

Annélides
9. LES ANNELIDES
Triploblastiques coelomates bilatériens protostomiens lophotrochozoaires eutrochozoaires spiraliens

|
On retrouve des animaux ayant la forme de vers (vermiformes) et ne possédant pas de pattes dans environ 14 embranchements d'invertébrés Protostomiens. Le mot ver désigne donc un grand nombre d'animaux qui ne sont ni des Mollusques ni des Arthropodes. Ces 14 embranchements dérivent tous d'un ancêtre commun Plathelminthe, mais ont peu d'autres choses en commun. Le lombric est l'Annélide qui vous est le plus familier mais il n'est pas nécessairement représentatif des formes variées chez les différentes espèces dans l'embranchement . Les Annélides sont un des embranchements actuellement les plus diversifiés (le 7ième) et possèdent plusieurs adaptations importantes qui ont assuré leur succès. Comme les membres de l'embranchement des Arthropodes, le plus diversifié actuellement, ils dérivent d'un ver plat. Leur caractéristique la plus évidente lorsque l'on examine leur anatomie externe est leur segmentation (ou métamérisation). Un examen attentif de l'anatomie interne révèle que les organes internes reflètent également cette segmentation et sont souvent répétés dans chaque segment ou métamère.
Pourquoi la métamérisation était-elle si importante? Pour répondre à cette question, il faut d'abord examiner les conditions de vie au moment de l'apparition des Annélides. L'ancêtre des Annélides était un ver plat qui pouvait se déplacer un peu comme la planaire, soit en ondulant ou en rampant à l'aide de cils. Chez les Annélides il y a eu 3 adaptations importantes:
1. L'apparition d'un vrai coelome,
2. la métamérisation, et
3. le développement d'un système circulatoire.
Le coelome permet le mouvement péristaltique par le passage d'une onde de contraction de l'arrière vers l'avant de l'animal. Ce mouvement péristaltique n'est cependant pas très efficace à moins que le coelome ne soit segmenté de manière à ce que chaque segment puisse fonctionner comme un squelette hydrostatique indépendant. Le coelome et la segmentation permirent donc aux Annélides de mieux se déplacer, mais surtout de pouvoir fouir efficacement et ainsi exploiter de nouvelles ressources inaccessibles aux planaires.
Les sédiments marins, au moment de l'apparition des Annélides il y a de cela 600 millions d'années, contenaient déjà d'importantes quantités de matériel organique. Ce matériel, exploité en surface par plusieurs types d'organismes rampants et suspensivores, devenait inaccessible dès qu'il était recouvert par de nouvelles particules. L'apparition des soies permit d'améliorer encore la locomotion des Annélides et d'exploiter cette riche source de nourriture.La métamérisation causa cependant un problème aux premiers Annélides. Chez ces animaux il y a un tube digestif complet qui permet une digestion séquentielle. Certaines régions sont spécialisées pour moudre la nourriture, d'autres pour la sécrétion d'enzymes ou pour l'absorption des nutriments. Cela implique que certains des métamères, isolés les uns des autres, ne sont pas en contact direct avec les éléments nutritifs puisqu'il ne contiennent pas une section du tube digestif où se fait l'absorption de ces éléments. Les métamères antérieurs, s'ils étaient parfaitement isolés des métamères postérieurs, ne pourraient donc pas avoir accès à ces éléments nutritifs. Comment alors avoir accès au matériel nécessaire? L'apparition de la segmentation fut donc accompagnée de l'apparition d'un système circulatoire permettant à chaque métamère de recevoir suffisamment d'éléments nutritifs. Ce système circulatoire sert également à distribuer l'oxygène dans tout le corps. L'apparition de pigments respiratoires est aussi venue augmenter l'efficacité de cette distribution de l'oxygène.
1.DEFINITION
- Métazoaires triploblastiques coelomates.
- Tronc métamérisé : chaque segment ou métamère présente deux sacs coelomiques. Présence de mésentères assurant la suspension des organes vitaux et la formation de cloisons transversales ou dissépiments.
- Complexité croissante de l’organisation générale et différenciation d’organes nouveaux : appareil circulatoire, respiratoire et excréteur.
- Système nerveux concentré et centralisé.
- Larve trocosphère
- Reproduction sexuée, asexuée, grande aptitude à la régénération
- Animaux libres et mobiles : primitivement marins puis dulçaquicoles pour devenir terrestres dans les groupes les plus évolués.
