

Classification / cladistique
LA SYSTEMATIQUE AUJOURD'HUI
1. Définitions préalables
1.1 L'espèce??
Une espèce est un ensemble monophylétique d'individus se reconnaissant comme partenaires sexuels. Leur descendance est féconde
1.2 Le caractère et l'homologie
?Un caractère est un attribut observable d'un organisme.
Pour aboutir à un arbre, il faut que les caractères soient comparables et comparés. Lorsque deux structures sont similaires mais pas exactement identiques , on peut distinguer deux états - au minimum - du même caractère. En d'autres termes , le caractère est la désignation de ce que l'on observe ( couleurs des yeux ou site numéro 177 de la séquence du gène de l'hémoglobine bêta) et l'état est la formulation de ce que l'on trouve de discriminant au sein de l'échantillon d'organisme ( marron, bleu ou G,T). Caractère et état de caractère différent dans leur désignation mais pas dans leur nature. Dans une optique comparative , un caractère est donc une structure identifiée comme similaire dans deux ou plusieurs organismes. Similaire signifie que :
1) la structure peut être exactement identique , ou différer par différents degrés au sein des organismes observés .
2) cette structure puisqu'analysée phylogénétiquement , constitue une hypothèse d'homologie ( on dit encore homologie primaire ) .
On constate d'emblée que dans cette approche comparative , la notion de caractère est indissociable de la notion de similarité , et donc d'homologie . La définition complète du caractère en systématique est donc une série d'attributs observables d'organismes sur lesquels on peut poser une hypothèse d'homologie.
Dans un nombre de cours de sciences naturelles, on donne deux définitions de l'homologie sans les replacer l'une par rapport à l'autre d'un point de vue opérationnel.
Premièrement , on parle de structures homologues lorsqu'elles sont héritées d'un ancêtre commun. C'est l'homologie de descendance, dite homologie secondaire. Enoncée comme cela, cette notion n'est pas intéressant opérationnellement puisque ;
1) elle ne dit pas comment on fait pour savoir qu'elle est héritée d'un ancêtre commun.
2) elle ne spécifie pas le cadre taxonomique dans lequel ces structures sont observées. En effet l'homologie de descendance ne prend tout son sens que dans un cadre taxonomique donné. Sans un cadre taxonomique de référence , dans la mesure où l'arbre de vie est unique , tout serait finalement homologue à tout. Par exemple l'aile du Perroquet est homologue à l'aile de l'Aigle en tant qu'aile ( homologie au niveau taxinomique des oiseaux ) et aussi en tant que membre antérieur ( niveau taxinomique des tétrapodes ). Tandis que l'aile du perroquet et l'aile de la Chauve-souris, si elles restent bien homologues au niveau du taxon tétrapodes en tant que membres pairs antérieurs ne sont plus homologues en tant qu'ailes ( à ce même niveau ).
Deuxièmement, on parle aussi de structures homologues lorsque, comparées dans plusieurs organismes lde même plan d'organisation, ces structures entretiennent les mêmes connexions avec les structures voisines et ce quelles que soient leurs formes et leurs fonctions. Il s'agit de là de la définition que Richard Owen ( 1804-1892 ) a formulée en 1843 à partir du principe des connexions d'Etienne Geoffroy Saint Hilaire ( 1772-1844 ). En fait il faut replacer ces deux définitions dans le temps. On formule d'abord une hypothèse d'homologie ( seconde définition ), laquelle est ensuite - éventuellement - confirmée par l'arbre qui sera trouvé ( première définition ). La définition d'Owen correspond à une hypothèse initiale d'homologie dite homologie primaire. C'est ce que nous faisons lorsque nous déclarons initialement que l'os radius du Dauphin peut être comparé ( ou est similaire ) à l'os radius de l'Opossum ou au radius d'une chauve-souris.
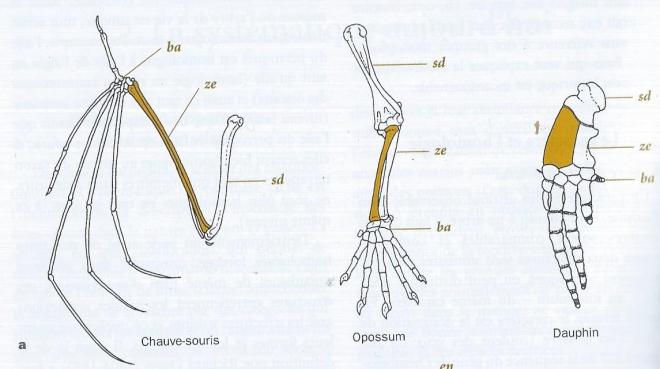 fig 1 a. Membre antérieur de trois Mammifères illustrant la notion de plan d'organisation : le radius est coloré ; ba = basipode ; sd = stylopode ; ze = zeugopode.
fig 1 a. Membre antérieur de trois Mammifères illustrant la notion de plan d'organisation : le radius est coloré ; ba = basipode ; sd = stylopode ; ze = zeugopode.
Puisque lorsque nous codons leurs différences en autant d'états sous la définition d'un même caractère "radius". Aucun de ces radius n'a la même forme . S'ils sont perçus comme homologues , c'est que ces os s'inscrivent chacun dans un plan d'organisation constant du membre chiridien , entre le stylopode proximal ( humérus ), et le basipode distal ( os du carpe). Le radius est l'un des éléments du segment intermédiaire le zeugopode. C'est aussi ce que nous faisons lorsque nous alignons des séquences d'acides nucléiques ou d'acides aminés. Lorsques les séquences similaires sont placées les unes au dessous des autres , nous mettons en place un plan d'organisation . Sur une colonne ( un site ) , les nucléotides ou les acides aminés sont supposés être homologues, car leurs positions dans la séquence sont les mêmes. C'est bien le même principe des connexions , appliqués aux molécules. Pour formuler une hypothèse d'homologie , on utilise donc un plan, sorte de grille de lecture abstraite des rapports entre les structures. Mais on peut également utiliser des données de l'ontogenèse ( développement de l'organisme ) montrer que les structures en présence dérivent l'une de l'autre. Par exemple, on sait par ce moyen notamment par l'embryologie des Marsupiaux, que deux des trois os de l'oreille moyenne des Mammifères ( marteau et enclume ) sont homologues à certains os impliqués dans l'articulation de la mâchoire au crâne des autres vertébrés ( articulaire et carré respectivement).
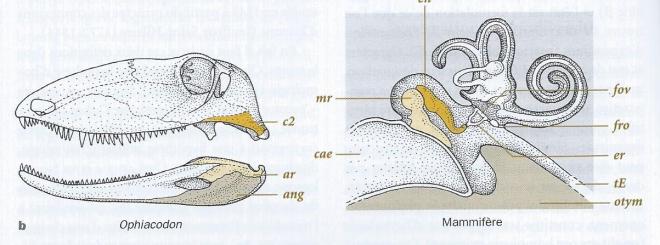
 fig 2
fig 2
Cette articulation a été conservée chez les Mammifères malgré un transfert dans la sphère auditive . Ce transfert documenté du point de vue de l'ontogenèse , est également très bien documenté par le registre fossile permo-triassique.
?L'homologie de descendance quant à elle, ne peut être obtenue qu'à travers un arbre . Elle correspond en fait au résultat de l'analyse. En effet, la méthode cladistique choisit l'arbre le plus parcimonieux, celui dont les données de départ coûtent le plus petit nombre d'évènements évolutifs le long de ses branches. Une fois l'arbre choisi, les positions dans l'arbre des changements de chaque caractère sont déterminés. On constate alors qu'un état de caractère est distribué de manière groupée dans l'arbre , de telle sorte que son apparition n'a lieu que sur une seule branche . Il s'agit alors d'une structure homologue car héritée d'un seul ancêtre commun hypothétique, chez qui cette structure est apparue pour la première fois. Cette structure homologue est la synapomorphie ou homologie secondaire. En revanche, une autre structure d'un autre caractère apparaît à différents endroits de l'arbre . Elle est homoplasique ; il s'agit d'une ressemblance qui n'est pas héritée d'un ancêtre commun. Elle révèle que l'hypothèse d'homologie initiale était fausse .
