

8. Métabolisme 2 : respiration cellulaire
RESPIRATION CELLULAIRE
- INTRODUCTION
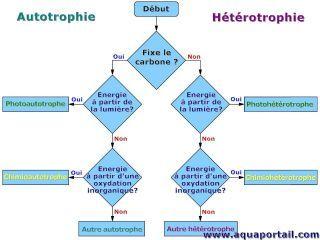
1. Autotrophes/hétérotrophes
Un hétérotrophe qualifie un organisme qui assure sa subsistance en assimilant des substances organiques et incapable de produire ces substances organiques à partir de matière minérale. Sa source de carbone est la matière organique, car un hétérotrophe ne peut pas fixer le carbone. Par définition, hétérotrophe s'oppose à autotrophe.
Schéma de différenciation d'un hétérotrophe et d'un autotrophe:
Les hétérotrophes qualifient des êtres vivants ayant besoin d'aliments organiques pour assurer leurs fonctions métaboliques. Tous les animaux, certaines plantes vasculaires parasites (phytoparasites, plantes parasites), les champignons et certaines bactéries sont hétérotrophes.
Autrement exprimé, un organisme hétérotrophe se distingue par un mode de nutrition dans lequel les composés organiques utilisés dans le métabolisme sont obtenus par consommation des corps et des produits d'autres organismes.
Les hétérotrophes peut être répartis en fonction de la façon dont ils obtiennent de l'énergie; si l'hétérotrophe utilise la lumière pour l'énergie, alors il est considéré comme un photohétérotrophe, tandis que si l'hétérotrophe utilise l'énergie chimique, il est considéré comme un chimiohétérotrophe.
Les hétérotrophes contrastent avec les autotrophes, tels que les plantes et les algues, qui peuvent utiliser l'énergie de la lumière solaire (photoautotrophes) ou des composés inorganiques (lithoautotrophes) pour produire des composés organiques tels que des glucides, des graisses et des protéines à partir du dioxyde de carbone inorganique. Ces composés de carbone réduits peuvent être utilisés comme source d'énergie par l'autotrophe et fournissent l'énergie dans les aliments consommés par les hétérotrophes. Quatre-vingt-cinq pour cent ou plus de tous les types d'organismes vivants sont hétérotrophes.
En biologie, ce qui est hétérotrophique est relatif à l'hétérotrophie.
2 Utilisation de l'énergie
Vivre c’est fournir du travail
organismes autotrophes : tirent leur énergie du rayonnement solaire pour leur fonctionnement (travail) et la synthèse de molécules organiques à partir de l'eau et du dioxyde de carbone : photosynthèse
organismes autotrophes : tirent leur énergie de la consommation de molécules organiques synthétisées par d'autres organisme , par décomposition de ces nutriments : respiration et fermentation.
|
Plantes --> herbivores --> énergie = E utilisable et pertes sous forme de chaleur ou Plantes --> herbivores(-->énergie) --> carnivores --> énergie = E utilisable et pertes sous forme de chaleur) |
Energie utilisable ---> Cellules---> travail : tâches essentielles :
- Assemblage de polymères
- Transport de substances au travers des membranes
- Déplacement
- Reproduction… ;
On voit que l’énergie accumulée à la base dans les molécules organiques des plantes vient de l’énergie solaire. L’énergie entre dans l’écosystème sous forme de lumière qui ressort sous forme de chaleur
3. sites cellulaires de la photosynthèse et de la respiration
La photosynthèse a lieu en général dans des organites cellulaires appelés chloroplastes chez les végétaux chlorophylliens
La respiration a lieu dans des organites cellulaires appelées mitochondries chez les animaux
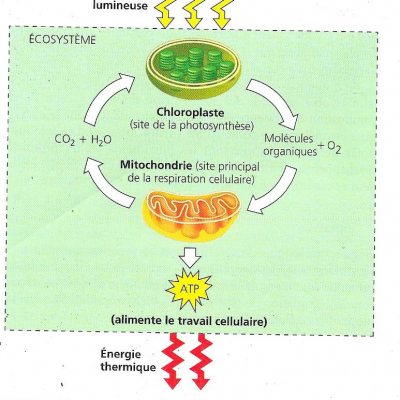
On voit sur le schéma que les substances chimiques essentielles à la vie quand-à elles sont recyclées.
4. Produits de la photosynthèse et de la respiration;
La photosynthèse génère :
- du dioxygène
- des molécules organiques
Ces molécules servent de combustible pour la respiration cellulaire des mitochondries chez les organismes eucaryotes (certains organismes photosynthétiques pratiquent en plus la respiration).
La respiration cellulaire décompose ces molécules pour produire de l’ATP.
La respiration cellulaire génère :
- du dioxyde de carbone
- de l’eau
Ces molécules sont les matières premières de la photosynthèse.
L’objet du présent chapitre est d’étudier la respiration qui en produisant le dioxyde de carbone et de l’eau à partir de substances organiques et du dioxygène produit de l’ATP. L’ATP est la substance qui alimente la majeure partie du travail cellulaire.
B; LES VOIES CATABOLIQUES GENERENT DE L’ENERGIE EN OXYDANT LES MOLECULES ORGANIQUES
Processus essentiels de la respiration cellulaire et de ses voies
- Les voies cataboliques et la production d’ATP
Comme nous l’avons vu précédemment, toute substance chimique, comme les molécules organiques, contient de l’énergie qui peut être libérée à différentes fins.
Cette énergie contenue dans les substances organiques provient de l’organisation de leurs atomes. Dans l’organisme, des enzymes modifient l’organisation des substances organiques (dégradation) complexes qui contiennent beaucoup d’énergie potentielle. Cette dégradation génère des produits plus simples qui contiennent moins d’énergie. Une certaine quantité d’énergie est ainsi rendue disponible. Celle-ci servira à fournir du
travail, et le reste se dissipera sous forme de chaleur. Ce processus correspond aux voies cataboliques.
Nous distinguons trois de ces voies :
- La fermentation : qui dégrade partiellement le glucose en l’absence de dioxygène et de chaîne de transport d’électrons
- La respiration cellulaire aérobie : voie catabolique la plus répandue et la plus efficace, elle utilise le dioxygène et les combustibles organiques comme réactifs. Elle utilise une chaîne transporteuse d’électron. Les mitochondries contiennent la majeure partie du matériel métabolique nécessaire à la respiration aérobie d’une cellule eucaryote.
- La respiration cellulaire anaérobie : plus marginale s’effectue en l’absence d’oxygène, mais nécessite une chaîne transporteuse d’électrons.
Bien que les mécanismes soient différents, on peut comparer la respiration cellulaire aérobie à la combustion de l’essence dans un moteur. A l’intervention de l’oxygène, le combustible (essence/nutriments) sera transformé en dioxyde de carbone et d’eau avec libération d’énergie. Selon :
Composés organiques + dioxygène ---> CO2 + eau + Energie
Les glucides, les lipides et les protéines peuvent servir, une fois traités, de combustible.
Nous traiterons le problème des étapes de la respiration cellulaire aérobie en décrivant la dégradation du glucose (C6H12O6).
C6H12O6 + 6 O2 à 6 CO2 + 6 H2O + Energie (ATP et Chaleur)
Une variation d’énergie libre accompagne cette réaction globale : ΔG = -2870 kJ/mol
Les produits ont donc moins d’énergie que les réactifs et la réaction est spontanée.
travail, et le reste se dissipera sous forme de chaleur. Ce processus correspond aux voies cataboliques.
Nous distinguons trois de ces voies :
- La fermentation : qui dégrade partiellement le glucose en l’absence de dioxygène et de chaîne de transport d’électrons
- La respiration cellulaire aérobie : voie catabolique la plus répandue et la plus efficace, elle utilise le dioxygène et les combustibles organiques comme réactifs. Elle utilise une chaîne transporteuse d’électron. Les mitochondries contiennent la majeure partie du matériel métabolique nécessaire à la respiration aérobie d’une cellule eucaryote.
- La respiration cellulaire anaérobie : plus marginale s’effectue en l’absence d’oxygène, mais nécessite une chaîne transporteuse d’électrons.
