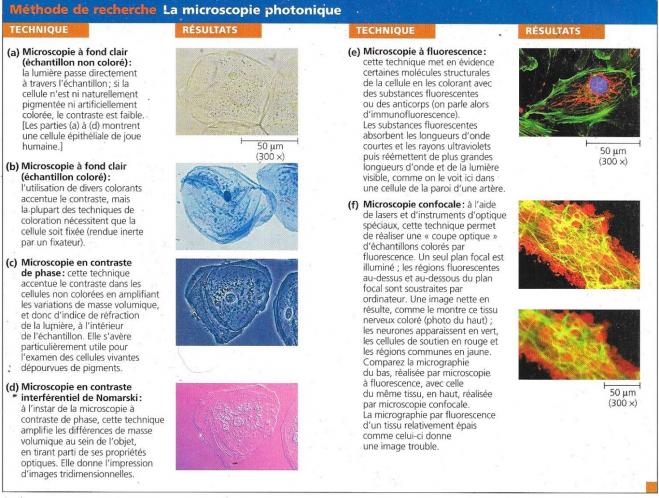
Lors de l’utilisation d’un microscope photonique, la lumière traverse la préparation et ensuite des lentilles de verre qui la réfractent en manière telle qu’une image agrandie se forme dans l’oeuil ou une plaque photographique. Deux facteurs sont importants en microscopie : en premier lieu le grossissement qui correspond au rapport de la taille de l’image à la taille de l’objet réel. L’autre facteur est le pouvoir de résolution qui donne une idée de la clarté de l’image, plus précisément il correspond à la mesure de la distance à partir de laquelle deux objets ne sont plus discernables (l’oeuil peut voir une étoile simple là où le télescope verra une étoile double).
Ce pouvoir de résolution est limité alors que le grossissement est théoriquement sans limite. Les microscopes photoniques grossissent efficacement jusque 1000 fois, mais au-delà les images deviennent brouillées. En effet le pouvoir de résolution est limité à 200 nanomètres qui correspondent à la taille d’une petite bactérie ou d’une mitochondrie. Ce pouvoir de résolution ne peut être modifié car il est lié aux longueurs d’ondes du spectre visible. C’est donc sur le contraste que l’on a amélioré les microscopes, soit mieux faire ressortir les détails déjà distinguables. A cela ont été adjointes des techniques de coloration permettant de mettre en évidence des compartiments cellulaires particuliers.
C MICROSCOPES ELECTRONIQUES
La plupart des organites ou structures cellulaires sont difficilement visibles en microscopie photonique. L’invention dans les années 1950 du microscope électronique a permis à la biologie cellulaire de faire des pas de géants. Au lieu d’utiliser la lumière, le microscope électronique utilise un faisceau d’électrons auquel correspond une longueur d’onde. Ce faisceau traversera la préparation ou la balayera et le pouvoir de résolution sera inversement proportionnel à la longueur d’onde utilisée. La longueur d’onde des faisceaux d’électrons est beaucoup plus grande que celle de la lumière et c’est ainsi que le pouvoir de résolution sera grandement augmenté.
De manière courante, le grossissement permis par le microscope électronique peut aller jusque 100.000 fois sans que l’image soit trop brouillée. On appelle ultrastructure cellulaire, l’anatomie cellulaire que le microscope électronique peut révéler.
En fonction de ce qui est écrit plus haut, on distingue des microscopes électroniques à balayage et des microscopes électroniques à transmission
Le microscope électronique à balayage permet de faire un examen détaillé de la surface d’un échantillon. Dans ce cas le faisceau électronique balaye cette surface qui est préalablement recouverte d’une mince pellicule d’or ou de platine. Cette pellicule va alors émettre des électrons « secondaires ». Ceux-ci seront détectés par un instrument qui traduit la disposition de ceux-ci en un signal électronique visible sur un écran.
Le microscope électronique à transmission permet d’étudier l’ultrastructure cellulaire interne. Le faisceau électronique traverse une couche très mince de l’échantillon (moins de 0,1 µm), un peu à la manière dont un rayon lumineux traverse l’échantillon en microscopie photonique. Pour accentuer le contraste, on colore des coupes très minces préalablement fixées. Les structures fixées sont des structures ayant subit des traitements physiques ou chimiques entraînant la mort de celles-ci. La coloration se fait à l’aide de certains atomes de métaux lourds qui s’attachent à certaines structures cellulaires. La densité électronique de certaines structures par rapport à d’autres s’en trouve ainsi accrue. Au niveau des structures plus denses, les électrons sont plus dispersés et moins d’électrons seront transmis. L’image est le résultat des différences de densité électronique dans l’échantillon.
D COMPARAISON DES DEUX GRANDES TECHNIQUES MICROSCOPIQUES
L’usage du microscope électronique permet de révéler des structures indécelables en microscopie photonique, mais celui-ci a l’avantage de pouvoir aider à mener des observations sur des échantillons vivants. Il faut également mentionner que la fixation des échantillons peut produire des «artefacts» à l’observation, qui sont des structures inexistantes sur cellules vivantes.
La cytologie utilise les diverses techniques microscopiques pour décrire la cellule sous tous ses aspects, mais cette description donne peut de renseignements sur la fonction des structures. C’est pour pallier ce problème que la biologie cellulaire moderne allie les points de vue cytologique et biochimique.
Evoquons maintenant la technique de fractionnement cellulaire qui a permis de faire évoluer grandement nos connaissances du sujet.
2.2 Le fractionnement cellulaire.
Il s’agit ici de décomposer la cellule de manière à pouvoir en isoler les principaux organites. L’instrument utilisé est une centrifugeuse capable de faire tourner à différentes vitesses des éprouvettes contenant des cellules dissociées. Il y aura fractionnement car la force centrifuge isole les constituants selon leur taille et leur masse volumique. Les centrifugeuses les plus puissantes appelées ultracentrifugeuses peuvent effectuer 130.000 révolutions par minute et appliquer aux particules des forces jusqu’un million de fois plus grandes que la gravitation.
Cette technique permet d’isoler des constituants cellulaires en grande quantité, permettant ainsi d’étudier leur composition et leur métabolisme.
Il a ainsi été possible aux cytologistes d’associer aux organites, leurs fonctions cellulaires.