- Trois classes : Polychétes, Oligochétes et Achétes.
2. LES POLYCHETES
Ce sont des vers presque tous marins. Les unes sont Errantes et carnivores, les autres Sédentaires, tubicoles et microphages.
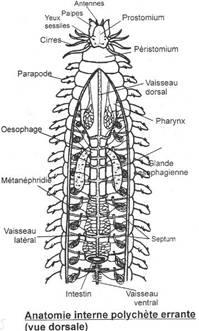
Le genre Nereis réalise leur plan d'organisation. Il mesure environ 10 cm et vit dans la zone littorale. Le corps présente trois régions : la tête, le tronc et le pygidium.
La tête
Elle porte les principaux organes sensoriels et la bouche. Elle est construite en deux parties : la partie antérieure ou Prostonium ou Acron et la partie postérieure ou Péristonium.
Le Prostomium est de forme triangulaire. Il porte deux paires d'yeux simples et une paire d'antennes filiformes à l'extrémité, ventralement et latéralement deux Palpes renflées insérées près de la bouche avec un bouton rétractile terminal. Antennes et palpes sont des organes tactiles et gustatifs. Il ne présente pas de coelome, ce n'est donc pas un vrai segment.
Le Péristomium porte ventralement la bouche et latéralement quatre paires de cirres tentaculaires, expansions latérales à fonction sensorielle. Les segments suivants ne portent que deux paires de cirres, le péristonium résulterait de la fusion de deux métamères. Il porte en outre l'Organe nucal à fonction olfactive.
Le pharynx peut se projeter à l'extérieur sous forme d'une trompe terminée par deux mâchoires cornées. Elle présente dorsalement et latéralement de petits épaississements chitineux appelés Paragnathes. L'augmentation de pression des liquides fait se dévaginer la trompe qui rentre d'elle-même quand la contraction cesse.
Le tronc
Il est composé de nombreux métamères plus petits à l'extrémité postérieure où ils se forment successivement tant que l'animal grandit. Il s'y trouve donc une Zone de prolifération.
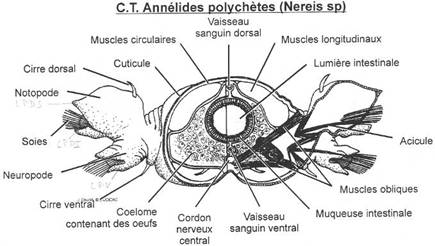
Chaque métamère porte latéralement une paire d'expansions de la paroi du corps, les Parapodes à rôle locomoteur.
Le parapode est composé d'un tégument : un épiderme unistratifié et recouvert d'une cuticule transparente, sous lequel existe un derme avec à l'extérieur des muscles circulaires et longitudinaux.
Les soies que portent les parapodes prennent naissance dans un Bulbe sétigère placé dans un enfoncement épaissi de l'épiderme. Elles sont souvent constituées en deux parties dont la forme varie selon les espèces : la Hampe est la partie basale et la Lame est la partie terminale. Elles tombent périodiquement et sont remplacées par de nouvelles, fournies par le bulbe à partir des Follicules.
Intérieurement deux vésicules coelomiques closes limitées par un endothélium renferment un liquide incolore contenant des cellules amiboïdes qui assurent la nutrition et une partie de l'excrétion tissulaire. Dans le plan sagittal les deux endothélia sont plaqués l'un contre l'autre et forment le Mésentère dorsal et le Mésentère ventral.
Dans chaque cavité s'ouvre un pavillon cilié, le Pavillon néphridien, avec un canal qui débouche dans le métamère suivant.
Ventralement et hors du coelome existe une paire de ganglions nerveux.
Les Dissépiments, rétrécissements diamétraux entre les métamères n'existent pas dans les huit premiers segments qui sont entièrement occupés par la trompe de l'animal.
Les cellules glandulaires à mucus sont nombreuses au niveau des parapodes. Les cellules neuroépithéliales sensitives sont nombreuses au niveau des organes sensoriels.
Le pygidium
Il n'y a pas de coelome. l'ouverture terminale est l'anus.
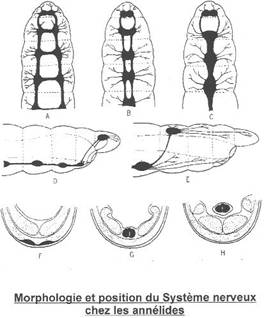
Le système nerveux
Il existe un cerveau à trois régions.