Bien que les mécanismes soient différents, on peut comparer la respiration cellulaire aérobie à la combustion de l’essence dans un moteur. A l’intervention de l’oxygène, le combustible (essence/nutriments) sera transformé en dioxyde de carbone et d’eau avec libération d’énergie. Selon :
Composés organiques + dioxygène à CO2 + eau + Energie
Les glucides, les lipides et les protéines peuvent servir, une fois traités, de combustible.
Nous traiterons le problème des étapes de la respiration cellulaire aérobie en décrivant la dégradation du glucose (C6H12O6).
C6H12O6 + 6 O2 à 6 CO2 + 6 H2O + Energie (ATP et Chaleur)
Une variation d’énergie libre accompagne cette réaction globale : ΔG = -2870 kJ/mol
Les produits ont donc moins d’énergie que les réactifs et la réaction est spontanée.
Les voies cataboliques ne prennent pas directement part au travail cellulaire (mouvement des flagelles, transports transmembranaires, …). Le lien entre catabolisme et travail cellulaire est assuré par un intermédiaire chimique : l’ATP.
Une fois l’ATP consommé pour le travail cellulaire : (ATP --> ADP + Pi + Energie), l’ATP doit être régénéré à partir d’ADP et de phosphate inorganique. C’est le rôle de la respiration cellulaire de pourvoir à cette fonction. La respiration alimente cette synthèse sur base de deux processus chimiques fondamentaux : l’oxydation et la réduction.
- Les réactions d’oxydation et de réduction : l’oxydoréduction.
Nous savons que l’oxydoréduction correspond à des transferts d’électron(s) d’un réactif à un autre.
La perte d’électron(s) correspond à une oxydation.
Le gain d’électron(s) correspond à une réduction.
Dans une réaction rédox l’oxydant est réduit et le réducteur est oxydé.
Voyons pour la formation de NaCl à partir de Na et de Cl et généralisons.
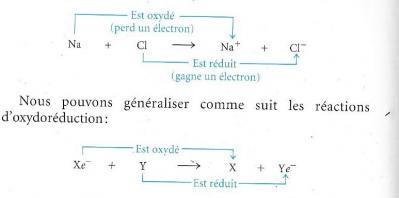
Ci-dessus :
- La substance X donne des électrons, c’est l’agent réducteur
- La substance Y capte des électrons, c’est l’agent oxydant
Dans une liaison électrovalente comme celle du NaCl, l’électron est totalement délocalisé vers l’agent le plus électronégatif. Dans le cas des liaisons covalentes, ce sont des charges partielles qui apparaissent suite à la délocalisation du doublet de liaison, ces δcharges sont responsables de la modification des nombres d’oxydation.
Examinons la réaction qui à partir du méthane et du dioxygène produit du dioxyde de carbone et de l’eau.
Dans le méthane, le carbone et l’hydrogène ont à peu prêt la même électronégativité, ils ont une affinité à peu près égale pour les électrons de valence. Les électrons sont ainsi mis en commun de manière égale et le carbone à ainsi la valence -4. Lors de la formation du dioxyde de carbone, l’oxygène étant plus électronégatif que le carbone, les 8 électrons des deux liaisons doubles sont délocalisés vers celui-ci, en quelques sortes arrachés au carbone, il apparaît ainsi 8 δ+ sur le carbone qui passe de l’étage d’oxydation – 4 à plus 4, le carbone du méthane est ainsi oxydé.
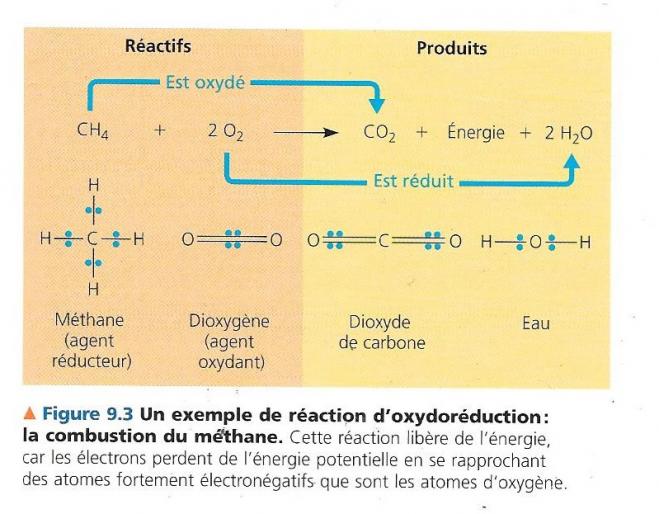
Dans la molécule de dioxygène, les électrons des deux doublets de liaison sont parfaitement partagés, l’étage d’oxydation de l’oxygène est égal à 0. Lors de la production de l’eau, les électrons des liaisons covalentes se rapprochent de l’oxygène qui joue ainsi le rôle de capteur d’électrons donc d’oxydant, il passe à l’étage d’oxydation -2.
Un atome électron de l’énergie lorsqu’il va d’un atome faiblement électronégatif vers un atome plus fortement électronégatif, ce mouvement est logique puisque spontanément l’électron va vers l’atome qui l’attire le plus. De cette manière il est plus fortement lié au noyau et dispose de moins d’énergie pour se mouvoir. Par conséquent une réaction d’oxydoréduction qui rapproche des électrons de l’oxygène libère de l’énergie chimique utilisable pour un travail. Comme l’oxydation du méthane par l’oxygène en dioxyde de carbone.
- L’oxydation des molécules organiques au cours de la respiration cellulaire.
La réaction d’oxydoréduction qui nous intéresse ici est l’oxydation du glucose qui nous intéresse ici est l’oxydation du glucose et d’autres molécules provenant du glucose.
Considérons l’équation de la respiration cellulaire aérobie, cette fois sous l’angle de l’oxydoréduction.

On a ici une oxydation du combustible (glucose) et réduction du dioxygène. Par l’oxydation, les électrons perdent de l’énergie potentielle et de l’énergie est libérée.
Les molécules organiques riches en hydrogène sont d’excellents combustibles car leurs liaisons comportent des électrons à haute énergie potentielle susceptibles de s’approcher de l’atome d’oxygène. Ainsi l’équation de la respiration cellulaire aérobie montre que l’hydrogène du glucose est transféré au dioxygène pour former de l’eau. Lors de cette réaction il y a libération d’énergie car le degré de covalence des électrons change quand l’hydrogène est transféré au dioxygène (la valeur de ΔG est négative).
Les nutriments énergétiques les plus courant, les glucides et les lipides, sont des réservoirs d’électrons associés à l’hydrogène.
La respiration cellulaire provoque l’oxydation du glucose qui libère de l’énergie utilisée à la production d’ATP.
Toute réaction, pour se produire doit passer la barrière énergétique d’activation. Cette barrière d’activation empêche un « raz de marée » d’électrons tendant à atteindre l’état énergétique le plus bas. Sans cette barrière, une molécule comme le glucose se combinerait spontanément au dioxygène. Lors de la combustion, on fournit l’énergie d’activation en déclenchant la combustion – c’est-à-dire l’oxydation rapide d’un combustible et la libération d’une énorme quantité d’énergie sous forme de chaleur -, chaque mole de glucose (soit 180 gr) brûle dans l’air en libérant 2870 kJ. La température corporelle seule n’est pas suffisamment élevée pour provoquer la combustion du glucose, mais si vous avalez du glucose, les enzymes de votre métabolisme se chargeront d’abaisser la barrière d’activation et le glucose sera oxydé lentement en une sérié d’étapes.
- Le transfert des électrons en une série d’étapes par l’entremise du NAD+ et de la chaîne de transport des électrons.
Il est difficile d’exploiter de manière efficace une énergie qui se libère en bloc (comme une combustion ou une explosion). Il ne servirait de rien que la respiration cellulaire aérobie oxyde le glucose en une seule étape explosive.