VI.3 Les membranes internes compartimentent les fonctions des cellules eucaryotes
L’unité fonctionnelle de base de tout organisme est la cellule, soit procaryote, soit eucaryote.
Seuls les organismes appartenant au domaine des bactéries et des archéobactéries sont des procaryotes. En revanche, les protistes, les mycètes, les végétaux et les animaux sont formés de cellules eucaryotes
3.1 Comparaison de cellules eucaryotes et procaryotes
Cellules procaryotes et eucaryotes possèdent des caractéristiques communes fondamentales :
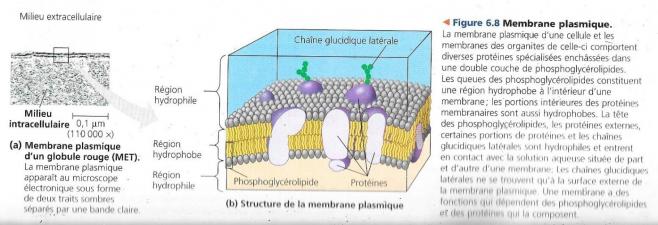
- Elles sont pourvues d’une membrane appelée membrane plasmique qui circonscrit leurs organites.
- Les organites baignent dans une substance semi-liquide appelée cytosol.
- L’ensemble formé par le cytosol et les organites porte le nom de cytoplasme
- Elles possèdent un matériel génétique sous forme d’ADN et des ribosomes qui sont de « petites usines » chargée de synthétiser les protéines sous le commandement des gènes.
Mais elles comportent des différences. :
- L’ADN des procaryotes est concentré dans une région que l’on appelle le nucléoïde, aucune membrane ne le sépare du reste du cytoplasme.
- L’ADN des eucaryotes, leurs chromosomes, se trouvent dans un organite entouré d’une membrane appelé noyau. Et, suspendus dans le cytosol entre la membrane nucléaire et la membrane plasmique, baignent divers organites membraneux aux formes spécifiques et aux fonctions spécialisées. La plupart de ces organites n’existent pas chez la cellule procaryote.
- Une différente se marque également au niveau de la taille des cellules eucaryotes et procaryotes. Comme d’autres caractéristiques générales de la structure cellulaire, la taille est liée à la fonction. Pour accomplir ses fonctions métaboliques, la cellule ne doit être ni trop grande ni trop petite. Les plus petites cellules sont les nanobactéries (découvertes en 1992) de 50nm de diamètre. Il s’agit peut-être là du plus petit format pouvant contenir suffisamment d’ADN pour programmer le métabolisme et assez d’enzymes et d’équipements cellulaires nécessaires pour maintenir la vie et la reproduction. La plupart des bactéries mesurent de 1 à 10 µm de diamètre. Elles sont donc de 20 à 200 fois plus grandes environ que les nanobactéries. Les cellules eucaryotes elles ont typiquement 10 à 100 µ.
La membrane plasmique, périphérie de chaque cellule, tient lieu de barrière spécifique assurant le passage nécessaire et adéquat de dioxygène, de nutriments et de déchets pour desservir le volume entier de la cellule. Plus la surface cellulaire (µm2) est importante par rapport au volume (µm3), mieux c’est possible. Surface et volume sont fonction du rayon comme la taille de la cellule. En fonction de la taille, du rayon, le volume croît plus vite que la surface et le rapport surface/volume diminue. A partir d’une certaine taille, la surface est trop faible pour permettre que le dioxygène, les nutriments et les déchets soient suffisamment échangés pour maintenir la vie et la reproduction. Les cellules sont donc, pour la plupart, microscopiques.
- Généralement les organismes plus grands n’ont pas de cellules plus grandes que les organismes de plus petite taille, ils en contiennent simplement plus. Le rapport surface volume est d’autant plus important que les cellules ont une fonction d’échange importante de matière avec le milieu, comme les cellules intestinales. La surface de ce genre de cellule comprend parfois de longs et fins prolongements appelés microvillosités qui augmentent la surface d’échange sans modifier de manière significative le volume cellulaire.
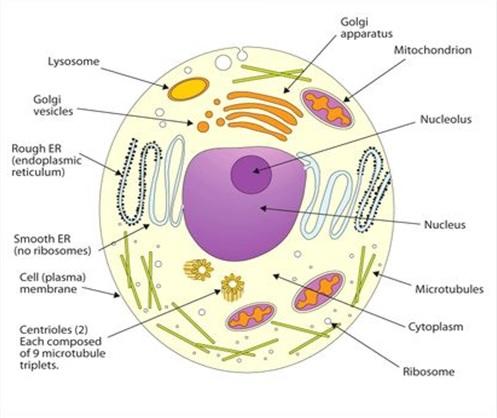
3.2 Vue d’ensemble de la cellule eucaryote.
Outre la membrane plasmique, la cellule eucaryote comporte un réseau élaboré et étendu de membranes internes. Ce sont les organites membraneux évoqués plus tôt et qui compartimentent la cellule Ces membranes participent également au métabolisme cellulaire car beaucoup d’enzymes y sont enchâssées. Chaque compartiment formant une sorte de microenvironnement favorisant des réactions métaboliques spécialisées. Des processus incompatibles peuvent ainsi se dérouler simultanément dans une cellule.
Ces membranes, importantes dans l’organisation complexe de la cellule se composent en général d’une double couche de phosphoglycérides et d’autres lipides associés à diverses protéines directement enchâssées dans la double couche ou fixées à la surface de cette membrane. Toutefois, chacune de ces membranes comporte une composition en lipides et protéines adaptée à ses fonctions spécifiques, à l’exemple de la membrane interne de la mitochondrie qui comporte en son sein une quantité importante d’enzymes de la respiration cellulaire.
Notons enfin que les différences entre cellule animale et cellule végétale sont bien moindre que les différences entre cellules procaryotes et eucaryotes, quoique non négligeables.
Les différences essentielles sont :
Vacuole centrale, chloroplastes, paroi cellulaire et plasmodesmes présents chez la cellule végétale et non chez la cellule animale.