Une synapomùorphie est donc un caractère dérivé ou apomorphie partagé par deux ou plusieurs taxons et définissant ainsi un groupe monophylétique.La synapomorphie est une notion plus récente que celle d'homologie, qui a émergé à la suite des travaux récents des phylogénéticiens. En effet des organes anatomiques homologues sont certes l'expression d'une synapomorphie, mais la synapomorphie ne concerne pas obligatoirement des organes différenciés dont il faut détecter l'homologie : elle peut porter sur des organes parfaitement identiques, voire sur des caractères qui ne sont pas anatomiques (par exemple biochimiques).
Pour conclure sur cette question on peut rappeler que le concept d'homologie se divise en homologie ( similitude héritée d'un ancêtre commun ) et homoplasie ( similitude non héritée d'un ancêtre commun : convergences, réversions)
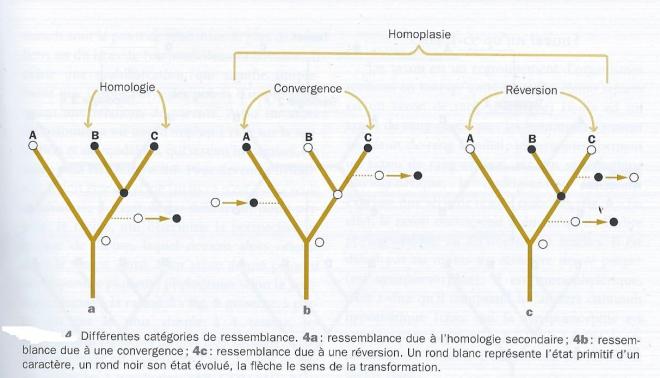 fig 3
fig 3
Extrait de : "Classification phylogénétique du vivant " de G. Lecointre et H. Le Guyadier chez Belin
?La découverte d'une similitude se traduit par une hypothèse d'homologie ou homologie primaire : on postule qu'il s'agit d'une similitude héritée d'un ancêtre commun. L'arbre le plus parciminieux donnera la réponse. L'homologie de descendance est dite homologie secondaire car elle constitue le résultat de l'analyse : toutes les homologie primaires confirmées sont des homologies secondaires ou synapomorphies. Ce sont des ressemblances héritées d'un ancêtre commun hypothétique.Toutes les homologies primaires réfutées sont des homoplasies , c'est à dire des ressemblances non héritées d'un ancêtre commun.
1.3 Les arbres en cladistique
La description des arbres utilise simultanément des termes de la théorie des graphes et des métaphores issues de la tradition évolutionniste. selon les premiers , un arbre est un graphe connexe non cyclique . Clui-ci comporte des liens ( ou arêtes, arcs, branches) qui relient entre eux des sommets. Le graphe est non cyclique parce qu'un seul chemin permet de relier deux sommets entre eux ( l'arbre n'est pas un réseau ). Les sommets internes sont des noeuds ( points de rencontre de trois liens ); les sommets externes sont des feuilles ( ou taxons terminaux, ou unités évolutives ). Les liens internes sont aussi appelés branches ( ou segments ) internodaux ; les liens externes , branches terminales . Un arbre dont chaque noeud est le point de rencontre entre trois liens est dit totalement résolu. Lorsque l'arbre est enraciné , on voit chaque branche interne donner deux branches filles . Cela correspond simplement au contenu en information de l'arbre,qui, dans ce cas est maximal: toutes les relations de parenté sont étayées. En revanche un arbre dont certains noeuds sont le point de rencontre de plus de trois liens est dit irrésolu ( ou partiellement irrésolu ) . Il existe une multifurcation qui exprime simplement que l'arbre porte des points d'imprécision quant aux relations de parenté . Avoir un arbre dichotomique ou non n'implique rien sur la spéciation et ses modalités , qui restent inconnues . Un arbre peut être non raciné . Pour devenir phylogénie il doit être enraciné ( mais ce n'est pas la seule condition ) : Il s'agit de lui définir un point d'origine , la racine , qui détermine la perspective de lecture de l'arbre, lequel devient alors orienté dans le temps. Ainsi à un arbre donné peuvent correspondre plusieurs phylogénies selon la position donnée à la racine. La figure qui suit présente à partir du cas le plus simple à quatre taxons, les trois topologies possibles ( arbres non racinés). Classer 4 taxons c'est les séparer 2 par 2 . C'est ce que montrent les trois topologies : (AB,CD);(AC,BD);(AD,BC).

fig 4
Dans ce type de présentation, les noeuds peuvent pivoter autour du segment médian. Cependant sur de telles représentations l'origine du temps ne peut être représentée . Indiquer la racine d'un arbre c'est indiquer l'origine du temps. Sur la figure ci-dessus, toutes les possibilitésde racines sont données pour chaque topologie . Comment choisir la racine ? Selon l'approche hennigienne classique , l'arbre se trouve obligatoirement sur l'extragroupe qui a été choisi pour polariser les caractères , c'est à dire pour déterminer le sens de de transformation de ses états. En analyse cladistique informatisée, on prend soin de coder les caractères d'un ou plusieurs extragroupes , sur lequel l'arbre résultant de l'analyse sera ultérieurement enraciné. Les logiciels de construction d'arbres trouvent la solution optimale sans avoir recours à une racine préétablie ; c'est l'utilisateur qui produit la racine en fin de course.
tiré de " Classification phylogénétique du vivant" par G.Lecointre et H. Le Guyadier chez Belin
Le principe dit du « groupe externe » ou « extra-groupe » est généralement utilisé pour enraciner un arbre phylogénétique, c'est-à-dire désigner le nœud correspondant à la classe racine (contenant toutes les autres classes). On estime que tout état de caractère observé en dehors du groupe interne est plésiomorphe pour le groupe interne. Au contraire, tout état propre à des taxons du groupe d'étude est considéré apomorphe.
Un caractère ancestral (homologie ancestrale ou plésiomorphie) chez une espèce au sein d'un taxon est un caractère qui n'a pas subi de modification au cours de l'évolution.
Ce caractère homologue n'a donc pas été conservé depuis le premier ancêtre du groupe le présentant (exemple : "membres pairs", caractère plésiomorphe), mais a pu évoluer chez des espèces qui en descendent (ce caractère est conservé, mais a pu dériver en apomorphie : les pattes des Tétrapodes sont des "membres pairs" mais ne sont plus des nageoires).
La pentadactylie des êtres humains est souvent considérée comme un caractère plésiomorphe, tout droit venu de nos ancêtres tétrapodes primitifs pentadactyles.
S'il est commun à plus d'un taxon -c'est-à-dire qu'au moins une autre espèce a hérité de ce caractère, et qu'il a par ailleurs dérivé dans une tierce espèce au moins du taxon, il constitue alors une symplésiomorphie.
Une des méthodes mettant en œuvre ce principe introduit un taxon du groupe externe dans l'analyse : par exemple un téléostéen pour une étude phylogénétique des tétrapodes ; un céphalopode ou un bivalve pour une étude phylogénétique des gastéropodes. Il faut veiller à ce que les caractères des groupes externe et interne soient comparables, ce qui incite à choisir un groupe externe relativement proche du groupe d'étude. Dans l'image ci-dessous, l'arbre de départ n'est pas enraciné alors que les deux arbres ci-dessous (sous forme de graphe ou de diagramme de Venn) sont enracinés, c'est-à-dire orientés grâce aux groupes externes A ou C.
Les relations de parenté et le sens des transformations de caractères ne peuvent être inférées qu'à partir d'un arbre enraciné (autrement dit : du cladogramme).
On remarque que le choix du groupe externe (ici A ou C) modifie la topologie de l'arbre.
Il arrive que la méthode décrite ci-dessus outrepasse le principe énoncé. En effet, le taxon du groupe externe introduit dans l'analyse ne présente pas nécessairement la totalité des caractères sous l'état plésiomorphe.