La respiration cellulaire se produit autrement :
Le glucose et les autres nutriments ou combustibles organiques sont dégradées en une série d’étapes toutes catalysées par une enzyme. Aux étapes clé, des atomes d’hydrogène sont arrachés au glucose. Comme c’est souvent le cas dans les réactions d’oxydation, les électrons sont transportés avec un proton autrement dit sous forme d’un atome d’hydrogène. Les atomes d’hydrogène ne rejoignent pas de manière directe le dioxygène et doivent généralement passer par un coenzyme appelé nicotinamide adénine dinucléotide ou NAD+, c’est un dérivé de la vitamine niacine, qui est accepteur d’électron et qui joue ainsi le rôle d’oxydant dans la respiration.
Le NAD+ capte les électrons du glucose et des autres molécules organiques par l’intervention d’enzymes déshydrogénase. Ces déshydrogénases retirent une paire d’atomes d’hydrogène (2 électrons et 2 protons) au substrat (un monosaccharide par exemple) l’oxydant du même coup. Les déshydrogénases apportent ensuite 2 électrons et un proton H+ au NAD+. Quant au proton restant il est libéré dans la solution environnante.

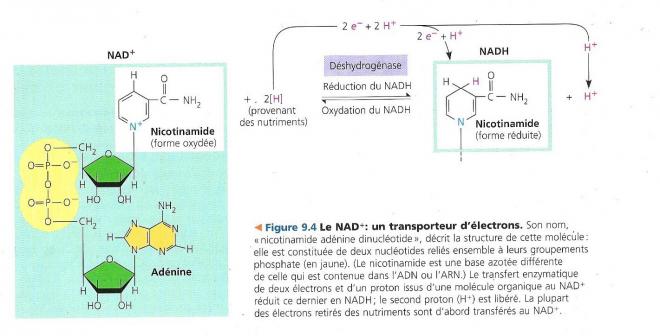
NADH signifie que NAD+ est réduit après acceptation d’un H+ et de deux électrons.
Les deux électrons négatifs et la charge positive du proton donnent globalement une charge négative qui va neutraliser la charge positive du NAD+àNADH
Le NAD+ est l’accepteur d’électrons le plus polyvalent dans la respiration cellulaire et il intervient dans plusieurs étapes d’oxydoréduction de la dégradation des monosaccharides. Les électrons perdent de l’énergie potentielle lorsqu’ils sont transférés par les déshydrogénases au NAD+. Cette perte est cependant faible, chaque fois qu’une môle de NADH + H+ est constituée pendant la respiration cellulaire aérobie, représente une réserve d’énergie qui pourra servir à produire de l’ATP, quand les électrons auront fini de descendre la pente énergétique menant du NADH + H+ au dioxygène.
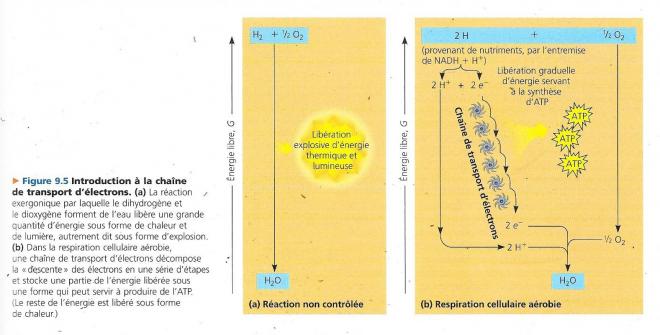
Ce qui nous importe maintenant est de connaître le mécanisme qui fait aboutir les électrons, issus des nutriments, du NADH au dioxygène. Nous allons commencer par une réaction beaucoup plus simple, celle qui produit de l’eau à partir de dihydrogène et de dioxygène. Mélangez ces deux gaz et fournissez l’énergie sous forme d’une étincelle. Ils se combineront de manière explosive. L’explosion produite correspond à la libération d’énergie survenant quand les électrons de l’hydrogène se rapprochent des atomes d’oxygène électronégatifs. La respiration cellulaire rapproche également de l’oxygène et de l’hydrogène pour former de l’eau, mais il y a deux différences :
- Dans la respiration cellulaire l’hydrogène provient de molécules organiques, plutôt que de dihydrogène.
- La respiration cellulaire aérobie utilise une chaîne de transfert d’électrons pour décomposer la « descente » vers le dioxygène en une série d’étapes libératrices d’énergie. Cette chaîne transporteuse d’électrons est constituée de plusieurs molécules (des protéines pour la plupart), insérée dans la membrane interne des mitochondries. Le NADH, riche en énergie apporte au sommet de la chaîne, les électrons retirés des nutriments. Au bas de la chaîne « pauvre en énergie », le dioxygène capture ces électrons en même temps que les protons, et de l’eau est formée.
Le transfert d’électrons du NADH + H+ au dioxygène est une réaction exergonique puisqu’il entraîne une variation d’énergie libre de -222kJ/mol environ. Mais cette énergie ne se libère pas en une seule étape. Les électrons descendent la chaîne en passant d’un transporteur à l’autre et en perdant à chaque étape une petite quantité d’énergie, jusqu’à ce qu’ils atteignent le dioxygène, le dernier accepteur qui se trouve au bas de la chaîne et qui a une très grande affinité pour les électrons. Lors de la descente de la chaîne transporteuse d’électrons, les transporteurs qui se rencontrent sont de plus en plus électronégatif, le dioxygène se trouvant au plus bas de cette pente.
|
Au cours de la respiration cellulaire aérobie, la majorité des électrons descendent la pente suivante : NutrimentàNADHà chaîne de transport d’électrons à dioxygène. |
|
Les électrons retirés des nutriments par le NAD+ dévalent donc une pente énergétique, celle de la chaîne de transport, jusqu’à ce qu’ils atteignent une position stable dans l’atome d’oxygène électronégatif. En d’autres termes, le dioxygène attire à lui les électrons de la chaîne de transport dans une cascade énergétique à la manière dont un corps subissant la loi de gravitation est attiré vers le bas. En résumé :
Etudions maintenant le processus entier de la respiration cellulaire aérobie.
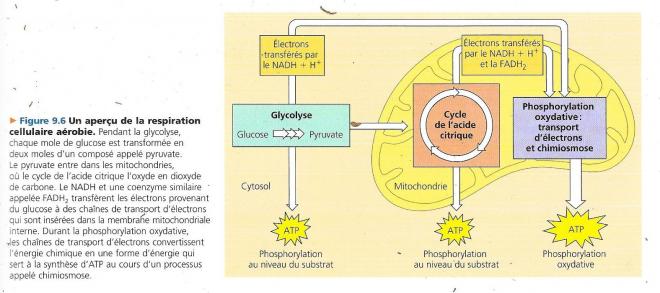
La respiration cellulaire aérobie comprend trois stades métaboliques
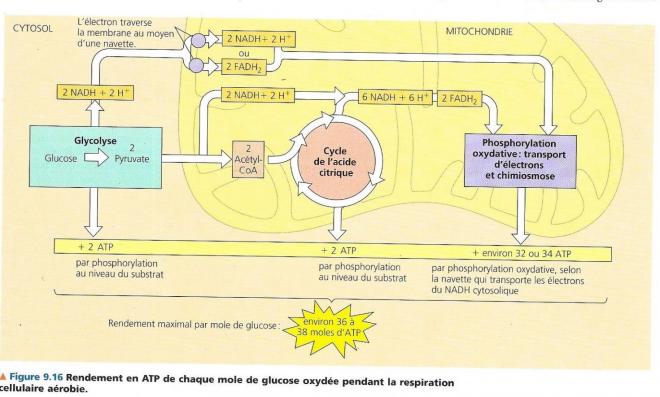
Techniquement, la respiration cellulaire aérobie ne comprend que les processus qui requièrent du dioxygène : Le cycle de l’acide citrique et la phosphorylation oxydative, nous y avons inclus la glycolyse qui n’utilise pas le dioxygène parce que la plupart des cellules aérobies qui tirent leur énergie du glucose font appel à ce processus pour obtenir le matériel de départ nécessaire à l’amorce du cycle de l’acide citrique. Les deux premières étapes, glycolyse et cycle de l’acide citrique sont les étapes cataboliques car elles dégradent le glucose et d’autres molécules organiques. La glycolyse qui a lieu dans le cytosol marque le début de la dégradation du glucose : elle scinde une môle de celui-ci en deux môles d’un composé appelé pyruvate. Le cycle de l’acide citrique qui se déroule dans la matrice mitochondriale termine la dégradation du glucose en oxydant un dérivé du pyruvate en dioxyde de carbone. Le dioxyde de carbone produit par la respiration représente donc des fragments de molécules organiques oxydées.