Cellule animale
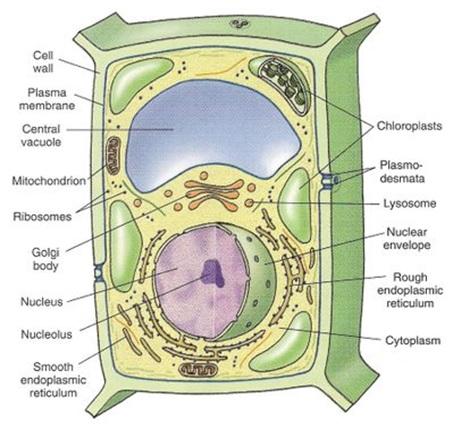
Cellule végétale
VI.4 L’information génétique de la cellule eucaryote est contenue dans le noyau et utilisée par les ribosomes pour fabriquer des protéines.
Le noyau héberge la majorité de l’ADN cellulaire et les ribosomes fabriquent les protéines à partir de l’information codée dans l’ADN.
4.1 Le noyau, porteur de l’information génétique de la cellule
En général un seul noyau par cellule contenant la plupart des gènes qui régissent la cellule eucaryote, les autres se trouvant dans les mitochondries ou les chloroplastes. Son diamètre moyen de 5 µm permet que ce soit l’organisme le mieux visible de la cellule eucaryote. Il est entouré d’une membrane double appelée membrane nucléaire.
Les deux membranes nucléaires sont elles-mêmes constituées d’une double couche de phosphoglycérides associée à des protéines. Ces deux membranes sont séparées par un espace de 20 à 40 nm environ. L’enveloppe nucléaire renferme des milliers de pores. Au niveau de ces pores, les deux membranes se rejoignent. Ces pores comportent également un complexe protéique de quelques dizaines de protéines. Ce complexe du pore nucléaire mesure environ 100 nm de diamètre et règle le passage de macromolécules et particules. La lamina nucléaire est un entrelacement de filaments protéiques tapissant l’enveloppe interne nucléaire et maintenant la forme du noyau. On a également découvert une matrice nucléaire constituée d’un réseau de fibres qui s’étend dans le noyau.
Le noyau contient un complexe d’ADN et de protéines que l’on appelle chromatine, cette dernière a une très grande affinité pour certains colorants et elle apparaît à la coloration comme un amas diffus, en microscopie photonique ou électronique. Au moment où la cellule s’apprête à se diviser, les minces filaments de chromatine se condensent et s’épaississent en chromosomes. Chaque espèce eucaryote comporte un nombre caractéristique de chromosomes. Chez l’espèce humaine, les chromosomes sont au nombre de 46 par noyau cellulaire, exception faite des cellules germinales qui en contiennent 23. Chez la drosophile, les cellules somatiques comportent 8 chromosomes et les cellules germinales en contiennent 4.
Entre les périodes de division cellulaire, la structure la plus visible du noyau est le nucléole. Celui-ci apparaît en microscopie électronique comme une masse compacte de granules et de fibres associée à la chromatine. Une forme d’ARN, l’ARN ribosomial est synthétisé dans le nucléole à partir de l’ADN du nucléole. Des protéines importées du cytoplasme sont associées à l’ARNr pour former de grandes et de petites sous unités ribosomiques ; Ces sous unités entrent dans le cytoplasme par les pores nucléaires. Là, une grande sous unité et une petite s’associent pour former les ribosomes. Le noyau comporte parfois plus de 2 nucléoles en fonction de l’espèce et du stade du cycle cellulaire. Ils auraient également un rôle dans le contrôle du vieillissement cellulaire.
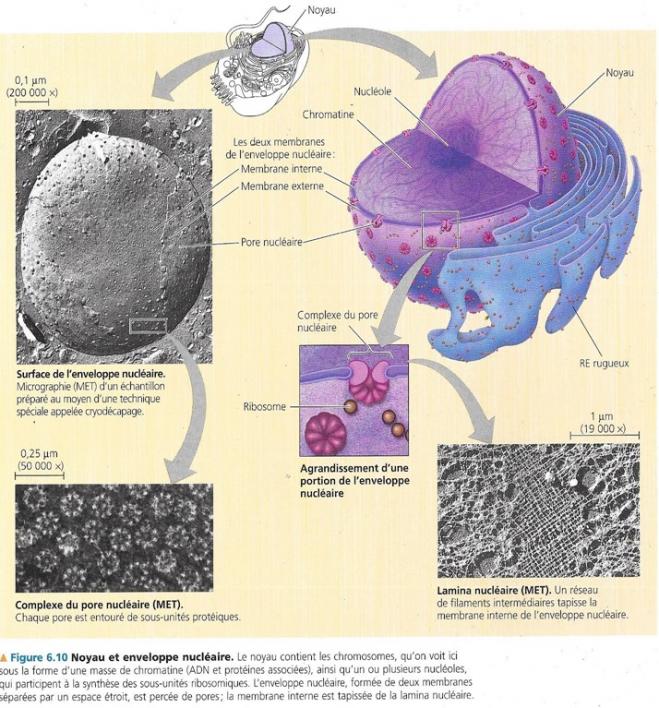
Nous verrons plus loin, que c’est au niveau du noyau que l’ARNmessager est produit d’après les directives de l’ADN. L’ARNm est ensuite expédié dans le cytoplasme par les pores nucléaires. Les molécules d’ARN messager seront traduites en protéines sous structure primaire par les ribosomes, conformément à leur message génétique. Ces deux processus portent respectivement les noms de transcription et de traduction.
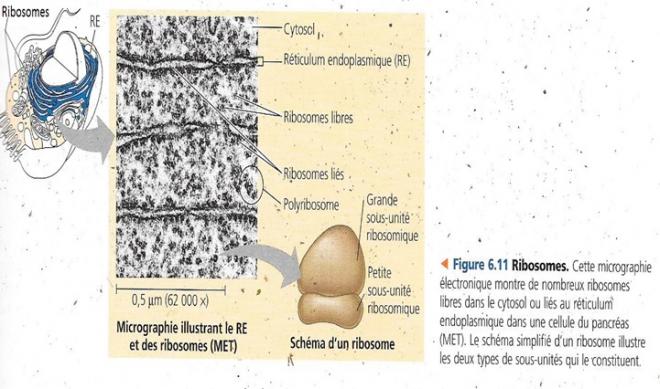
4.2 Les ribosomes : des usines à protéines
Les ribosomes, particules constituées d’ARN et de protéines sont les organites qui synthétisent les protéines. Les cellules qui synthétisent beaucoup de protéines sont caractérisées par un grand nombre de ribosomes, comme la cellule pancréatique humaine qui en comporte quelques millions. Dans ce cas, le nucléole est également volumineux. Les protéines sont assemblées à partir de l’activité de deux types de ribosomes : d’une part les ribosomes liés (à la surface du noyau ou de l’appareil de Golgi) et les ribosomes libres suspendus dans le cytosol. Les ribosomes libres seuls n’ont pas d’activité, ce n’est que lorsqu’ils s’assemblent en polyribosomes qu’ils produisent des protéines essentiellement destinées à une activité dans le cytosol (enzymes nécessaires aux premières étapes du catabolisme des glucides). Les ribosomes liés synthétisent quant à eux des protéines destinées à être insérées dans les membranes ou dans des organites comme les lysosomes , ou encore à être exportées (cellules sécrétrices). Les cellules spécialisées dans la sécrétion de protéines comme les cellules pancréatiques ou des enzymes diverses du tube digestif comportent une grande quantité de ribosomes : liés ou libres, les ribosomes ont la même structure, ils sont interchangeables et se répartissent en fonction des besoins de la cellule.