Nous avons défini jusqu'à présent :
-Homologie
-Homoplasie
- apomorphie
- Synapomorphie
- Plésiomorphie
- Symplésiomorphie
D'autres méthodes d'enracinement proposent d'appliquer ce principe pour chaque caractère, en comparant le groupe d'étude à plusieurs taxons du groupe externe. Ces taxons ne sont alors pas introduits dans l'analyse :
- soit un taxon hypothétique est reconstruit ;
- soit les caractères sont polarisés en amont de l'analyse (dans le cas d'une représentation par matrice) ;
- soit les caractères sont hiérarchisés en amont de l'analyse (dans le cas d'une représentation par hiérarchie).
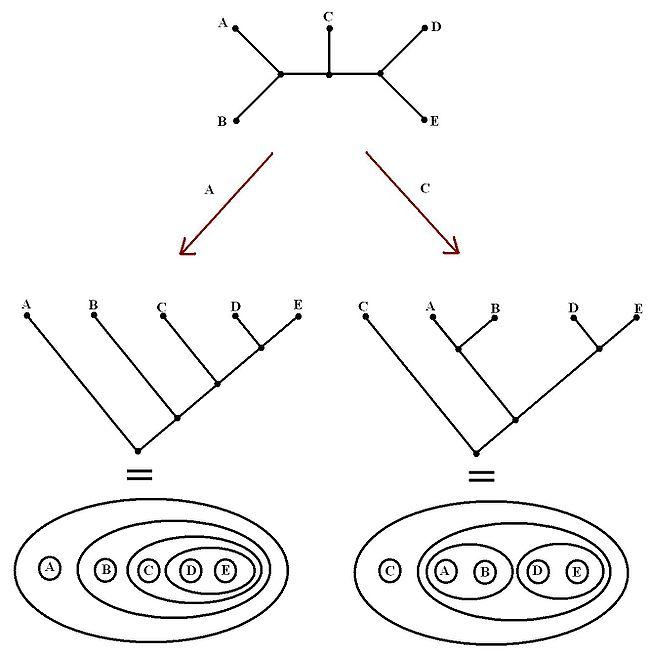
Tiré de Wikipédia article " Cladistique "
On donne des noms particuliers aux arbres selon la méthode qui a présidé à leur construction. Le terme dendrogramme est neutre et ne désigne rien d'autre qu'un arbre. Le cladogramme est un dendrogramme construit par l'analyse cladistique : il exprime les relation phylogénétiques entre taxons et ses noeuds sont définis par des synapomorphies. Le phénogramme est un dendrogramme construit par la phénétique à partir de distances entre taxons. Il exprime des degrés de similitude globale entre taxons. Le phylogramme est un cladogramme dont la longueur des branches est proportionnelle à la divergence des taxons en d'autres termes au nombre de changement de caractères ( synapomorphies, autoapomorphies, homoplasies ).
1.4 Notion de taxon
Un taxon est un regroupement d'organismes reconnu en tant qu'unité formelle . Homo sapiens est un taxon de rang spécifique ; Homo est un taxon de rang générique ; les hominidés forment un taxon de rang familial; les Primates forment un taxon de rang ordinal etc. En systématique phylogénétique , le cladogramme lui même exprime une série de taxons emboîtés. En effet, le taxon correspond à un noeud de l'arbre phylogénétique ou à l'une de ses feuilles. Il est défini par au moins un caractère dérivé propre ( ou synapomorphie ) . Il est monophylétique , c'est à dire qu'il comprend un ancêtre commun hypothétique ( chez qui la synapomorphie est apparue pour la première fois ) et la totalité de ses descendants connus . Deux taxons frères ont le même rang.
deux remarques sont à faire. Dans la mesure où les cladogrammes sont représentés, on ne s'attardera pas à spécifier le rang des taxons décrits ( classe, infra-division, sous-ordre, super famille , ...). Le rang est donné par la position de la branche dans l'arbre. L'arbre nous montre que les mammifères sont un taxon de même rang que les Sauropsides , d'un rang inférieur à celui des tétrapodes et supérieur à celui des primates . Il vaut mieux retenir la logique des rangs que les rangs eux-mêmes . En d'autres termes, il vaut mieux retenir le rang relatif ( la position dans l'arbre ) et non le rang absolu ( qui est toujours arbitraire), d'autant plus que l'ancienne logique des rangs est bouleversée par la systématique phylogénétique. La subdivision des vertébrés actuels en 5 classes ( poissons, amphibiens, reptiles, oiseaux, mammifères ) ne tient plus, non seulement parce que les trois premières de ces cinq classes ne sont pas monophylétiques , mais aussi parce qu'une classification phylogénétique ne peut pas les mettre toutes au même rang de classe. Par exemple selon l'arbre que nous présenterons, si l'on conservait le rang de classe aux mammifères, il faudrait un rang de classe aux sauropsides, de sous classe aux chéloniens et aux diapsides, d'infra classe aux lépidosauriens et aux archosauriens , de super ordre aux crocodiliens et aux oiseaux, et cela sans tenir compte des nombreuses branches fossiles.
La seconde remmarque concerne les taxons terminaux des arbres. De tels arbres phylogénétiques présentent une série de taxons emboîtés qui pourrait détailler tous les groupes à l'infini aussi loin que les connaissances puisse le permettre. Il a bien fallu couper l'arbre à une hauteur donnée dans chaque grande branche . Les choix - toujours arbitraires - sont limités par des contraintes de temps et de volume. Le fait que l'on ne détaille pas telle ou telle phylogénie ne préjuge rien ni de leur rang, ni des connaissances phylogénétiques que l'on a de ces taxons.

fig 5
Tiré de " Classification phylogénétique du vivant ", Belin
2 Les principes
2.1 Paraphylétique, monophylétique, polyphylétique.
En cladistique ( phylogénie moléculaire ), un taxon est dit paraphylétique quand il regroupe une espèce ancestrale et une partie seulement de ses descendants, et monophylétique s'il regroupe l'ensemble de ses descendants. Un groupe monophylétique définit un Clade. Le groupe des reptiles est un exemple de paraphylie , les Crocodiliens sont généalogiquement plus proches des Oiseaux ( Aves) que des autres Sauropsidés ( Chéloniens, Squamates, sphénodontiens )
Un taxon polyphylétique est un taxon défini par une ressemblance qui n'a pas été héritée d'un ancêtre dommun. Ce terme désigne des taxons qui n'ont aucune pertinence pour retracer les liens de parenté et donc l'évolution. Il est nécessaire pour cela d'utiliser la méthode cladistique. Il peut s'agir d'un terme du langage courant désignant un ensemble d'espèces présentant des caractères communs, mais regroupant de ce fait des clades d'origines variées ; un tel taxon peut également découler d'une notion scientifique invalidée par l'analyse phylogénétique voire par la systématique classique.La formation de tels taxons permet donc d'avantage de décrire un mode de vie et des adaptations communes , résultant par exemple d'une convergence évolutive et non de définir une parenté évolutive ( ex : les Algues, les Vautours, les Mammifères marins, les ongulés...).
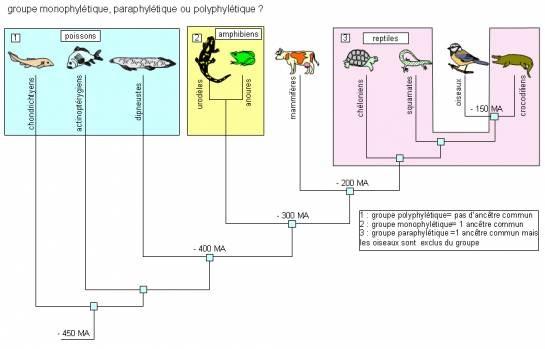
fig 6
2.2 Plésiomorphie, apomorphie
Un caractère ancestral ( homologie ancestrale ou plésiomorphie ) chez une espèce au sein d'un taxon est un caractère qui n'a pas subi de modification au cours de l'évolution
Ce caractère homologue a donc été conservé depuis le premier ancêtre du groupe le présentant ( exemple : "membre pair" , caractère plésiomorphe), mais a pu évoluer chez des espèces qui en descendent ( ce caractère est conservé, mais a pu dériver en apomorphie : les pattes des tétrapodes sont des " membres pairs mais ne sont plus des nageoires . La pentadactylie des êtres humains est souvent considérée comme un caractère plésiomorphe), tout droit venu de nos ancêtres primitifs pentadactyles.