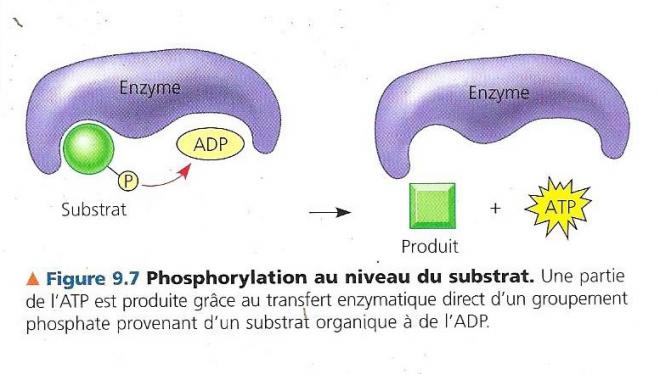
Quelques-unes des étapes de la glycolyse et du cycle de l’acide citrique sont des réactions d’oxydoréduction dans lesquelles les déshydrogénases transfèrent des électrons du substrat au NAD+, en formant du NADH + H+ . La phosphorylation oxydative et la chaîne de transport d’électrons qui représente le troisième stade de la respiration cellulaire aérobie accepte les électrons provenant des produits des deux premiers stades (le plus souvent par l’entremise de NADH + H+) et elle les transmet d’une molécule à l’autre. A la fin de la chaîne, les électrons se combinent à du proton (H+) et à du dioxygène pour former de l’eau. L’énergie libérée à chaque stade de la chaîne est emmagasinée sous une forme que la mitochondrie peut utiliser pour former de l’ATP. Ce mode de synthèse de l’ATP s’appelle phosphorylation oxydative, car il est alimenté par les réactions d’oxydoréduction de la chaîne de transport d’électrons ; le mot phosphorylation fait référence au fait que le phosphate transféré peut également s’appeler phosphoryle. Chimiosmose et le transfert d’électrons qui constituent ensemble la phosphorylation oxydative ont lieu dans la membrane interne de la mitochondrie. Près de 90 % de l’ATP générés par la respiration cellulaire provient de la phosphorylation oxydative. Une quantité moindre se forme directement au cours de certaines étapes (réactions) de la glycolyse et du cycle de l’acide citrique et ce grâce à un mécanisme appelé phosphorylation au niveau du substrat.
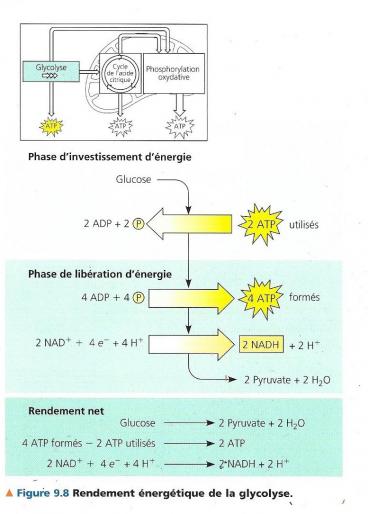
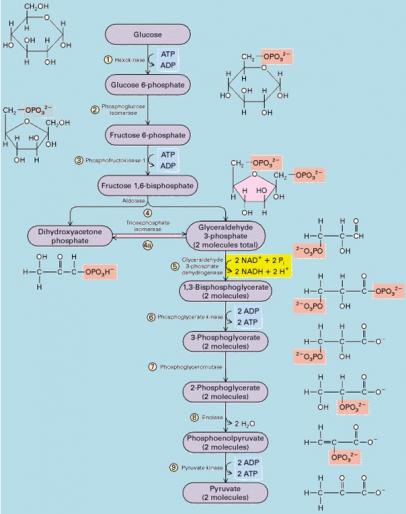
Dans ce mode de synthèse de l’ATP, une enzyme transfère un groupement phosphate d’un substrat à l’ATP au lieu d’ajouter un phosphate inorganique (le substrat fait ici référence à une molécule organique produite pendant le catabolisme du glucose). On estime que pour chaque mole de glucose dégradée en dioxyde de carbone et en eau au cours de la respiration cellulaire aérobie, la cellule produit environ 36 à 38 moles d’ATP, chacune contenant 30,5kJ/mol d’énergie libre. La respiration transforme l’énergie totale de la molécule de glucose en petites portions d’énergie plus utilisables. Voyons maintenant plus en détail les trois stades qui constituent la respiration cellulaire aérobie. C.LA GLYCOLYSE / libération d’énergie chimique en oxydant le glucose en pyruvate La glycolyse, c’est la dégradation du glucose. En premier lieu, le glucose (monosaccharide en C6) est scindé en deux monosaccharides en C3 Ensuite, ces petits monosaccharides sont oxydés pour donner deux molécules de pyruvate. (acide organique en C3 ionisé) La glycolyse se déroule en 10 étapes toutes connues et catalysées chacune par une enzyme spécifique. La première phase de la glycolyse est une phase d’investissement d’énergie, elle consomme de l’ATP (la cellule doit dépenser de l’ATP). La seconde phase est un retour sur investissement durant la phase de libération d’énergie (dividendes).
Le rendement net de la glycolyse est de deux môles d’ATP et de deux môles de NADH + H+ par mole de glucose
Le glucose entre dans la cellule grâce à une perméase, par diffusion facilitée puis il est phosphorylé par une enzyme Kinase, l’hexokinase. Toute kinase a pour fonction d’adjoindre un groupement phosphate provenant de l’ATP à une molécule organique, ici le glucose. Toutes les kinases nécessitent Mg2+ comme cofacteur pour fonctionner. La charge électrique négative conférée par le groupement phosphate empêche le glucose de sortir de la cellule.
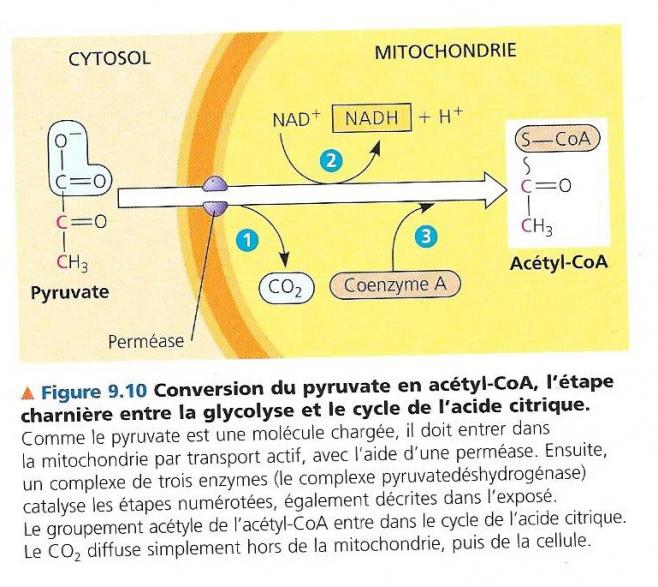
Groupement phosphate lié à un radical organique E LE CYCLE DE L’ACIDE CITRIQUE/achève l’oxydation des molécules organiques génératrice d’énergie. La glycolyse libère moins d’un quart de l’énergie contenue dans le glucose ; le reste est stocké dans les deux molécules de pyruvate. Le pyruvate entre dans la mitochondrie grâce à un mécanisme de cotransport de protons et de pyruvate. Les enzymes du cycle de l’acide citrique, synthétisées dans la mitochondrie terminent l’oxydation des deux môles de pyruvate. L’acide pyruvique est un α-cétoacide de formule CH3-CO-COOH, qui porte une fonction acide carboxylique (COOH) et une fonction cétone. Le pyruvate est la forme ionisée CH3-CO-COO- de cet acide. Après son entrée dans la mitochondrie, le pyruvate est d’abord converti en un composé appelé acétyl-CoA.