VI.5 Le réseau intracellulaire de membranes dirige la circulation des protéines et accomplit des fonctions métaboliques dans la cellule.
5.1 Introduction
Existence d’un réseau intracellulaire de membranes accomplissant diverses tâches dans la cellule :
- Synthèse de protéines
- Transport de celles-ci vers d’autres membranes ou organites
- Transport vers l’extérieur de la cellule
- Métabolisme et mouvement des lipides
- Détoxication de poisons
Ces membranes sont liées les unes aux autres de deux façons :
- Elles se prolongent l’une l’autre
- Elles échangent des portions d’elles même par l’intermédiaire de vésicules minuscules (sacs membraneux).
Ces membranes n’ont cependant pas toutes la même structure et fonction, elles diffèrent essentiellement :
- Par leur épaisseur
- Par leur composition moléculaire
- Par le type de réactions chimiques auxquelles participent les protéines des membranes qui peuvent changer au cours de la vie de la cellule.
Le réseau intracellulaire de membranes se compose de :
- L’enveloppe nucléaire
- Le réticulum endoplasmique
- L’appareil de Golgi
- Les lysosomes
- Les peroxysomes
- Diverses vacuoles
Ce réseau est également lié à la membrane plasmique qui n’est cependant pas intracellulaire.
5.2 Le réticulum endoplasmique est une usine biosynthétique.
Il s’agit d’un labyrinthe membraneux faisant réseau dans les cellules eucaryotes. Il représente au moins la moitié de toute la substance membraneuse de la cellule.
5.2.1 Morphologie
Il est constitué morphologiquement :
- Un réseau de tubules
- Des sacs membraneux appelés citernes.
Bien sûr liés en continuité.
La membrane du RE isole le cytosol du contenu des citernes
Sa membrane étant en continuité avec l’enveloppe nucléaire, le contenu des citernes est en contact directe avec l’espace ménagé entre les deux membranes de l’enveloppe nucléaire.
Le RE se partage en deux régions comportant certaines différences moléculaires et fonctionnelles :
- Le RE lisse : qualifié ainsi parce qu’il ne porte pas de ribosomes sur sa face cytoplasmique
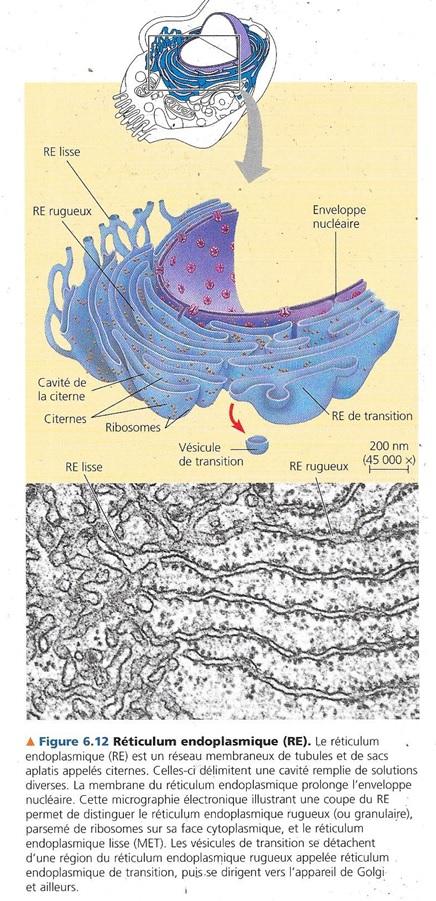
- Le RE granuleux ou rugueux : qualifié ainsi car il a un aspect granulaire en microscopie électronique, en effet, il est parsemé à sa surface externe ou cytoplasmique de ribosomes. Remarquons qu’en se prolongeant en continuité avec l’enveloppe nucléaire, on trouve également des ribosomes à la surface du noyau.
5.2.2. Fonctions du réticulum endoplasmique lisse
a) Fonctions métaboliques
- Synthèse des lipides
- Métabolisme des glucides
- Détoxication des médicaments
- Détoxication des drogues et des poisons
b) activité enzymatique particulière
- Synthèse enzymatique des lipides : graisses, phosphoglycérolipides et des stéroïdes comme dans les cellules animales, notamment, les hormones sexuelles des vertébrés et les hormones stéroïdes des glandes surrénales. Les cellules qui synthétisent ces hormones, par exemple dans les testicules et les ovaires sont riches en réticulum endoplasmique lisse ( lien structure-fonction)
- Activité enzymatique de détoxication de drogues, médicaments, poisons, particulièrement dans le foie. La détoxication consiste souvent un l’ajout d’un groupement hydroxyle à la molécule toxique, ce qui augmente sa solubilité et favorise son élimination. C’est le cas du sédatif phénobarbital. En fait toutes ces diverses activités de détoxication sur médicaments, drogues, poisons, alcool…provoque l’accroissement du REL cellulaire et du taux de ses enzymes de détoxication, l’homme acquiert ainsi un plus grand pouvoir de détoxication et devient plus tolérant aux produits en question. De cette manière ce pouvoir se généralise par le large spectre d’action de certaines enzymes de détoxication et la tolérance à l’un des produits (barbituriques par exemple) peut entraîner la tolérance à d’autres produits (antibiotiques ou autres médicaments par exemple).
c) Stockage de calcium dans les citernes du REL
Le REL emmagasine des ions calcium, en particulier dans les cellules musculaires. Dans ces cellules, une membrane spécialisée du REL extrait les ions calcium du cytosol et les accumule dans ses citernes. Lorsqu’un influx nerveux arrive au niveau de la cellule musculaire, le calcium traverse la membrane du REL, se libère dans le cytosol et déclenche la contraction musculaire. Cette libération de calcium peut, dans d’autres types cellulaires, provoque d’autres réactions.