Parmi les Angiospermes, les pollens sont ancestralement caractérisés par la présence d'une seule aperture , qui constitue ainsi un caractère plésiomorphique partagé par les dicotylédones "ancestraux" et les monocotylédones. En revanche l'apparaition de trois apertures sur le pollen est un caractère dérivé ( apomorphique ) qui détermine le clade des dicotylédones vraies ou eudicotylédones
- S'il est commun à plus d'un taxon, c'est à dire qu'au moins une autre espèce a hérité de ce caractère et qu'il a par ailleurs dérivé dans une tierce espèce au moins du taxon, il constitue alors une symplésiomorphie.
Inversément, un caractère dérivé ( ou homologie dérivée) ou apomorphie au sein d'un taxon est un caractère nouveau qui est le résultat d'une modification d'un caractère ancestral au cours de l'évolution précédent ce taxon. Ce caractère est commun à tout un groupe d'espèces et à leur espèce ancestrale.
- S'il est spécifique à un seul taxon, il s'agit plus précisément d'une autoapomorphie ( c'est notamment le cas si ce taxon constitue un groupe terminal, cul de sac évolutif par exemple.
- S'il est commun à plus d'un taxon, il constitue une synapomorphie, qui détermine un groupe monophylétique, ou holophylétique, donc un clade en phylogénétique.. C'est le cas des quatre membres marcheurs des Tétrapodes, qui en font un clade, même si ce caractère a pu régresser totalement ( cas des serpents ou des orvets, qu'on classe dans les Tétrapodes. Des caractéristiques osseuses vestigiales de ces membres y subsistent) .
2.3. La cladistique
L'analyse cladistique vise à reconstruire la phylogénie d'un taxon par distinction, au sein d'un caractère, de l'état primitif ( plésiomorphe ) de l'état dérivé ( apomorphe ). Ces qualités ne sont valables qu'au sein du taxon. Le premier, Hennig a compris qu'on ne pouvait pas regrouper les espèces sur la base d'un caractère primitif puisque celui-ci est déjà présent en dehors du taxon étudié. Au sein de ce taxon, seuls les états de caractères dérivés partagés ( synapomorphies ) sont des signes d'apparentement exclusif; les regroupement sur base de d'états dérivés partagés conduisent donc à la création de groupes monophylétiques. On parle de caractères dérivés propres lorsque sous cette rubrique sont exposésdes états de caractères dérivés propres au groupe dont il est question , et partagés par les membres du groupe. Ce sont les synapomorphies, ou encore les innovations du groupe.
2.4. Qu'est-ce que la parenté ?
Chercher la parenté est chercher le groupement et non l'ancêtre . Il s'agit de chercher pour un groupe ( ou une espèce ) donné(e), le groupe avec lequel il ( elle ) partage un ancêtre exclusif, c'est à dire qu'il(elle) ne partage pas avec aucun autre groupe de l'échantillon . Cela ne conduit pas à chercher l'ancêtre au sens propre ( au sens génétique ), car celui-ci restera à jamais innaccessible. Cela conduit plutôt à déduire certains caractères que devrait posséder cet ancêtre à partir des caractères dérivés qui définissent le groupe, les synapomorphies.
2.5. Comment trouve-t-on la parenté
Dans une approche hennigienne classique, la construction d'une classification phylogénétique a pour première étape de polariser les caractères, c'est à dire d'en définir l'état primitif( ou plésiomorphe ) de(s) état(s) dérivé(s) ou (apomorphes). La seconde étape consiste à déduire une parenté ( c'est à dire construire un groupe )sur le partage d'états dérivés.
Comment trouve-t-on la polarité d'un caractère ? On apère à partir de deux critères : le critère extra-groupe et le critère ontogénétique, qui sont appliquès en partant d'une collection d'espèce que l'on a échantillonnées afin de connaître leurs relations de parenté . Cette collection constitue le groupe d'étude , ou groupe intérieur ( ingroup en anglais ); il est supposé monophylétique, ce qui n'implique pas que l'on sache à l'avance les relations de parenté entre ses membres. Le but est de déterminer au sein du groupe d'étude , quel est l'état primitif d'un caractère donné et quel est son état dérivé.
Le critère extra-groupe ( fig suiv.), le plus utilisé consiste à trouver une espèce ( ou un groupe d'espèces) dont on soit sûr qu'elle ( il) est extérieur(e) au groupe d'étude, c'est à dire qu'elle (il) ne viendrait en aucun cas se brancher à l'intérieur. Cette espèce est appelée groupe extérieur ou extragroupe ( outgroup en anglais ) et sert à raciner l'arbre. Car, on sait que l'ancêtre commun situé à la racine est l'ancêtre le plus ancien si le groupe extérieur est monophylétique, contenu dans le groupe extérieur si celui-ci est paraphylétique. Sa définition n'utilise aucune information a priori sur les parentés possibles dans le groupe d'étude et doit être considérée comme faisant partie des hypothèses de départ. Si, chez une ou plusieurs espèce(s) du groupe d'étude ( par exemple Crocodile et Alligator ), l'état du caractère ( écaille ) est le même que dans le groupe extérieur ( présence d'écailles chez la tortue), cet état sera considéré comme primitif ( plésiomorphe ) dans le groupe d'étude. Si, chez d'autres espèces (poulet, autruche ), l'état est différent ( plumes ) de celui présent chez le groupe extérieur ( écaille), cet état sera considéré comme dérivé ( ou apomorphe ).
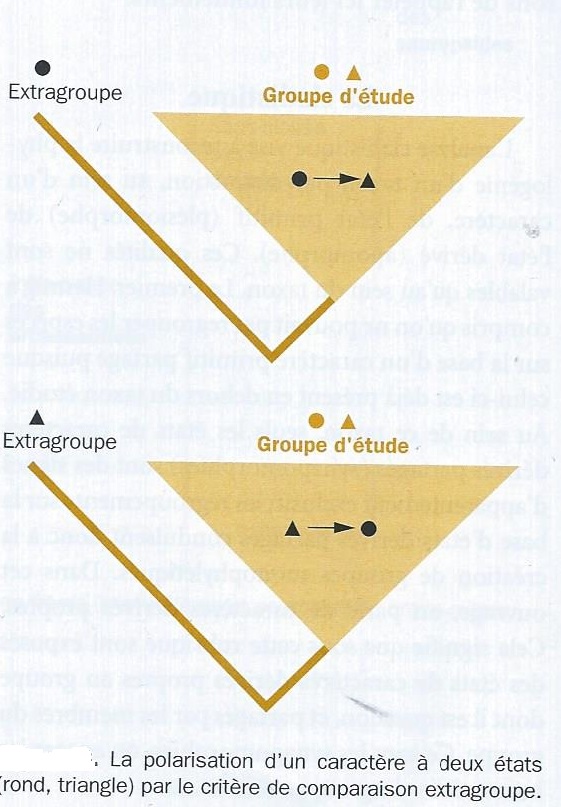
fig 7
3. Exemple traité
Prenons un exemple biologique simple qui permet de replacer les éléments précédents les uns par rapport aux autres. Le cas le plus simple en reconstruction phylogénétique comprend un groupe d'étude de trois espèces et un groupe extérieur d'une espèce.
Soit les espèces :
- homme ( Primate).
- gris du Gabon ( Oiseau ).
- molosse du Cestoni ( Chauve-souris ).
La question est de savoir quelles sont les deux espèces les plus proches entre elles qu'elles ne le sont de la troisième, le cas présent n'offre que trois possibilités :
- homme ( oiseau, chauve-souris)
- oiseau ( homme, chauve souris)
?- chauve-souris ( homme oiseau )
?Pour choisir nous l'une d'entre elles, nous disposonsd'une collection de caractères observés. Examinons ces caractères observés afin de savoir lequel de ces trois arbres ils soutiennent.