Cette étape charnière entre la glycolyse et le cycle de l’acide citrique est catalysée par un complexe multienzymatique qui active trois réactions :
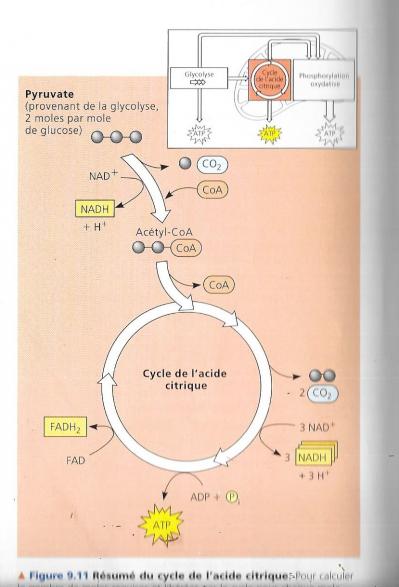
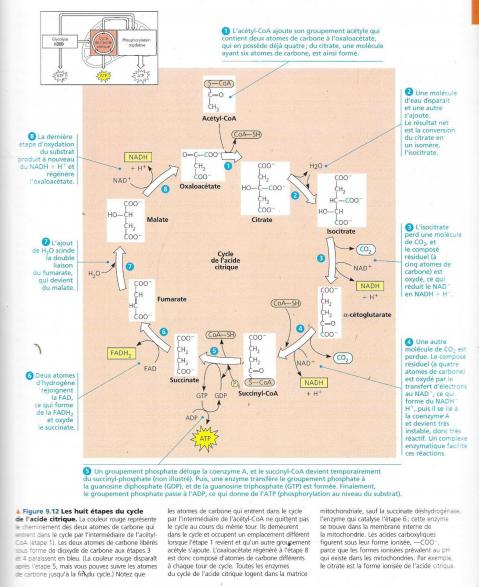
Cycle de l’acide citrique = cycle des acides tricarboxyliques car plusieurs des acides formés dans ce cycle possèdent trois groupements carboxyliques. On l’appelle également cycle de Krebs du nom du biochimiste allemand prix Nobel qui l’a découvert dans les années 1930. Il s’agit d’une véritable « fournaise métabolique » qui oxyde les carburants organiques issus du pyruvate.
La figure précédente qui montre un sommaire des entrées et des sorties pour ce cycle quand le pyruvate est dégradé en trois molécules de CO2 dont celle qui est libérée lors de la conversion du pyruvate en AcétylCoA. La phosphorylation au niveau du substrat produit une molécule d’ATP par cycle, mais une grande partie est transférée au NAD+, et à la coenzyme FAD durant les réactions d’oxydoréduction. Une fois réduites, les coenzyme NADH et FADH2 apportent leur chargement d’électrons très riches en énergie vers la chaîne de transfert d’électrons. Le cycle de l’acide citrique comporte huit étapes toutes catalysées par une enzyme spécifique. Le schéma suivant nous montre qu’à chaque tour de cycle deux atomes de carbone (en rouge) entrent sous forme de l’acétate (étape1) et deux autres atomes de carbone sortent (en bleu) sous la forme totalement oxydée du dioxyde de carbone (étapes 3 et 4). L’acétate entre dans le cycle après qu’une enzyme le lie à l’oxaloacétate, ce qui donne le citrate (étape1), le citrate étant la forme oxydée de l’acide citrique. Durant les sept étapes subséquentes la dégradation du citrate, et de nouveau, de l’oxaloacétate est formé. C’est la régénération de l’oxaloacétate qui fait que ce processus constitue un cycle. Pour chaque groupement acétyle qui entre dans le cycle, 3 moles de NAD+ sont réduites en NADH + H+ (étape 3,4 et 8). Au cours de l’étape 6, les électrons ne sont pas transférés au NAD, mais à un autre accepteur d’électrons, la Flavine adénine dinucléotide (ou FAD, molécule également issue du complexe de la vitamine B). A l’instar de la glycolyse, le cycle de l’acide citrique comprend une étape, étape 5, qui forme directement une môle de GTP par phosphorylation au niveau du substrat. Ce GTP est utilisé pour former de l’ATP, le seul ATP formé directement par le cycle de l’acide citrique. La majeure partie de l’ATP fournie par la respiration résulte de la phosphorylation oxydative, lorsque le NADH + H+ et le FADH2 engendrés par le cycle de l’acide citrique transmettent les électrons extraits des nutriments à la chaîne de transport des électrons. Ce faisant, ils fournissent l’énergie nécessaire à la formation de l’ATP à partir de l’ADP.
Jusqu’à présent nous avons abordé deux étapes de la respiration cellulaire à savoir la glycolyse et le cycle des acides tricarboxyliques, ces deux étapes ne livrent que deux moles d’ATP par mole de glucose au moyen de la phosphorylation au niveau du substrat. Il en ressort que ce sont le NADH + H+ et le FADH2 de libérer la majeure partie de l’énergie extraite des nutriments. Ces transporteurs d’électrons relient la glycolyse et l’acide citrique, à la machinerie de la phosphorylation oxydative qui alimente la synthèse de l’ATP avec l’énergie libérée par la chaîne de transport des électrons.
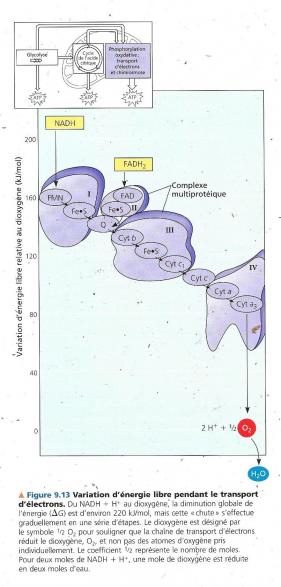
La figure qui suit montre la succession de transporteurs d’électrons dans la chaîne et la baisse d’énergie libre qui accompagne le transfert des électrons. Durant le transport d’électrons dans cette chaîne, les transporteurs d’électrons oscillent entre l’état réduit et l’état oxydé Chaque élément de la chaîne devient réduit lorsqu’il accepte des électrons de son voisin d’amont (qui a moins d’affinité pour les électrons), puis il retrouve sa forme oxydée en cédant des électrons à son voisin d’aval (qui a plus d’affinité pour les électrons). Différentes techniques (rayons ultra-violets, poisons) ont permis de déterminer l’ordre dans lequel les transporteurs se succèdent.
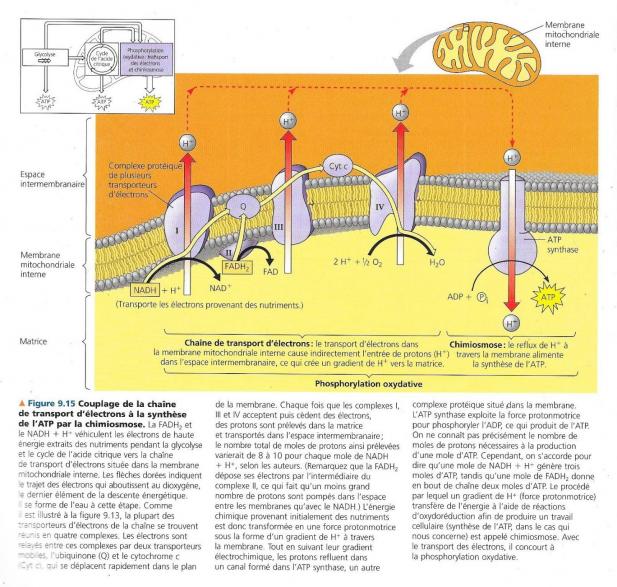
Les électrons extraits des nutriments par le NAD+ au cours de la glycolyse et du cycle de l’acide citrique sont transférés par le NADH + H+ à la première molécule de la chaîne qui est une flavoprotéine contenant un groupement prosthétique appelé flavine mononucléotide (désigné par FMN dans le complexe I), les électrons sont ensuite transférés à une protéine contenant du fer et du souffre fortement liés (Fe.S dans le complexe I). A partir de là, les électrons sont transférés à un liquide appelé ubiquinone ( Q dans la figure ) , c’est une petite molécule avec une queue hydrophobe et donc mobile dans la membrane. La plupart des transporteurs entre Q et le dioxygène sont des cytochromes : Ils possèdent un groupement prosthétique appelé groupement hème, il possède un atome de fer qui accepte et cède des électrons (Fe2+/Fe3+), tous les cytochromes de la chaîne de transport d’électrons ne sont que légèrement différents et comportent un groupement hème. Le dernier cytochrome de la chaîne (a3 sur la figure) cède des électrons au dioxygène, très oxydant, c’est ainsi que le dioxygène en captant deux protons dans le milieu produit de l’eau. O2 + 2e- + 2 H+ à H2O Le NADH2 autre coenzyme réduite du cycle de l’acide citrique apporte aussi des électrons, mais le niveau d’énergie auquel il apporte ses électrons à la chaîne de transport (complexe II) de ceux-ci est plus faible que celui du NADH + H+. Le chaîne de transport, dans ce cas, fournit à la synthèse d’ATP 33% d’énergie en moins qu’avec NADH + H+. La fonction de la chaîne n’est pas de former de l’ATP directement, mais de libérer de l’énergie de manière contrôlée. C’est un mécanisme appelé chimiosmose qui couple cette libération d’énergie à la synthèse d’ATP.