5.2.3 Fonctions du REG ou rugueux
Les ribosomes du REG produisent les protéines sécrétées par beaucoup de cellules spécialisées (ex : insuline des cellules pancréatiques).
Mécanisme de sécrétion :
Un ribosome lié synthétise une chaîne polypeptidique à entrée de la chaîne polypeptidique dans la lumière du REG par un pore à repliement de la protéine en conformation native -à Grâce à l’activité d’enzymes enchâssées dans la membrane du REG, un petit polysaccharide est attaché par covalence à la protéine et devient une glycoprotéine.
La plupart des protéines de sécrétions sont des glycoprotéines
Le réticulum isole les protéines sécrétées de celles produites par les polyribosomes du cytosol qui vont rester dans le cytosol, cet isolement se matérialise par la création de vésicules de sécrétion, elles émanent d’une région appelée réticulum endoplasmique de transition, Dans ce cadre le réticulum endoplasmique rugueux peut également accroître la membrane plasmique en y ajoutant des protéines et des phosphoglycérolipides. Ces protéines sont issues de l’activité des ribosomes et s’insèrent dans la membrane par leurs parties hydrophobes. Les phosphoglycérolipides sont assemblés, quant à eux à partir de matériaux du cytosol par des enzymes.
de la membrane du REG. Le nouveau matériel peut aussi être transféré à d’autres organismes contenant des membranes.
5.3 L’appareil de Golgi est un centre d’expédition et de réception.
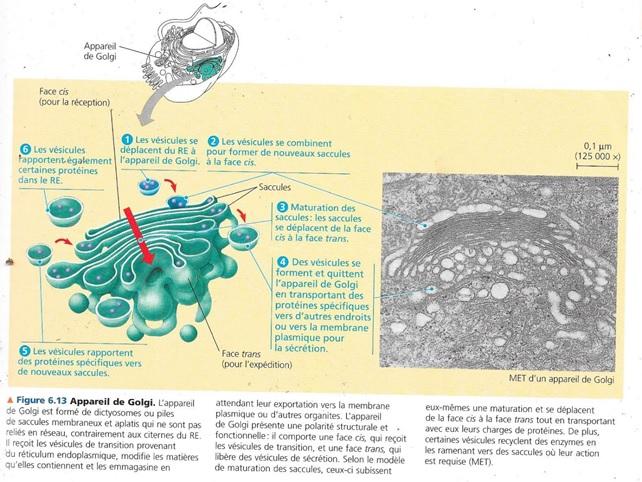
5.3.1 Introduction.
Les vésicules de sécrétion issues du REG se dirigent en général vers l’appareil de Golgi. On peut comparer celui-ci à :
- Centre de fabrication
- Centre d’entreposage
- Centre de triage
- Centre d’expédition
Les produits du RE y sont modifiés, entreposés, puis envoyés vers différentes destinations. Il va de soi que l’appareil de Golgi est particulièrement important et étendu dans les cellules sécrétrices. Cet appareil se situant en général à proximité du noyau.
5.3.2 Morphologie
Il est constitué d’un ensemble d’empilements de saccules membraneux aplatis appelés dictyosomes. Chaque cellule peut en contenir plusieurs centaines. Les membranes concernées séparent le contenu de AG de celui du cytosol. Les vésicules de sécrétions qui se concentrent à proximité de AG véhiculent des matières entre celui-ci et d’autres structures cellulaires.
L’appareil de golgi est caractérisé par une polarité :
- L’appareil de golgi possède une polarité, la membrane des saccules située à l’opposé des dictyosomes n’ont ni la même composition moléculaire, ni la même épaisseur. Ces pôles s’appellent face cis et face trans et ont pour fonction respective de recevoir et d’expédier les matières
- La face cis est convexe et située près du RE de transition, elle en reçoit les vésicules de transition. Si tôt libérées, ces vésicules fusionnent avec les membranes cis d’un dictyosome et y incorporent leur membrane et leur contenu.
- La face trans concave donne naissance à des vésicules de sécrétion qui s’acheminent vers d’autres sites.
5.3.3 Fonctions de l’appareil de Golgi.
En général, les contenus des vésicules de transition issus du RE subissent des modifications dans AG au long de leur transfert de la face cis à la face trans et les protéines et phosphoglycérolipides de la membrane des vésicules peuvent y subir des modifications :
- Au départ du RE, les parties glucidiques des glycoprotéines qui y sont assemblées sont de même nature, en fin de RE ces parties glucidiques commencent à être modifiées.
- Une fois transférées dans l’AG par sa face cis, ces glycoprotéines sont encore modifiées, ce dernier déloge certains monomères des polysaccharides et les remplace par d’autres, il produit ainsi des glucides différents donc des glycoprotéines différentes de ce qu’ils étaient à leur origine.
- L’appareil de Golgi fabrique certaines macromolécules, comme certains nombreux polysaccharides sécrétés par les cellules comme la lignine et d’autres polysaccharides végétaux insérés à la cellulose des parois cellulaires végétales, la cellulose étant synthétisée par des enzymes de la membrane cellulaire qui les transfère à la face externe de la membrane.
- Les produits de l’appareil de Golgi destinés à la sécrétion sont isolés dans des membranes, constituant ainsi des vésicules de sécrétion qui fusionneront avec la membrane plasmique.
L’appareil de Golgi élabore et affine ses produits par étapes :
- Celles-ci correspondent aux différents saccules entre la face cis et la face trans d’un dictyosome qui contiennent des enzymes différentes
- On a longtemps considéré que les modifications se faisaient de saccule par l’intermédiaire du transport par des vésicules de sécrétion, c’est le modèle vésiculaire
- On a tendance actuellement à considérer un modèle appelé modèle de maturation des saccules dans lequel l’AG est une structure dynamique dans laquelle les vésicules transportent mais modifient également les produits de sécrétion.