?Il faut tout d'abord polariser les états de caractères. Nous sommes en mesure de faire intervenir une quatrième espèce , la truite , dont on sait qu'elle est extérieure à l'ensemble ( homme oiseau chauve-souris), grâce à une série d'informations extérieures à cette analyse. Par exemple le fait que ces trois dernières espèces soient des amniotes ( l'embryon se développe dans la poche amniotique ) exclut la truite qui ne présente pas d'amnios. La truite est donc ici le groupe extérieuret servira à raciner l'arbre. Remarquons bien que ce choix n'utilise aucune information anticipant les relations de parenté entre homme, chauve souris et oiseau. Contrairement à une erreur courante , la définition du groupe extérieur n'a rien de circulaire.
?dans le tableau qui suit nous examinons 6 caractères : 1. Mâchoires: deux états : présence, absence; 2. type d'appendices pairs ; deux états : nageoires rayonnées, membre chiridien; 3.dents; deux états: présence, absence; 4.constitution de la mandibule; deux états : un seul os, le dentaire, plusieurs os ; 5. Réserves vitellines : oeuf alolécithe avec très peu de quantités de vitellus, oeuf télolécithe avec de grandes quantités de vitellus se séparant du cytoplasme. 6. Appendices pairs antérieurs ; deux états : transformés en ailes, absence d'ailes. On constate tout d'abord que tous n'ont pas la même qualité informative.
Caractères non informatifs.
?Le caractère 1, la présence de mâchoires, est trouvé chez la truite. Par convention, on code 0 pour l'état de caractère trouvé dans le groupe extérieur. On trouve une mâchoire pour chacune des quatre espèces. la présence d'une mâchoire sera donc codée 0 pour les quatre. La simple présence de mâchoires ne peut pas discriminer deux des trois espèces du groupe d'étude et constitue donc un caractère non-informatif pour le présent échantillon d'espèces.
| Homme | Chauve-souris | Oiseau | Truite | |
| 1. Mâchoire | 0 | 0 | 0 | 0 |
| 2.Membres | 1 | 1 | 1 | 0 |
| 3.Dents | 0 | 0 | 1 | 0 |
| 4. Constitution de la mandibule | 1 | 1 | 0 | 0 |
|
5.Réserves vitellines de l'oeuf |
1 | 1 | 0 | 0 |
| 6. Ailes | 0 | 1 | 1 | 0 |
fig 8
Matrice de caractères ( voir texte ). 1: mâchoire ( présence, absence), 2: type d'appendices pairs ( nageoire rayonnée, membre chiridien), 3 : dents ( présence, absence ), 4 : constitution de la mandibule ( faite de plusieurs os dont le dentaire, mandibule exclusivement dentaire ), 5 : réserves vitellines de l'oeuf ( énormes, quasi nulles ), 6 : appendice pair antérieur ( aile, pas d'aile ).
Le caractère 2, types d'appendices est codé 0 chez la truite qui présente bien des appendices pairs mais sous forme de nageoires rayonnées ( état primitif ). Homme, chauve-souris et oiseau ont tous des appendices pairs sous forme de membres marcheurs , et cet état est codé 1 ( état dérivé). Cependant ce caractère n'est pas informatif pour notre échantillon, car il ne permet pas de discriminer 2 des 3 espèces du groupe sous étude. Le Le caractère 3 , dents est codé 0 chez la truite . Par définition, la présence de dents est donc l'état primitif . Chez l'homme et la chauve souris on trouve des dents , et cet état est codé 0 ( état primitif ). L'oiseau présente une absence de dents et cet état est codé 1 . Nous sommes ici en présence d'un caractère qui discrimine deux des trois espèces du groupe d'étude, il paraît donc informatif . Cependant il ne permettra pas de produire un regroupement dans le groupe d'étude ; En effet, la méthode cladistique ne permet pas de regroupement sur la base de caractères primitifs partagés. Homme et chauve souris ne peuvent pas être regroupés parce qu'il présentent des dents, car ces dents ont déjà été trouvées chez la truite. L'acquisition du caractère dérivé, l'absence de dents n'est observée ici que chez l'oiseau . C'est ce que l'on appelle une autoapomorphie , c'est à dire un caractère dérivé propre à un seul des taxons ; ce caractère n'est donc pas informatif du point de vue de la méthode employée ; il nous dit que l'oiseau est un oiseau.
Caractères informatifs
Le caractère 4, relatif à la constitution de la mandibule , est codé 0 chez la truite et chez l'oiseau où l'on trouve une mosaïque d'os ( état primitif ) et 1 chez l'homme et la chauve souris où l'on ne trouve qu'un seul os par hémimandibule , l'os dentaire ( état dérivé ). Ce caractère permet le regroupement de ces deux dernières espèces , car elles partagent le même caractère dérivé. La situation est la même pour le caractère 5 relatif aux réserves vitellines de l'oeuf : la truite et l'oiseau ont des oeufs très riches en vitellus qui se sépare du cytoplasme cellulaire ( état primitif car trouvé chez la truite , tandis que la chauve souris et l'homme ont un oeuf pratiquement dépourvu de vitellus ( état dérivé). Le caractère 6, membres antérieurs transformés en ailes permet aussi de faire un regroupement dans le groupe d'étude mais pas le même : ( chauve souris oiseau) . Deux remarques s'imposent alors 1. seuls sont informatifs pour la méthode utilisée les caractères présent au moins deux fois à l'état dérivé au sein du groupe d'étude; et 2. les caractères peuvent parfaitement se contredire ( présence d'homoplasies, convergences évolutives, réversions...).
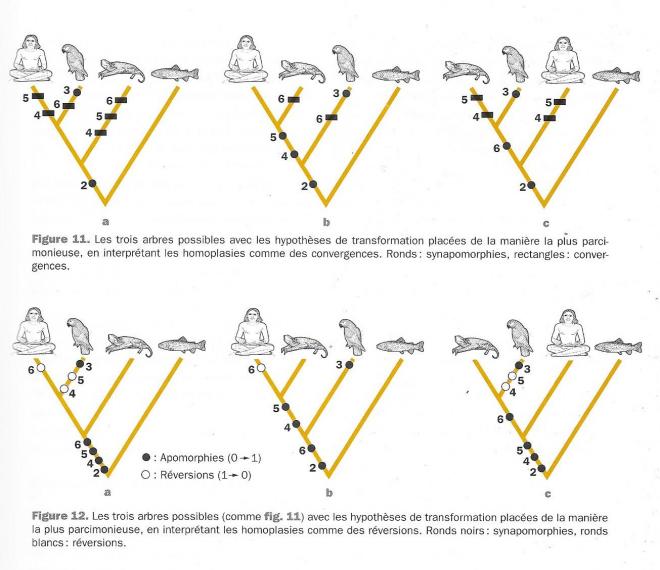
?Superposons à présent nos caractères sur chacun des trois arbres de parenté possibles ( cladogramme fig 9 ). Rappelons qu'une parenté, c'est à dire un groupe, ou encore un noeud de l'arbre, ne peut être fondée que sur base d'un caractère dérivé partagé, c'est à dire d'un rond noir commun. Au niveau du rond noir , on passe de l'état 0 à l'état 1. L'arbre 9a montre que le noeud ( homme,oiseau) n'est fondé sur aucun rond noir commun. L'arbre 9b présente le noeud ( chauve souris homme ) qui, lui est argumenté par les caractères 4 et 5 : mandibule constituée du dentaire et oeuf alécithe sont chacun un état dérivé commun, ou synapomorphie, qui témoigne d'une parenté possible entre homme et chauve souris. L'arbre 9c présente le groupement de l'oiseau avec la chauve-souris, fondé sur le caractère dérivé que constitue la présence d'ailes. Deux caractères plaident donc pour le regroupement [oiseau(homme, chauve souris)], un seul pour [homme,(oiseau, chauve souris)], et aucun pour ( chauve-souris,(homme, oiseau)].
L’homoplasie est la similitude d’un état de caractère chez différentes espèces qui, contrairement à l’homologie, ne provient pas d’un ancêtre commun1. Il existe différents types d’homoplasie : la convergence, le parallélisme et la réversion.
- la convergence : ressemblance apparue indépendamment dans divers taxons sans lien de parenté.
- le parallélisme : ressemblance apparue chez des taxons relativement proches.
- la réversion : un état dérivé d’un caractère revient à un état ancestral (antérieur).