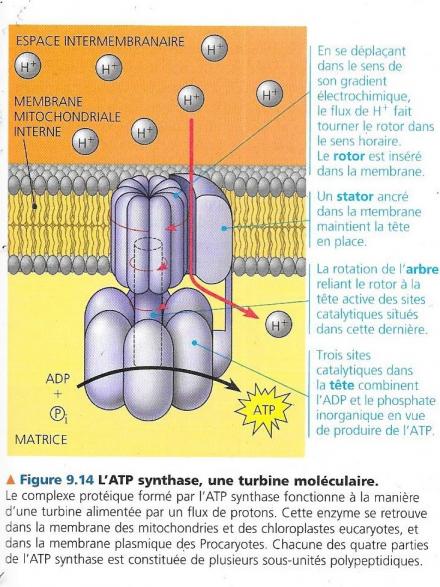
La membrane interne de la mitochondrie renferme de nombreux exemplaires d’un complexe protéique appelé ATP synthase, l’enzyme qui fabrique réellement l’ATP à partir de l’ADP et du phosphate inorganique. Celle-ci ressemble à une pompe ionique qui fonctionne à rebours, elle utilise l’énergie d’un gradient électrochimique existant pour synthétiser l’ATP. Il s’agit en fait d’un gradient de protons, correspondant à une différence de concentration en H+ de part et d’autre de la membrane ou différence de pH. C’est de là que provient la source d’énergie de l’ATP synthase. Ce mécanisme d’utilisation de l’énergie du gradient de proton s’appelle chimiosmose. Cela correspond à une pression chimiosmotique de poussée des de H+ à travers une membrane. L’ATP synthase est un complexe protéique composé de plusieurs sous-unités regroupées en quatre parties :
En traversant l’étroit passage entre le rotor et le stator dans le sens de leur gradient électrochimique, les protons déclenchent une rotation du rotor et de l’arbre (une centaine de révolutions par seconde), ce qui entraîne un changement de conformation de la tête immobile. Ce changement de conformation entraîne une activation de trois sous-unités formant la tête, ce qui amène l’ADP à se combiner avec le phosphate inorganique pour former l’ATP. Le gradient de protons est créé, car la chaîne est un convertisseur d’énergie qui utilise le flux exergonique d’électrons pour véhiculer les H+ au travers de la membrane, de la matrice vers l’espace intermembranaire. La différence de concentration en H+ ainsi générée pousse les protons à refluer vers la matrice et le seul endroit perméable aux H+ est l’ATP synthase qui produit une phosphorylation oxydative de l’ADP alimentée par le transfert exergonique des H+. a certains endroits de la chaîne transporteuse d’électrons, certaines composantes de celle-ci captent et libèrent les protons en même temps que les électrons. Donc à certaines étapes de la chaîne, les transporteurs captent les H+ et libèrent des H+ dans la solution environnante. Les transporteurs d’électrons sont disposés dans la membrane en manière telle que les H+ soient prélevés dans la matrice mitochondriale puis déposés dans l’espace inter-membranaire. Le gradient électrochimique ainsi formé se nomme force protonmotrice.
Dans les mitochondries, ce sont les réactions chimiques exergoniques qui permettent l’entrée des électrons dans la chaîne de transport et la constitution du gradient de protons. La synthèse d’ATP représente le travail effectué. Dans les chloroplastes, ce même mécanisme existe pour produire de l’ATP pendant la photosynthèse. Cependant, dans ce cas, c’est l’énergie lumineuse qui permet l’entrée des électrons dans la chaîne et la formation du gradient de H+
Pendant la respiration aérobie, la majeure partie de l’énergie suit la séquence suivante (sauf pour l’ATP produit par la glycolyse) : Glucose à NADH + H+ à chaîne de transport d’électrons à force protonmotriceà ATP Faisons le bilan du profit net réalisé en ATP chaque fois qu’une molécule de glucose est oxydée en 6 moles de CO2. Les 3 services de l’entreprise métabolique qu’est la respiration sont la glycolyse, le cycle de l’acide citrique et la phosphorylation oxydative. La figure qui suit montre un bilan du rendement en ATP par mole de glucose oxydée. Globalement, la glycolyse et le cycle de Krebs produisent 4 moles d’ATP auxquelles il faut ajouter le nombre de moles d’ATP produites par phosphorylation oxydative par apport de NADH + H+ ou FADH2. Chaque mole de NADH + H+, en apportant ses électrons à la chaîne de transport contribue assez à la force protonmotrice pour produire au maximum 3 môles d’ATP.
Grossièrement, l’on considère donc qu’une mole de NADH + H+ génère une force protonmotrice suffisante pour permettre la formation de 3 ATP, le FADH2 quant à lui peut permettre la formation de 2 ATP maximum. Tenant compte de ce fait et du nombre de FADH2 et de NADH + H+ produits par la glycolyse et le cycle de l’acide citrique, on déduit que la production maximum d’ATP par mole de glucose se situe au maximum entre 36 et 38 moles d’ATP. Voyons pourquoi nous ne pouvons donner de nombre exact. A. La phosphorylation oxydative et les réactions d’oxydo-réduction ne sont pas directement couplées : le rapport entre le nombre de moles de NADH et le nombre de moles d’ATP n’est pas un nombre entier. L’entrée de 1 NADH permet d’extraire 10 H+ vers l’espace intermembranaire et la synthèse d’un ATP par l’ATP nécessite le retour de 3 à 4 H+ dans la matrice mitochondriale. Un NADH peut donc produire indirectement 2,5 à 3,3 ATP, généralement on arrondit et on considère que 1 NADH peut générer environ 3 ATP. Le FADH2 (1 môle) lui, intervenant à un niveau plus bas d’énergie dans la chaîne transporteuse ne peut générer que 1,5 à 2 moles ATP. B. L’entrée des électrons dans la mitochondrie, via le NADH se fait par divers types de navettes qui n’ont pas toutes la même efficacité car selon le type de navette, les 2 électrons provenant de la glycolyse sont transférés au NAD+ ou au FAD, en fonction de cette différence, les deux électrons transférés permettront la formation de 2 ou de 3 ATP au maximum. C. Une partie de la force protonmotrice peut également être utilisée pour d’autres fonctions comme l’entrée de pyruvate dans la mitochondrie et réduire le rendement de production d’ATP. Si ce n’est pas le cas et que l’ensemble de la force protonmotrice sert à générer de l’ATP, cette production serait de 34 moles d’ATP par phosphorylation oxydative et 4 moles d’ATP provenant de la phosphorylation au niveau du substrat lors de la glycolyse et le cycle de l’acide citrique. Ce qui fait au total 38 ATP, à moins que la navette utilisée pour introduire les électrons de NADH issu de la glycolyse soit la navette à FAD, dans ce cas c’est 36 ATP qui seraient produits. En effet : Si la navette à FAD n’est pas utilisée, il y a 10 NADH + 10 H+ qui arrivent à la phosphorylation oxydative et 2 FADH2 : à 10 . 3 + 2 . 2 = 34 ATP produits par la phosphorylation oxydative. A cela il faut ajouter les 4 ATP issus de la glycolyse et du cycle de l’acide citrique, cela fait au total 38 ATP par mole de Glucose. Si la navette à FAD est utilisée, on obtient 36 ATP Il est maintenant possible d’évaluer l’efficacité de la respiration cellulaire aérobie, c’est-à-dire le pourcentage d’énergie chimique enfermée dans le glucose qui est utilisé pour former de l’ATP La combustion complète du glucose libère 2870 kJ par mole de glucose (ΔG = -2870 kJ/mol). En conditions chimiques cellulaires, la phosphorylation de l’ADP emprisonne environ 30,5 kJ/mol dans les liaisons d’une mole d’ATP. La respiration cellulaire aérobie cellulaire produisant au maximum 38 moles d’ATP par mole de glucose, l’équivalent en énergie est de 38 x 30,5 kJ = 1159 kJ. Le rendement sera de 1159/2870, c’est-à-dire 0,4 soit 40 % de l’énergie stockée dans le glucose qui est transférée et stockée dans l’ATP. On croit cependant que la valeur dans les conditions cellulaire est encore plus élevée. Le reste de l’énergie du glucose se perd sous forme de chaleur pouvant servir à maintenir notre température corporelle ou encore à évaporer l’eau lorsque nous transpirons. Le processus de respiration aérobie est un processus de conversion d’énergie fort efficace, notamment à une voiture moderne qui convertit seulement 25 % de l’énergie contenue dans l’essence en mouvement.