Avant d’émettre les vésicules de sécrétion, par sa face trans, l’appareil de Golgi doit trier ses produits et déterminer leur destination :
- Des « étiquettes moléculaires » comme des groupements phosphate sont apposées sur les produits et jouent un peu le rôle d’un code postal dans une adresse.
- On croit que les vésicules de sécrétion provenant de l’AG portent des molécules externes qui reconnaissent des sites récepteurs spécifiques à la surface des organites ou sur la membrane plasmique qui permet de les cibler.
5.4. Les lysosomes : des compartiments destinés à la digestion.
5.4.1 Morphologie et mécanismes de fonctionnement
- Sac membraneux contenant une dizaine d’enzymes hydrolytiques qui digèrent toutes sortes de macromolécules. Les enzymes lysosomiales ont une activité qui est optimale en milieu acide (pH5). Si un lysosome a une fuite ou s’il se désagrège, il répand son contenu dans le cytosol, mais comme le pH cytosolique est proche de la neutralité, les enzymes sont inactivées.
Par-contre si une quantité suffisante de lysosomes sont détruits, leur contenu peut détruire la cellule. Ceci montre l’importance de la compartimentation dans la cellule. Le lysosome met la cellule à l’abri des dommages que pourraient provoquer les enzymes hydrolytiques si elles circulaient librement dans le cytosol.
- Les enzymes hydrolytiques et la membrane du lysosome ont pour origine le Réticulum rugueux endoplasmique puis transférées séparément à la face cis de l’appareil de Golgi dans lequel leur traitement se poursuit.
- Il semble que certains lysosomes sont issus du bourgeonnement de la face trans de l’appareil de Golgi
- les protéines de la face interne du lysosome ainsi que les enzymes hydrolytiques sont protégées de l’autodestruction grâce à leur conformation tridimensionnelle qui protège leurs liaisons vulnérables de l’activité enzymatique.
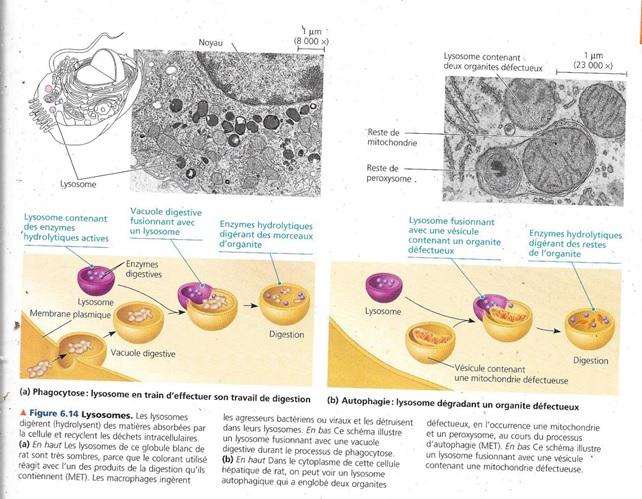
La fonction digestive des lysosomes s’opère dans diverses circonstances :
- Certaines cellules se nourrissent par endocytose. C’est un processus par lequel la cellule ingère des nutriments en laissant passer ces nutriments en formant des vacuoles. Chacune de celles-ci fusionne avec un lysosome qui en digère le contenu grâce à ses enzymes. Les produits de la digestion sont des glucides simples, des acides aminés et d’autres monomères transitent dans le cytosol et fournissent à nouveau de la matière et de l’énergie à la cellule.
- Certaines cellules humaines comme les macrophages du système immunitaire détruisent les bactéries, les virus et des substances étrangères par phagocytose. C’est un processus par lequel la cellule se déforme en tout ou en partie, afin d’entourer complètement le corps étranger. Ce dernier se trouve ainsi dans une vacuole digestive ;
- Le lysosome peut aussi recycler les matières organiques intracellulaires par autophagie. Au cours de ce processus, un organite défectueux ou endommagé ou encore un peu de cytosol s’entourent d’une membrane et forment une vésicule qui fusionne avec un lysosome. Ce dernier à l’aide de ses enzymes dégrade la matière organique ingérée en monomères.
L’absence de certaines enzymes lysosomiales constitue les maladies de surcharge qui sont un groupe de troubles correspondant à un manquement du métabolisme lysosomial, ces maladies sont héréditaires. Il y a alors engorgement des lysosomes par des substances inutilisables. L’absence d’une lipase ou son inactivité provoque l’accumulation de lipides dans les lysosomes et finalement dans les cellules nerveuses comme dans le cas de la maladie de Tay-Sachs qui entrave le fonctionnement de l’encéphale, ces maladies sont rares.
5.5 Les vacuoles : des compartiments d’entretien divers.
Une cellule de plante ou de champignon peut avoir une ou plusieurs vacuoles ; Les vacuoles pratiquent l’hydrolyse, sont donc semblables aux lysosomes, mais elles accomplissent aussi d’autres fonctions : Nous avons parlé des vacuoles digestives, mais leur rôle ne s’arrête pas là :
- Beaucoup de protistes d’eau douce possèdent une ou plusieurs vacuoles pulsatiles afin de maintenir une concentration appropriée de sels et d’autres molécules.
- Les cellules végétales matures contiennent généralement une grande vacuole centrale entourée d’une membrane appelée tonoplaste qui fait partie du réseau intracellulaire de membranes. C’est la fusion de vésicules de sécrétion du RE et de l’AG qui constitue la vacuole. Le tonoplaste a une perméabilité sélective comme toutes les membranes du réseau intracellulaire de membranes, ce qui explique la différence entre la composition de la solution cytosolique et la solution intérieure à la vacuole.