La région Antérieure innerve les palpes, la région moyenne innerve antennes et yeux et la région postérieure innerve l'organe nucal.
Les organes des sens
Il existe des Cellules neuroépithéliales sur cils, antennes et palpes, dont la fonction est gustative et tactile.


L'Organe nucal, une dépression tégumentaire dorsale, est cilié et capte les informations chimiques. Sa fonction est donc olfactive.
Les Yeux dont des invaginations épidermiques dont le fond est garni de cellules visuelles alternant avec cellules visuelles et pigmentaires. Chez les formes plus évoluées il y a en plus un cristallin et une rétine.
Les Statocystes sont les organes de l'équilibration formés de petites vésicules tapissées de cellules neurosensorielles ciliées et contenant des grains de sable ou de calcaire.
Le tube digestif
Il est constitué d'un tube rectiligne de la bouche à l'anus et porte une seule couche de cellules doublée extérieurement d'une couche musculaire. C'est donc un épithélium simple. A l'avant est présente une trompe portant des mâchoires.
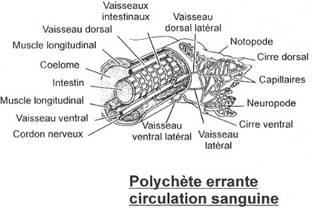
L'appareil circulatoire
Il est entièrement clos. Les vaisseaux sont formés par écartements du mésentère qui forment un vaisseau dorsal et un vaisseau ventral reliés par anastomose au niveau des dissépiments. Le système émet des ramifications dans les parapodes où se fait l'Hématose, les échanges respiratoires et des ramifications au niveau du tube digestif où se fait l'absorption.Le vaisseau dorsal et les anses vasculaires latérales de la région postérieure présente des dilatations de la paroi contractile, ce qui met le segment en mouvement.
L'appareil excréteur
Chaque métamère présente une paire de métanéphridies.
Dans l'embranchement des Annélides le type le plus primitif de néphridies est représenté par les Protonéphridies qui n'existent que chez les larves de polychètes et chez le groupe des archiannélides. C'est un tube à paroi syncipitale renflée en une ampoule coiffée par une cellule qui émet de nombreux flagelles : la Flamme vibratile. Grâce au mouvements de cette flamme le liquide est secrété à l'extérieur par les parois où sont stockés les déchets.
Il existe deux stades dans la complexité des néphridies. Les néphridies à Solennocytes sont constituées par un tube commun et plusieurs cellules monoflagellées à la coiffe. Ces cellules font saillie dans le coelome et sont présentes chez Phyllodoce, Nephthys et Glycera.
Les métanéphridies représentent le stade le plus évolué: elles s'ouvrent dans le coelome par un Pavillon cilié.
A ce système excréteur d'origine ectodermique se superpose souvent le pavillon génital et son canal évacuateur qui sont des évaginations ciliées des vésicules coelomiques ou Coelomoductes. Il existe différents cas selon le rapport entre métanéphridie et canal évacuateur. Chez Phyllodoce la structure est néphridienne à solennocytes, au moment de la maturité apparaît le pavillon cilié génital qui s'ouvre dans le canal néphridien.
Chez presque toutes les formes sédentaires et quelques formes errantes le pavillon génital existe chez le jeune et se superpose à la néphridie.
Chez Capitella le pavillon génital s'ouvre directement à l'extérieur, il forme un conduit sexuel propre et indépendant de la néphridie.
Chez Nereis et Nephthys le pavillon génital se forme mais ne débouche plus à l'extérieur. Il est représenté par l'organe Cilio-phagocytaire.
L'appareil respiratoire
La respiration est cutanée chez Nereis, elle s'effectue par les téguments au niveau des parapodes. Chez quelques polychètes, certaines régions de ce tégument sont différenciées en branchies par des évaginations plus ou moins ramifiées où pénètrent des vaisseaux sanguins.
Chez Arenicola les branchies dérivent du cirre modifié.
Chez Sabella, Serpula, Spirographis les métamères antérieurs portent des branchies ramifiées volumineuses et forment un panache qui peut s'épanouir ou se rétracter rapidement. Chaque segment de ramification porte des divisions secondaires appelées Pinnules.
L'excrétion
Dans le métamère n existe une paire de métanéphridies, pavillons ciliés qui s'ouvrent dans le coelome et suivis d'un canalicule très pelotonné. L'ensemble qui est une vessie débouche dans le métamère n+1.