En l’absence de dioxygène, très électronégatif qui attire les électrons dans la chaîne de transport, celle-ci s’arrête. Certains organismes s’en passent et produise de l’ATP par respiration cellulaire anaérobie. Dans ce cas il existe une chaîne de transport d’électron, mais l’accepteur final n’est pas de dioxygène. (NO3-, SO42-, CO2, Fe3+ …)
D’autres organismes sont capables de produire de l’ATP sans chaîne de transport d’électrons et sans dioxygène, par un processus chimique que l’on appelle la fermentation. Dans l’oxydation des nutriments qui signifie perte d’électrons, l’oxydant (accepteur d’électrons) ne doit pas toujours être l’oxygène, d’autres accepteurs peuvent convenir. Ainsi dans l’oxydation des nutriments au cours de la glycolyse, c’est le NAD+ qui joue ce rôle. En fait, certains microorganismes ont besoins d’un milieu sans oxygène pour survivre, l’oxygène étant parfois pour eux un poison produisant des oxydations nocives. Remarquons déjà que même en conditions sans oxygène, la glycolyse fonctionne toujours puisque l’accepteur d’électrons est le NAD+. Le catabolisme anaérobie, peut en plus emprunter la voie de la fermentation qui est une continuation de la glycolyse à partir du pyruvate. La glycolyse peut continuer à fonctionner tant qu’il y a du NAD+ disponible, mais au fur et à mesure que NADH + H+ se forme, le stock de NAD+ à tendance à diminuer et pourrait devenir insuffisant s’il n’était pas régénéré à partir du NADH. Dans la respiration aérobie, le NADH produit est consommé dans la phosphorylation oxydative. En anaérobie, c’est la fermentation qui consomme le NADH et régénère ainsi le NAD+ Il y a deux voies possibles de fermentation :
Fermentation lactique : dans ce cas, le pyruvate se fait directement réduire par le NADH+ H+ et donne ainsi du lactate (forme ionisée de l’acide lactique) sans que du dioxyde de carbone soit libéré. Cette production d’acide lactique sert dans l’industrie laitière à fabriquer des fromages et du yogourt. D’autres sous-produits d’autres types de fermentation peuvent produire par exemple de l’acétone ou du méthanol utilisés dans l’industrie à d’autres fins.
Remarquons que le dernier sous-produit de la fermentation est une molécule qui contient encore de l’énergie mais que la cellule ne peut plus oxyder (éthanol, lactate).
Que ce soient respiration cellulaire ou fermentation, elles font toutes deux appel à la glycolyse qui fournit de manière nette deux moles de ATP 2 ATP également par le cycle de l’acide citrique) par mole de glucose (phosphorylation au niveau du substrat). Au niveau de la fermentation comme au niveau de la respiration cellulaire aérobie ou anaérobie, c’est le NAD+ qui est l’agent oxydant qui accepte les électrons dérivés de la transformation des nutriments au cours de la glycolyse. Dans la respiration cellulaire (aérobie ou anaérobie) et la fermentation, les deux mécanismes d’oxydation de NADH +H+ en NAD+ sont différents. Ce mécanisme est indispensable pour régénérer le NAD+ nécessaire à la glycolyse. Dans la fermentation, le dernier accepteur d’électrons est une molécule organique (le pyruvate pour la fermentation lactique et l’acétaldéhyde dans la fermentation alcoolique). Dans la respiration cellulaire, le dernier accepteur est le dioxygène pour la respiration aérobie et NO3-, SO42-, CO2 ou Fe3+ dans la respiration anaérobie. Pendant la respiration cellulaire , non seulement du NADH est régénéré, mais aussi une importante quantité d’ATP par la phosphorylation oxydative lorsque les électrons du NADH + H+ issu de l’étape 6 de la glycolyse sont déversés dans la chaîne respiratoire aérobie. L’oxydation du pyruvate dans la respiration cellulaire par l’intermédiaire du cycle de l’acide citrique engendre d’avantage d’ATP que la fermentation. L’ensemble de la respiration cellulaire génère 36 à 38 moles d’ATP par mole de glucose et la fermentation n’en génère que 2 l’énergie du pyruvate ne pouvant que très peu être exploitée. La respiration a un rendement de 40 % de l’énergie du glucose, ce qui nous donne l’évaluation de 2% de l’énergie du glucose extraite de celui-ci par la fermentation.
La glycolyse est commune à la fermentation et à la respiration cellulaire, similitude expliquée par l’évolution. En effet, bien avant que le dioxygène apparaisse dans l’atmosphère terrestre, c’est la glycolyse qui servait à fournir de l’ATP. Les fossiles de bactéries les plus anciens datent de 3,5 milliards d’années, mais le dioxygène n’est apparu en quantité notable dans l’atmosphère qu’il y a 2,7 milliards d’années en conséquence de l’activité photosynthétique des cyanobactéries. Les premiers procaryotes produisaient sans doute leur ATP uniquement à partir de la fermentation. D’autre part, la glycolyse est une voie métabolique extrêmement répandue, ce qui suggère qu’elle est apparue très tôt dans l’histoire de la vie. Remarquons également qu’elle se déroule dans le cytoplasme sans avoir besoin d’organites cellulaire des eucaryotes apparus un milliard d’années après les procaryotes. La glycolyse existe encore chez tous les organismes modernes, aussi bien dans la fermentation que comme stade précurseur de la respiration cellulaire.
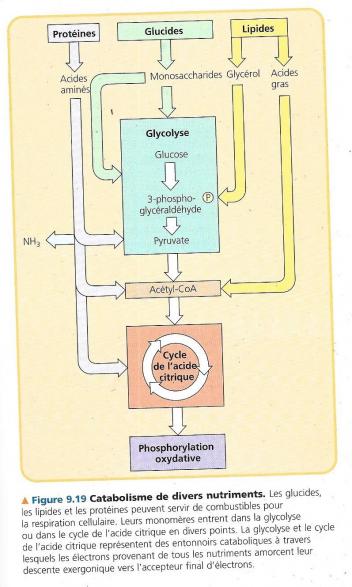
La glycolyse et le cycle de l’acide citriques sont au carrefour de plusieurs voies cataboliques et anaboliques. Jusqu’à présent, nous n’avons considéré que le glucose comme combustible de la fermentation et de la respiration cellulaire. Pourtant le glucose ne constitue pas la majorité des nutriments ingérés par exemple par l’homme. La respiration cellulaire peut produire de l’ATP à partir d’autres nutriments comme :
Dans le système digestif :
Les molécules organiques que nous ingérons ne sont pas toutes destinées à la production d’ATP. En effet, doit fournir aux cellules les chaînes carbonées nécessaires à la formation de molécules structurales. Certaines monomères organiques issues de la digestion peuvent être utilisés directement. C’est le cas des acides aminés provenant des protéines alimentaires sont hydrolysées en acides aminés qui serviront à la synthèse de protéines de l’organisme. Il arrive cependant que l’organisme ait besoin de molécules précises que l’alimentation ne fournit pas. Les produits intermédiaires de la glycolyse et du cycle de l’acide citrique peuvent être détournés du catabolisme et entrer dans l’anabolisme en servant de précurseurs à la synthèse des molécules nécessaires :
Ces voies anaboliques de biosynthèse ne produisent pas d’ATP, elles en consomment. La glycolyse et le cycle de l’acide citrique permettent à nos cellules de convertir certaines molécules selon leurs besoins et les circonstances. Par exemple le dihydroxyacétone phosphate de la glycolyse peut être converti en un des principaux précurseurs de lipides. Si notre apport alimentaire est trop important, même sans apport de lipides, nous engraissons. Tout ce métabolisme ne peut fonctionner que si les enzymes nécessaires aux réactions sont présentes.