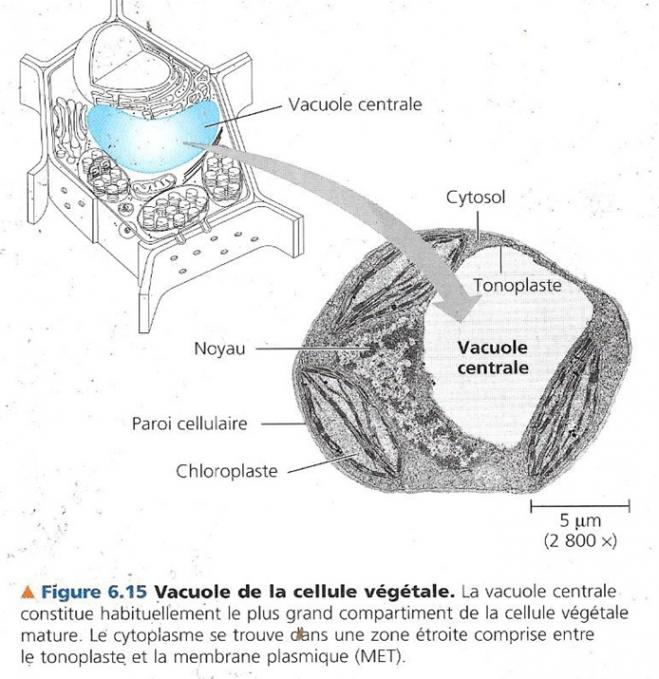
La vacuole végétale est polyvalente :
- Elle sert à emmagasiner des composés organiques cruciaux à l’exemple de l’accumulation de protéines dans les cellules nutritives des graines produites par une plante.
- Elle peut également constituer un réservoir d’ions inorganiques comme les ions potassium et chlorure ;
- Elle permet aussi d’isoler les sous-produits du métabolisme qui s’ils s’accumulent dans le cytosol deviennent toxiques.
- Elles peuvent contenir des pigments tels que les pigments rouges ou bleus qui attirent les insectes pollinisateurs vers les pétales.
- Les vacuoles contiennent parfois des substances toxiques ou désagréables au goût pour les prédateurs.
Les vacuoles jouent également un rôle dans la croissance végétale, le fait qu’elles absorbent de l’eau permet à la cellule de s’allonger sans devoir produire du cytoplasme. La croissance de la vacuole coince le cytosol entre la membrane plasmique et le tonoplaste, c’est ainsi que le rapport surface volume de la membrane par rapport au cytosol reste élevé même pour une cellule de grande taille.
- Elle permet aussi d’isoler les sous-produits du métabolisme qui s’ils s’accumulent dans le cytosol deviennent toxiques.
- Elles peuvent contenir des pigments tels que les pigments rouges ou bleus qui attirent les insectes pollinisateurs vers les pétales.
- Les vacuoles contiennent parfois des substances toxiques ou désagréables au goût pour les prédateurs.
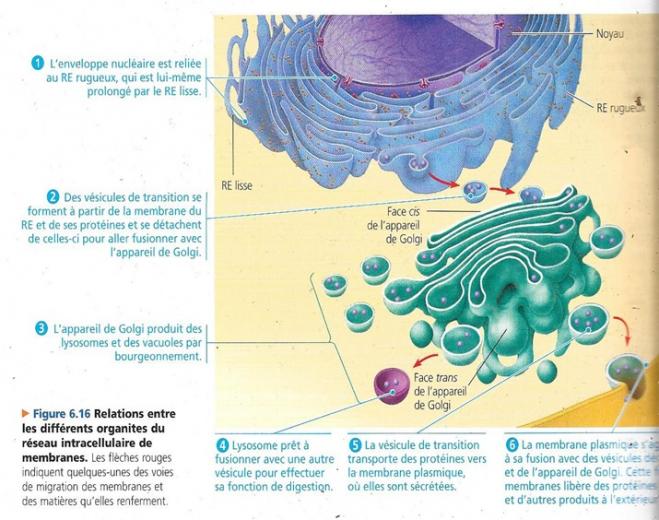
Le réseau intracellulaire de membranes se charge de la circulation de lipides et de protéines dans ses divers organites (vacuole, lysosomes AG RG …). Les membranes de ces organites n’ont pas toutes la même composition moléculaire quoique basée sur la même structure de base. En conséquence, les fonctions métaboliques sont différentes. Le réseau intracellulaire de membranes est un réseau complexe constituant une entité ayant un rôle crucial dans les conversions d’énergie ayant lieu dans la cellule.
VI.6 : Mitochondries et chloroplastes convertissent l’énergie d’une forme à une autre.
6.1 Introduction
Mitochondries et chloroplastes des cellules eucaryotes transforment l’énergie puisée dans l’environnement (rayons solaires, nutriments) en des formes utilisables par la cellule.
- Les mitochondries sont le site de la respiration cellulaire aérobie qui permet de produire de l’ATP à partir du dioxygène et des nutriments ( glucides, lipides, autres substances…).
- Les chloroplastes, organites propres aux cellules végétales et aux algues sont le site de la photosynthèse. Ce processus métabolique convertit l’énergie solaire en énergie chimique Cette énergie sert ainsi à la synthèse de glucides à partir de dioxyde de carbone et d’eau.
- Bien que recouverts d’une membrane, les chloroplastes et les mitochondries ne font pas partie du réseau intracellulaire de membranes. Ils possèdent au moins deux membranes pour séparer leur espace interne du cytosol. Leurs protéines membranaires proviennent de leurs propres ribosomes et des polyribosomes libres du cytosol (et non ceux du REr).
- Leur espace intérieur contient une petite quantité d’ADN et leurs permet de synthétiser eux même une petite partie de leurs protéines, la plus grande partie provenant de la traduction dans le cytosol d’ARNm issus de l’ADN du noyau.
- Mitochondries et chloroplastes sont des organites semi-autonomes qui croissent et se reproduisent à l’intérieur de la cellule indépendamment de la division cellulaire. L’ADN qu’il contiennent est transmis aux organites qui sont issus de leur division.
6.2 Les mitochondries, convertisseurs d’énergie chimique.
6.2.1 Généralités
Présentes presque dans toutes les cellules eucaryotes : celles des végétaux, des animaux, des eumycètes, des protistes. Certaines cellules n’en contiennent qu’une, de grande taille, mais en général elles en contiennent des centaines voire des milliers. Leur nombre dépend de l’activité métabolique de la cellule. Ainsi, les cellules contractiles ou très mobiles proportionnellement au volume une quantité plus importante de mitochondries que les cellules moins actives. Leur longueur varie entre 1 et 10 µm. Par des techniques de visualisation en accéléré sur des cellules vivantes, on peut voir que les mitochondries se déplacent, se déforment, se divisent en deux. Les mitochondries ne sont donc pas statiques comme on peut l’imaginer en regardant des microphotographies électroniques de cellules fixées.