L'organe cilio-phagocytaire phagocyte et accumule les déchets solides prélevés dans le coelome. Il fonctionne donc comme un rein d'accumulation.
L'appareil génital
Les sexes sont séparés. Il n'existe pas d'organes génitaux bien différenciés.
Les produits génitaux ou cellules germinales se développent aux dépends d'amas de cellules germinales émigrées dans la Splanchnopleure ou épithélium coelomique interne. Il existe une gonade par métamère.
A maturité les produits génitaux tombent dans la cavité générale où ils terminent leur maturation et prennent parfois des couleurs vives. Il n'existe pas de conduits génitaux, l'emission des gamètes se fait par rupture des téguments conduisant à la mort des anneaux chez Nereis. D'autres espèces possèdent des conduits génitaux propres.

L'Epitoquie et son déterminisme
Un certain nombre d'espèces sédentaires présentent à maturité sexuelles des modifications externes et internes importantes. L'animal est alors le siège d'une véritable métamorphose appelée Epitoquie ou Epigamie. Nereis épitoque appartient alors à un genre nouveau, Platynereis.

Figures : appareil circulatoire, ANATOMIE
Déterminisme endocrinien atoque - épitoque
L'épitoquie a pour fonction d'adapter l'animal à une vie pélagique. Il peut alors nager et gagner la surface.
La transformation s'effectue en quelques semaines. Les yeux grossissent, antennes et palpes se rabattent. Dans la région postérieure les métamères se raccourcissent, s'élargissent, se tassent. L'animal devient moins long. Au niveau des parapodes les cirres deviennent dentelées, les soies tombent et sont remplacées par deux larges faisceaux de soies natatoires dont la lame ou article distal est dilaté en palets. Généralement ce sont les régions moyennes et postérieures qui sont modifiées extérieurement. L'animal peut acquérir des couleurs : le mâle de Nereis pelagica est blanc atoque et rouge épitoque, la femelle devient bleutée. Intérieurement il y a histolyse du tube digestif et des muscles longitudinaux. Les muscles des parapodes sont remaniés et permettent la nage.
L'importance des variations épitoques est fonction des espèces. Chez Perinereis cultrifera l'épitoquie est plus importante chez les mâle que chez la femelle. Cette transformation varie également selon les régions. Perinereis cultrifera est épitoque sur les côtes de la Manche et atoque en Méditerrannée. A Arcachon les deux formes coexistent. Nereis diversicolor ne subit
aucune transformation en Manche.L'épitoquie est sous contrôle hormonal. L'expérience a été réalisée par DURCHON sur Perinereis cultrifera et Nereis irrodata. DURCHON a montré qu'en séparant le corps en deux parties à l'aide d'une ligature, le fragment postérieur devient épitoque indépendamment de la maturité sexuelle. Il y a donc eu levée d'une inhibition dans la région postérieure provenant de la région antérieure qui empêchait l'épitoquie de se réaliser et qui cesse à maturité sexuelle.Si on enlève les régions médianes et dorsales du cerveau qui contiennent les cellules neurosécrétrices on obtient une épitoquie prématurée chez les individus jeunes. Si on implante un cerveau chez les individus amputés l'epitoquie n'apparaît pas dans la plupart des cas. Le facteur inhibiteur humoral de l'épitoquie est donc fourni par les cellules neurosécrétrices des ganglions cérébroïdes. De plus le cerveau secrète un facteur inhibiteur de la maturité sexuelle. L'ablation du cerveau entraîne une maturité génitale précoce. Dans le cas normal l'activité inhibitrice du cerveau diminue avec l'âge, ce qui entraîne une apparition de l'épitoquie et de la maturité sexuelle.
Une fois épitoques les Néréides benthiques quittent le fond et se rapprochent de la surface car ils peuvent maintenant nager. La migration en surface se fait en même temps pour tous les individus. C'est un Essaimage qui se produit à dates généralement fixes.Nereis limibata a une migration en relation avec les phases lunaires. Les mâles épitoques montent les premiers au crépuscule peu après la Pleine Lune de juin. Les femelles montent ensuite et secrètent de la Fertilisine, substance induisant une nage désordonnée des mâles conduisant à la rupture de leurs téguments et à la libération des spermatozoïdes. Cette libération induit la rupture des téguments chez les femelles et la libérations des ovocytes. La fécondation se produit dans l'eau de mer.