L’économie métabolique respecte la loi de l’offre et de la demande. La cellule ne gaspille pas d’énergie à produire plus d’une substance qu’il ne lui en faut. Lors d’un surplus en un acide aminé particulier, la voie anabolique qui le synthétise à partir d’un intermédiaire du cycle de l’acide citrique se ferme. Il s’agit d’un mécanisme de rétro-inhibition : le produit final de la voie anabolique inhibe l’enzyme qui catalyse une des premières étapes de cette voie. L’organisme évite ainsi de mobiliser des produits intermédiaires à des usages non essentiels.
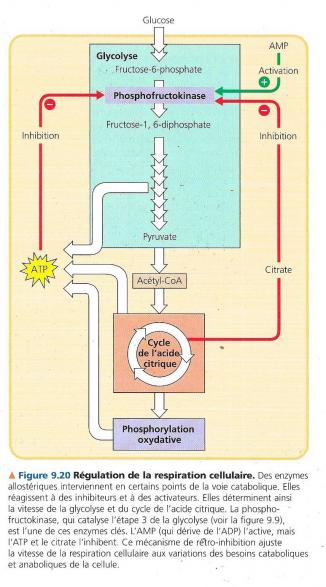
La cellule gère aussi son catabolisme. Si elle travaille dur et que sa concentration en ATP commence à diminuer, la respiration cellulaire s’accélère. Quand il y a amplement d’ATP pour satisfaire la demande, la respiration cellulaire ralentit. Cela permet à la cellule d’économiser des précieuses molécules organiques en vue d’autres fonctions. Ici encore, la régulation intervient sur l’activité enzymatique en des points stratégiques de la voie catabolique. L’une de celles-ci est la phosphofructokinase, qui catalyse l’étape 3 de la glycolyse. C’est la première étape durant laquelle un substrat est irréversiblement dirigé vers la voie glycolytique. En contrôlant le débit de cette étape, la cellule peut accélérer ou ralentir le processus catabolique entier ; par conséquent, la phosphofructokinase détermine la vitesse de la respiration cellulaire. La phosphofructokinase est une enzyme allostérique qui possède en plus de son site actif, des sites récepteurs destinés à des inhibiteurs et des activateurs spécifiques. L’ATP l’inhibe et l’AMP (adénosine monophosphate, un dérivé de l’ATP). Donc lorsque l’ATP s’accumule, l’inhibition de la phosphofructokinase ralentit la glycolyse. Inversement lorsque la cellule consomme d’avantage d’ATP qu’elle n’en produit, l’enzyme est réactivée. En outre la phosphofructokinase est sensible au citrate, le premier produit du cycle de l’acide citrique. S’il augmente beaucoup dans les mitochondries une certaine quantité passe dans le cytosol à l’aide d’une perméase et inhibe la phosphofructokinase. Au fur et à mesure que le citrate s’accumule , la glycolyse ralentit et l’apport d’acétate au cycle de l’acide citrique. Si au contraire la consommation de citrate augmente à la suite d’une augmentation de la de la demande d’ATP ou à cause de l’utilisation de produit intermédiaire du cycle de l’acide citrique à des fins anaboliques, la glycolyse s’accélère et s’adapte à la demande. D’autres enzymes interviennent également en d’autres points de la glycolyse et du cycle de l’acide citrique. Elles sont contrôlées par des mécanismes qui favorisent l’équilibre métabolique. Le métabolisme cellulaire est un processus économique efficace et souple. La respiration cellulaire est à mettre dans le contexte des processus énergétiques et chimiques des écosystèmes. La respiration cellulaire libère de l’énergie nécessaire à la vie et ne la produit pas nos cellules extraient l’énergie que la photosynthèse a préalablement stockée dans notre nourriture.
|



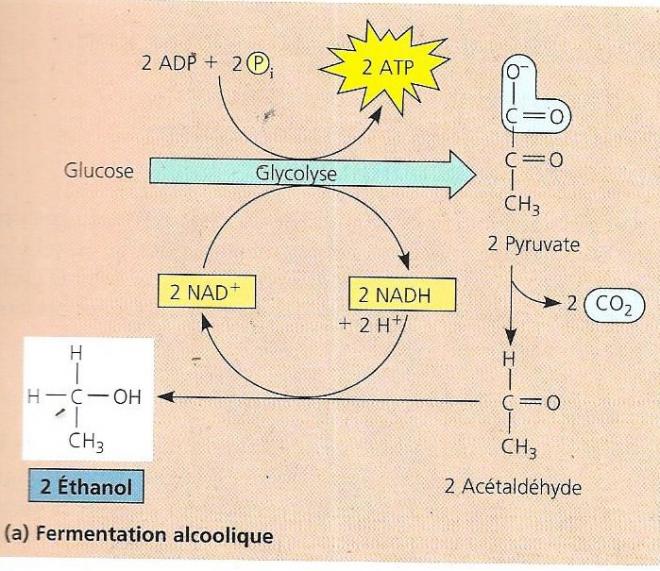
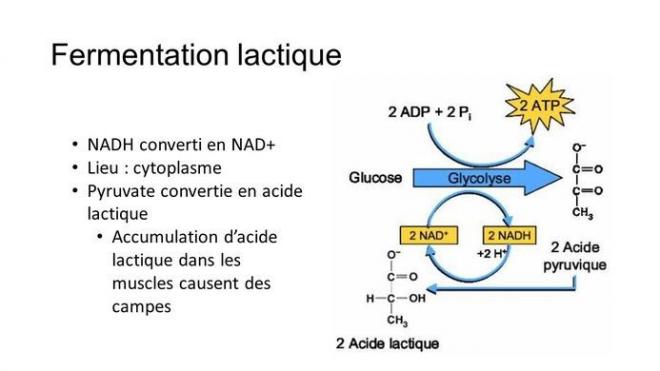 Le pyruvate est donc au carrefour de deux voies cataboliques, soit le pyruvate entre dans la mitochondrie pour donner de l’acétyl-coA qui entre dans le cycle de l’acide citrique, soit il emprunte une voie de fermentation.
Le pyruvate est donc au carrefour de deux voies cataboliques, soit le pyruvate entre dans la mitochondrie pour donner de l’acétyl-coA qui entre dans le cycle de l’acide citrique, soit il emprunte une voie de fermentation.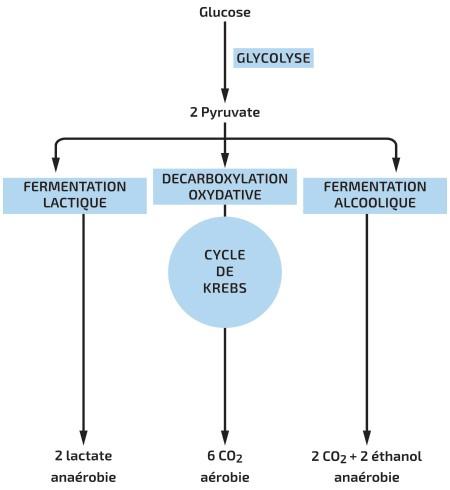
Date de dernière mise à jour : 24/12/2018
Ajouter un commentaire