6.2.2 Morphologie
- Membrane double constituant l’enveloppe, chaque membrane est constituée d’une double couche de phospoglycérolipides dans laquelle s’insère un assemblage unique de protéines.
- La membrane externe est lisse et la membrane interne est hérissée de crêtes tournées vers l’intérieur
- La membrane interne divise la mitochondrie en deux compartiments : en premier lieu un espace intermembranaire entre membrane interne et membrane externe et en second lieu un espace intérieur à la membrane interne appelé matrice mitochondriale.
- plusieurs étapes métaboliques ont lieu dans la matrice, comme le cycle de Krebs … celle-ci contient de l’ADN, des ribosomes et diverses enzymes.
- d’autres protéines nécessaires à la respiration cellulaire dont celle qui réalise la synthèse de l’ATP, sont intégrées à la membrane interne.
- Les crêtes de la membrane interne permettent d’augmenter sa surface sans crête d’un facteur 5 et de donner plus d’espace pour la respiration cellulaire (phosphorylation oxydative).
6.3 Les chloroplastes, capteurs d’énergie lumineuse.
6.3.1 Généralités.
Les chloroplastes font partie d’une famille d’organites de la cellule végétale appelés plastes :
- Amyloplastes (ou leucoplastes) : plastes incolores renfermant de l’amidon, présents dans les racines et les tubercules
- Chromoplastes : ils synthétisent des pigments qui donnent aux fruits et aux fleurs leurs couleurs particulières orangées et jaunes.
- Chloroplastes : contiennent le pigment vert appelé chlorophylle de couleur verte, des enzymes et les molécules nécessaires à la production de glucides lors de la photosynthèse.
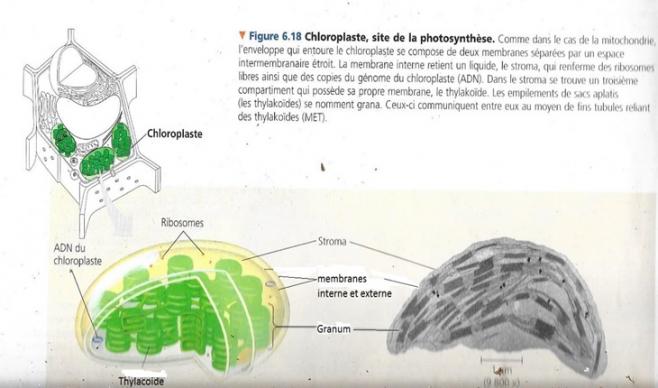
6.3.2 Morphologie
- Contenu isolé par une double membrane ménageant un espace intermembranaire mince
- Intérieur du chloroplastes contenant plusieurs empilements de sacs aplatis appelé thylacoïdes, chaque empilement est appelé granum ( grana au pluriel). C’est la membrane des thylacoïdes qui contient la chlorophylle.
- Le liquide où baignent les grana est appelé stroma et contient de l’ADN, des ribosomes et de nombreuses enzymes.
- L’ensemble membranaire de ce chloroplaste le divise en trois parties : l’espace intermembranaire, le stroma et l’espace intrathylacoïdien.
- Comme les mitochondries, les chloroplastes ne sont pas « statiques », ils se divisent, se déforment et se déplacent sur des « rails» constitués par le cytosquelette.
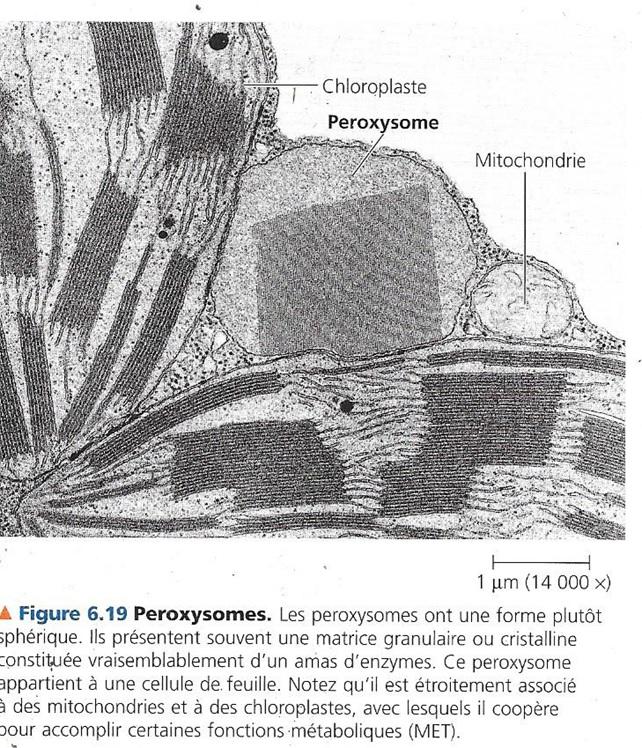
6.4 Les péroxysomes : des organites oxydatifs.
Compartiments métaboliques spécialisés délimités par une membrane simple.
Ils contiennent des enzymes qui transfèrent de l’hydrogène de divers substrats au dioxygène. Leur nom provient du nom d’un sous-produit de ce transfert, le peroxyde d’hydrogène (H2O2). Diverses fonctions :
- Utilisation du dioxygène pour décomposer des acides gras des lipides
- Détoxication de l’alcool et d’autres composés nocifs dans les cellules hépatiques en transférant l’hydrogène de ces substances au dioxygène.
Le peroxyde d’hydrogène est toxique mais une enzyme des lysosomes le transforme en eau. Les lysosomes contiennent donc une enzyme qui produit le peroxyde d’hydrogène et une enzyme qui le dégrade en eau. Certains peroxysomes spécialisés, appelés glyoxysomes contiennent une enzyme qui est capable de déclencher la conversion des acides gras en glucides, un processus qui produit de l’énergie et une source de carbone au jeune plant, jusqu’au moment où il est capable de produire lui-même ses glucides grâce à la photosynthèse.
Les peroxysomes ne proviennent pas d’un bourgeonnement du réseau intracellulaire de membranes. Il intègre des protéines produites principalement dans le cytosol, des lipides synthétisés dans le RE et des lipides produits par le peroxysome lui-même. Les peroxysomes se divisent en deux lorsqu’ils ont atteint une taille suffisante.
Date de dernière mise à jour : 21/12/2018
Ajouter un commentaire