L'homoplasie de fonction est une analogie, c'est-à-dire une ressemblance de caractères remplissant les mêmes fonctions biologiques.
Il faut alors calculer le nombre de transformations des caractères que nécessite chacun des arbres. Le caractère 6 contredit les caractères 4 et 5. Deux positions sont à envisager face à l'homoplasie , on peut supposer indifféremment pour un caractère homoplasique la convergence ( fig 11) ou la réversion ( fig 12 ce sont les mêmes trois arbres ). Ces deux options reviennent au même en terme d'évènements dans l'arbre, elles diffèrent simplement par la position de ces évènements . Prenons tout de suite l'arbre le plus économique en évènement évolutifs ( la figure 11b ) : le rond noir symbolise la synapomorphie, c'est à dire l'homollogie secondaire, celle de la descendance . Sur le segment de branche réunissant l'homme, la chauve-souris et l'oiseau, le caractère 2 passe de son état 0 à son état 1 ( ce sera la même chose pour chacun des trois arbres ), ce sera déjà un pas . Sur le segment de branche qui réunit homme et chanve=souris , les caractères 4 et 5 changent d'état ( 0 -->1) ; on compte ici deux pas supplémentaires . Le caractère 3 se transforme de 0 à 1 sur la branche qui mène à l'oiseau ( autoapomorphie) , on compte alors un quatrième pas. On peut supposer une convergence en ce qui concerne le caractère 6, c'est à dire deux transformations indépendantes 0 vers 1 , d'une part sur la branche qui mène à l'oiseau et d'autre part sur la branche qui mène à la Chauve-souris ( voir les rectangles) , cette convergence coûte deux pas, ce qui donne 6 pas au total pour cet arbre. On peut supposer aussi une réversion qui nécessite deux pas ( voir les ronds blancs dans l'arbre 12b qui est le même) : une transformation de 0 vers 1 sur la branche qui mène à l'homme, à la chauve -souris et l'oiseau ( rond noir ) puis une réversion de 1 vers 0 ( rond blanc) sur la branche qui mène à l'homme , l'homme aurait perdu ses ailes ! Le réversion amène également à un nombre de pas égal à 6 sur l'arbre 12b.
Considérons ensuite l'arbre 11a, il faut supposer une synapomorphie ( rond nois pour le caractère 2), une auto-apomorphie ( rond noir pour le 3) et 3 homoplasies, ( rectangles pour 4,5,6). Chaque homoplasie coûte deux pas, qu'il s'agisse d'une convergence ( 11a ) ou d'une réversion ( 12a): celà amène le nombre de pas total à 8 .Considérons enfin l'arbre 11c , il faut supposer synapomorphies ( caractères 2 et 6), une autoapomorphie ( caractère 3 ) et deux homoplasies ( caractères 4 et 5 ) chaque homoplasie coûtant 2 pas , ce qui amène le nombre total à 7. Le critère de choix de l'arbre est celui du nombre minimal de pas évolutifs conformément au principe de parcimonie ou principe d'économie d'hypothèses. on choisit l'arbre 11b qui est le plus parcimonieux ( ou 12b sous la forme " réversions"). La parenté choisie est celle de l'homme et de la chauve-souris. L'arbre le plus parcimonieux nous révèle que poils et mamelles ont été acquis chez un ancêtre commun exclusif à l'homme et la chauve-souris. Pour ce qui est de l'homoplasie, les ailes ont été acquises deux fois séparément ( 11b) ou ont fait l'objet d'une disparition secondaire chez l'homme (12b) . Un arbre comprenant plus d'espèce nous montrerait que c'est la première des deux hypothèses qui est à retenir.
4. Résumé du déroulement de la construction d'une classification phylogénétique
La reconstruction des liens de parentés ne résulte pas d'une découverte fortuite ou révélée , mais de décisions préalables réfléchies qui sont autant de postulats :
1. Une question posée au départ : Etape de définition du problème phylogénétique, c'est à dire délimitation du groupe d'intérêt ( ingroup ) .
2. Le choix d'un échantillon d'organismes : propre à répondre à cette question , c'est l'échantillonage taxinomique. Cette étape consiste aussi à choisir un ou plusieurs points de référence , le(s) extragroupe(s). Ces organismes sont postulés extérieurs au groupe d'intérêt , c'est à) dire extérieurs à tout ceux dont on veut définir les relations de parenté. ces extragroupes serviront à enraciner le(s) futur(s) arbre(s).
3. Le choix et/ou la détection des caractères qui seront informatifs pour la question posée, c'est l'échantillonage des caractères .
4. Le codage et la construction de la matrice des aractères. Ce codage comprend l'étape d'établissement des homologies primaires ( principe de connexion, alignement des séquences ) et l'étape de polarisation des caractères ( critère du groupe extérieur ou critère ontogénétique ) . Puis, l'inscription du fruit de cette analyse dans une matrice de caractères.
5. L'exploration des arbres possibles. A un nombre fixé de taxons dans la matrice correspondante correspond un nombre précis d'arbres possibles . Pour quatre taxons dont un est préalablement considéré comme extra groupe, il existe trois arbres racinés possibles. pour 5 taxons, il existe 15 arbres racinés possibles . L'idéal est de les évaluer tous au regard des informations contenues dans la matrice.
6. L'évaluation de chacun de ces arbres au regard de la matrice .
7. Le choix d'un critère de choix d'un arbre : ce critère est, en cladistique, la parcimonie . On choisit l'arbre le plus parcimonieux c'est à dire l'arbre qui requiert le plus petit nombre d'évènements pour les états de caractères présentés par les organismes, mais il faut savoir qu'il existe, hors la cladistique, d'autres critères de choix d'un arbre ( approche probabiliste, méthode des moindres carrés)
8. Le choix d'un arbre : ce sera l'arbre le plus court, c'est à dire celui qui nécessite le moins d'hypothèses de transformations de ces caractères. Cet arbre permet :
- l'identification des états des caractères qui définissent chacun des noeuds ( = clades) de l'arbre obtenu ( les synapomorphies = homologies de descendance ou homologies secondaires )
- l'identification des état homoplasiques et éventuellement de conclusions sur le mode d'évolution des caractères.
- la création d'une classification phylogénétique, c'est à dire l'assignation d'un nom de groupe à chaque noeud ( ! clade) fiable de l'arbre, c'est ce que l'on appelle le ranking .
remarque : les étapes 2,3 et 4 sont parfois confondues. En effet, il arrive que la variabilité des caractères amène à modifier l'échantillonage taxonomique. En phylogénie moléculaire, les étapes 4,5,6,7,8 sont parfois confondues. En effet, pour un jeu de séquences appartenant aux espèces que nous avons choisies ,certains logiciels calculent directement l'alignement des séquences qui donneront l'arbre le plus court.
UN ARBRE GENERAL DE LA VIE CONSTRUIT A PARTIR DE CETTE METHODE
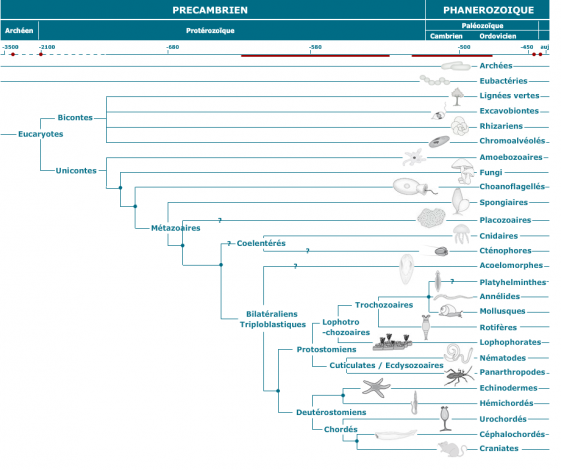
il semble intéressant de définir certains termes de ce tableau :
1) Parmi les procaryotes, on distingue deux groupes :
- Les archées : Les archées sont des micro-organismes unicellulaires procaryotes (il n'y a pas de noyau dans la cellule). Leur taille varie entre 0,1 et 15 microns et elles vivent dans à peu près tous les milieux. On trouve des archées dans des milieux extrêmes (anaérobies, à forte salinité, très chauds ou à grande profondeur)
.Les analyses génétiques et les méthodes de classification de la phylogénétique ont conduit à les séparer des bactéries, les archées étant aussi différentes d'elles que des eucaryotes (plantes et animaux notamment). Depuis la création de ce groupe, l'ensemble des organismes vivants se divise en trois : les archées, les bactéries et les eucaryotes.
Les archées sont un domaine (et aussi un royaume) qui est divisé en cinq branches reconnues, mais elles identifient plus que ça. Parmi ces groupes, Crenarchaeota et Euryarchaeota sont les plus étudiés. La classification des archées est encore difficile, car la grande majorité n'a jamais été étudiée en laboratoire et n'a été détectée que par l'analyse de leurs acides nucléiques dans des échantillons prélevés dans l'environnement.
Les Archaea et les bactéries sont assez semblables en taille et en forme, bien que certaines archées aient des formes très inhabituelles, telles que les cellules aplaties et carrées de Haloquadratum walsbyi. Malgré cette similitude visuelle avec les bactéries, les archées possèdent des gènes et plusieurs voies métaboliques plus proches de celles des eucaryotes, en particulier dans les enzymes impliquées dans la transcription et la traduction. D'autres aspects de la biochimie des archées sont uniques, tels que les éthers lipidiques de leurs membranes cellulaires. Les archées exploitent une variété de ressources beaucoup plus grande que les eucaryotes, des composés organiques communs tels que les sucres, à l'utilisation de l'ammoniac des ions métalliques ou même de l'Hydrogène comme nutriments. Les archées tolérantes au sel (haloarchées) utilisent la lumière du soleil comme source d'énergie, et d'autres espèces d'archées fixent le carbone; cependant, à la différence des usines et des cyanobactéries, il n'y a aucune espèce connue d'archées qui est capable des deux. Les archées se reproduisent de manière asexuée et sont divisées par fission binaire, fragmentation ou bourgeonnement; contrairement aux bactéries et aux eucaryotes, aucune espèce d'archées formant des spores n'est connue.
Initialement, les archées étaient toutes méthanogènes ou extrêmophiles (azote) vivant dans des environnements hostiles tels que les sources chaudes et les lacs salés, mais les archées se trouvent maintenant dans les habitats les plus divers, tels que le sol, les océans, les marais et le côlon humain. Les archées sont particulièrement nombreuses dans les océans, et celles qui font partie du plancton pourraient être l'un des groupes d'organismes les plus abondants de la planète. Ils sont actuellement considérés comme une partie importante de la vie sur Terre et pourraient jouer un rôle important à la fois dans le cycle du carbone et dans le cycle de l'azote. On ne connaît pas d'exemples clairs d'archées pathogènes ou parasitaires, mais elles sont généralement mutualistes ou diners. Les exemples sont les archées méthanogènes qui vivent dans les intestins des humains et des ruminants, où ils sont présents en grandes quantités et contribuent à digérer les aliments. Les archéobactéries ont leur importance dans la technologie, il y a des méthanogènes qui sont utilisés pour produire du biogaz et dans le processus de purification de l'eau, et les enzymes des archées extrêmophiles sont capables de supporter des températures élevées et des solvants organiques, donc utilisés en biotechnologie.
De nombreuses études ont confirmé le caractère monophylétique de ce groupe. Ces microorganismes ressemblent par leur forme aux bactéries, mais d'un point de vue moléculaire, si certains de leurs traits les rapprochent des bactéries, d'autres les rapprochent plutôt des eucaryotes. Il n'est donc pas possible de voir les archées comme étant des ancêtres des bactéries.
Le classement des archées, et des procaryotes en général, est à la fois en évolution rapide et un domaine litigieux. Sur la base de critères uniquement métaboliques, les archées ont été divisées en quatre grands groupes selon qu'elles sont méthanogènes, halophiles, thermophiles ou sulfo-dépendantes.
Les systèmes de classifications actuels visent à organiser les archées en groupes d'organismes qui partagent des caractéristiques structurelles et des ancêtres communs. Ces classifications s'appuient fortement sur l'usage de la séquence des gènes de l'ARN ribosomique pour révéler les relations entre les organismes (phylogénétique moléculaire). La plupart des archées cultivables sont membres de deux principaux embranchements : Euryarchaeota et Crenarchaeota. D'autres groupes ont été provisoirement créés. Par exemple, les espèces propres Nanoarchaeum equitans, qui ont été découvertes en 2003, ont été classées dans un nouveau phylum : Nanoarchaeota. Un nouveau phylum, Korarchaeota, a également été proposé ; il contient un petit groupe d'espèces thermophiles inhabituelles qui partagent les caractéristiques des deux principaux embranchements, mais qui sont plus étroitement liées aux Crenarchaeota. Récemment mises en évidence, d'autres espèces d'archées, tels que les archaeal Richmond Mine acidophilic nanoorganisms (ARMAN), qui ont été découvertes en 2006, sont liées seulement de loin aux autres groupes antérieurement connus. Le superphylum TACK a été proposé en 2011, pour regrouper les Thaumarchaeota, Aigarchaeota, Crenarchaeota et Korarchaeota .
L'archée Loki, identifiée en 2015 par son génome qualifié de Candidatus en nomenclature bactérienne, appartiendrait, du point de vue phylogénétique, à l'embranchement le plus proche des eucaryotes22,23. De nouvelles lignées d'archées, apparentées à Loki, ont été identifiées dans les sédiments aquatiques par analyse métagénomique. Les archées Odin, Thor et Heimdall, formeraient, avec Loki, le super-embranchement Candidatus "Asgard", proposé en 2017 d'après le nom du royaume des dieux de la mythologie nordique
- Les eubactéries :
Nous développerons ce groupe dans une prochaine section " microbiologie "
2) Parmi les eucaryotes ( cellules à vrai noyau) on distingue deux groupes :
- Bicontes : un organisme biconte est un organisme eucaryote dont les cellules possèdent deux flagelles dont un flagelle classique et un flagelle mastigonème (.Les mastigonèmes (il y en a toujours plusieurs) sont de fines digitations (prolongements tubulaires) de la membrane d'un flagelle). Un autre trait commun aux bicontes est la fusion de deux gènes en un seul : le gène de la thymidylate synthase (TS) et celui de la dihydrofolate réductase (DHFR) codant donc ici pour une unique protéine ayant une double fonction (Cavalier-Smith, 2003). Ces deux gènes sont traduits séparément chez les organismes unicontes. Certaines recherches suggèrent que certains unicontes (unicellulaire eucaryote possédant un unique flagelle) ont été les ancêtres des opisthocontes et des Amoebozoa, et que les bicontes sont à l'origine des Archaeplastida, Excavata, Rhizaria, et Chromalveolata. Thomas Cavalier-Smith pense, par ailleurs, que les Apusozoa, qui sont classiquement considérés comme incertae sedis (groupe au classement incertain), sont en fait des bicontes.
Les bicontes regroupent quatre embranchements: la lignée dite "verte", les excavobiontes, les rhizariens et les chromoalvéolés.
- La lignée verte : Ils pratiquent donc la photosynthèse, au minimum par le biais de la chlorophylle a (un pigment photosensible), ce qui leur permet de synthétiser des sucres mis en réserve sous forme de grains d'amidon. Les membres de la lignée verte sont les descendants de l'être qui a réalisé la première endosymbiose chloroplastique.
Ce groupe rassemble quelque 283.000 espèces du monde entier, qui se répartissent entre les glaucophytes (13 espèces, comme Cyanophora paradoxa) et les métabiontes (par exemple les fougères, la laitue de mer ou encore les pommiers).
La lignée verte désigne donc l'ensemble des végétaux (plantes et algues) possédant un plaste vert issu d'une endosymbiose primaire (toutes les plantes vertes et la plupart des algues vertes) ou secondaire avec une algue verte (les autres algues vertes : les Chlorarachniophytes et les Euglénophytes).
Du point de vue de la phylogénie des hôtes de l'endosymbiose, c'est un groupe polyphylétique.
Du point de vue de la phylogénie des endosymbiotes (plastes), c'est un groupe monophylétique.
- Les excavobiontes : les excavata (Excavés, ce qui signifie creusés) ou excavobionta sont un règne d'Eukaryotes unicellulaires (« protistes ») hétérotrophes et généralement flagellés. Le type ancestral se caractérise par la présence d'une zone creusée, le cytostome, permettant l'ingestion de fines particules alimentaires. De nombreux excavates sont anaérobies ou microaérophile et contiennent des organites particuliers et spécialisés : les mitosomes et les hydrogénosomes, EX : Diplomonade
De nombreux Excavobiontes sont des parasites de l'homme ou d'autres animaux. Ex : Trypanosomes, Giardia
Certains possèdent des plastes (endosymbionte secondaire) et sont photosynthétiques : Euglénoides
Les Euglenobionta et les Metamonada forment deux groupes monophylétiques.
- Les rhizariens :Les rhizaires aussi appelés rhizariens (ou Rhizaria) sont un groupe d'organismes eucaryotes, le plus souvent unicellulaires. Ils peuvent vivre dans des eaux oligotrophes en s'associant symbiotiquement à des algues
Les Rhizaires ont une morphologie amiboïde.
Parce qu'ils ont une forme amiboïde, ils ont été historiquement classés parmi les Amoebozoa.
Leur positionnement est hésitant puisqu'on peut les présenter comme groupe frère des Excavata puis des Straménopiles (ou plus précisément les Halvaria.....pour mémoire).
Selon ce dernier point de vue il s'agit d'une division, non plus des Chromistes, mais du sous-règne des Harosa.
Selon World Register of Marine Species (21 juillet 2017) :
Embranchement des Cercozoa
Embranchement des Foraminifera
Embranchement des Radiozoa
Traditionnellement ce sont des protiste
Les chromoalvéolés portent également le nom de lignée brune : le clade présumé des Chromistes (ou la lignée brune selon Le Guyader et Lecointre) est composé d'êtres vivants uni- et pluricellulaires, photosynthétiques pour la plupart, bien que des organismes vivants autrefois classés parmi les « champignons inférieurs », et donc hétérotrophes, soient aujourd'hui inclus dans ce taxon.
La pertinence de ce clade est remise en cause par des études récentes rapprochant les Stramenopiles des Rhizaria ( pour mémoire). Il a donc été proposé d'utiliser le terme de Chromista pour désigner l'ensemble présumé monophylétique formé par les Harosa et les Hacrobia à la place du terme de Chromalveolata au sens large. Elles sont moins diversifiées que les algues vertes, mais plus que les algues rouges
- Unicontes : Le taxon Unikonta ou Unikonte (ou encore du nom vernaculaire Uniconte) représente les cellules eucaryotes possédant à l'origine un unique flagelle postérieur. Les recherches actuelles suggèrent que les Opisthokontes et les Amoebozoaires ont pour ancêtres des organismes unicellulaires unicontes, et le clade regroupant Amoebozoaires et Opisthokontes est de ce fait nommé Unikonta. De même, des unicellulaires bicontes (eucaryotes possédant deux flagelles) sont les ancêtres des Archaeplastida (Plantae), Excavata et Chromalveolata (incluant Rhizaria), et l'ensemble de ces taxons est réuni dans le taxon des Bicontes. Ce taxon a été proposé, en tant que clade, par le professeur en biologie Thomas Cavalier-Smith.
Les organismes unicontes possèdent également une triple fusion de gènes n'existant pas chez les bicontes, ainsi qu'un unique centriole (quelques unicontes possèdent 2 centrioles mais leur origine est différente de celles des bicontes, indiquant ici une convergence évolutive). Les trois gènes ayant fusionné chez les unicontes, mais pas chez les procaryotes ni chez les bicontes, codent des enzymes intervenant dans la synthèse de nucléotides pyrimidine : carbamoyl phosphate synthase, dihydroorotase, aspartate carbamoyltransferase (Cavalier-Smith 2003). Il semble probable que cette triple fusion soit le résultat de 2 doubles fusions successives, un événement rare dans l'évolution et qui a peu de chances d'être réversible. Cette situation est en faveur de la théorie proposant un ancêtre commun aux Opisthocontes et aux Amoebozoa.Remarquons que : les opisthocontes (Super règne faisant partie des unicontes) ) Opisthokonta constituent un groupe d'eucaryotes regroupant des organismes très divers en apparence, résultant du rapprochement récent de plusieurs taxons, dont les champignons et les métazoaires. Il est fondé sur l'étude de plusieurs gènes analysés séparément, ce qui confère à l'existence des opisthocontes une certaine fiabilité. À partir de cette mise en évidence moléculaire, d'autres synapomorphies sont apparues comme évidentes, confirmant qu'il s'agit bien d'un groupe monophylétique. Les opisthocontes se seraient séparés des autres eucaryotes il y a un milliard d'années.s...
nous mentionnons en particulier les amoebozoaires qui ne sont pas encore traités dans ce site :
Les amoebozoaires (Amoebozoa) constituent un grand groupe de protozoaires, la majorité se déplaçant par vagues cytoplasmiques internes. Leur pseudopode est appelé lobopodia. La plupart sont unicellulaires, et sont souvent rencontrés dans les sols et les milieux aquatiques, où ils cohabitent avec d'autres organismes. Plusieurs sont pathogènes. Les Amoebozoa peuvent aussi être multicellulaires et peuvent produire des spores, généralement visibles à l'œil nu.
La taille des Amoebozoa varie beaucoup. Beaucoup font seulement 10-20 μm, mais certains font la taille des plus grands protozoaires. La fameuse espèce Amoeba proteus peut atteindre 800 μm de longueur et est souvent étudiée comme représentant une cellule (en raison de sa taille). Quand ils s'agglutinent, il peuvent couvrir de très grandes surfaces.
Morphologie :La cellule est généralement divisée en une masse granuleuse centrale, appelée endoplasme, et une couche extérieure, appelée ectoplasme. Au cours de la locomotion, des flux endoplasmiques se produisent à l'avant et à l'arrière de la cellule. De nombreux amibes ont un antérieur et un postérieur, la cellule fonctionne comme un seul pseudopode. En général, ils produisent de nombreuses projections appelées subpseudopodia, qui ne sont pas directement impliquées dans la locomotion.
D'autres amoebozoa peuvent avoir de multiples pseudopodes indéterminés, qui sont plus ou moins tubulaires et sont pour la plupart remplis de granulés endoplasmiques. Le flux de masse des cellules dans l'un des principaux pseudopodes, et les autres se rétracte seulement si elle change de direction. En plus de quelques formes comme Amoeba et Chaos, cela concerne la plupart des amibes qui produisent des coquillages. Ces être peuvent être composés de matériaux organiques, comme dans l'Arcella, ou de particules collectées et collés, comme dans Difflugia, avec une seule ouverture par laquelle le pseudopodia émerge.
Le principal mode de nutrition est la phagocytose: la cellule entoure de potentielles particules d'aliments, puis la vacuole se referme et l'absorbe. Certaines amibes ont un postérieur appelé une ampoule uroid, qui peut servir à accumuler des déchets, qui se détache périodiquement du reste de la cellule. Lorsque la nourriture est rare, la plupart des espèces peuvent former des kystes pouvant alors devenir aérien et les emmener vers de nouveaux environnements. Ces structures sont appelées spores, et la forme de structures appelées pétiolées organes de fructification ou sporanges. (Ce texte est mal traduit de l'anglais).
La plupart des Amoebozoa n'ont pas de flagelles et, plus généralement, ne sont pas soutenus par un réseau microtubulaire sauf pendant la mitose. Toutefois, des flagelles se produisent chez certains Archamoebae, et de nombreux moules produisent des gamètes biflagellate. Les flagelles sont généralement ancrées à un cône de microtubules, ce qui suggère une relation étroite à l'opisthokonta. Les mitochondries caractéristique de branchement ont des cristas tubulaires, mais ont été perdu à l'archamoebae.
l'ensemble des autres unicontes sont ou seront traités dans ce site.
Chapitre à suivre : La phénétique
Date de dernière mise à jour : 07/01/2019
Ajouter un commentaire