Un cas célèbre d'essaimage est réalisé par Eunica viridis, dans les régions coralliennes des îles. La partie postérieure sexuée se détache à maturité et monte en surface. Le phénomène se produit avec régularité le troisième jour qui suit le dernier quartier de la lune d'octobre. Les parties supérieures sont très nombreuses et blanchissent la mer. Elles sont recueillies par les indigènes qui s'en nourrissent. C'est le Palolo.
Développement embryonnaire




Le stade Morula, puis Blastula avec blastocoele sont obtenus ensuite. La gastrulation s'effectue par Epibolie. L'archentéron qui dérive des macromères se forme par recouvrement à partir des micromères qui prolifèrent en deux massifs latéraux appelés Plaques somatiques qui se rejoignent et se soudent dans la région médiane. L'anus est en bas, la bouche est en haut. On obtient enfin une larve Trochophore, microscopique, nageuse, planctonique en forme de toupie et à symétrie bilatérale.
L'ectoderme recouvre la larve et se différencie localement en deux couronnes ciliées locomotrices : la Prototroque en avant de la bouche et la Métatroque en arrière. Il existe aussi de petites couronnes ciliées autour de l'anus.
Au sommet de la larve existe une touffe de cils sensoriels, l'Organe syncipital. Sous cette touffe se trouve un ganglion nerveux qui émet quatre paires de cordons nerveux méridiens réunis par un anneau équatorial.
En avant de la larve la bouche et l'ectoderme donne le tube digestif courbé qui se termine par l'anus. Cette larve possède une paire de métanéphridies à solennocytes. De chaque côté du tube digestif au niveau de l'anus existent deux ébauches mésodermiques pleines et non-segmentées. La cavité générale est le blastocoele dans lequel se trouvent quelques cellules de mésenchyme.

La métamorphose de la larve trochophore
Elle est marquée par la formation de métamères s'intercalant entre la région antérieure, le prostomium, et la région postérieure, le pygidium, tous deux dépourvus de coelome.
Un peu en avant de l'anus apparaît une Zone de prolifération où se forment successivement tous les métamères. Les premiers formés sont refoulés vers l'avant par les nouveaux venus. En même temps les ébauches mésodermiques s'allongent dans le blastocoele, se segmentent et chaque massif se creuse d'une cavité à l'origine d'une vésicule coelomique : il y a Schizocoelie. Chaque métamère possède une paire de ces vésicules.
la chaîne nerveuse ventrale se constitue par prolifération de l'ectoderme côté interne. Les néphridies se forment et des initiales germinales se glissent dans la paroi coelomique. Plaque syncipitale, couronnes ciliées et protonéphridies régressnt tandis que se développe antérieurement les palpes et les ganglions cérébroïdes. Finalement l'animal n'étant plus soutenu par les couronnes ciliées tombe au fond, rampe et termine son développement.
Chez certaines espèces de polychètes, à l'éclosion del'oeuf apparaît une larve Métatrochophore qui présente déja les premiers segments.




Biologie des Polychètes
Elles présentent des capacités de régénération importantes. Beaucoup d'espèces peuvent régénérer les palpes, les antennes et des régions plus ou moins importantes de leur corps.
Si on sectionne la région postérieure d'une néréis le morceau postérieur dégénère tandis que la région antérieure régénère une région postérieure. La faculté de régénérer une queue est
limitée à une région postérieure plus ou moins étendue, de même que la faculté de régénérer une région antérieure est limités à la région antérieure de l'animal. Cette faculté diminue quand on s'éloigne de ces régions.
Chez certaines polychètes ces deux régions se touchent et le territoire coupé peut régénérer une tête et une queue. Un fragment trop court dans une région où ces deux territoires se chevauchent peut régénérer une tête à chaque extrémité. Il y a alors Hétéromorphose.
La multiplication asexuée des polychètes présente un pouvoir important en relation avec la régénération directe.
Deux phénomènes d'autotomie suivis de régénération constituent une phase de multiplication asexuées fréquente chez les Néréides et les Syllidés. Lorsqu'elle est indépendante de la reproduction sexuée il y a Schizogénèse. Un ver se coupe en deux : chaque fragment redonne un individu complet. Elle peut devenir le processus normal de reproduction, ayant pu être suivie pendant plus de dix ans chez certaines espèces sans apparition de sexualité.
Le terme ultime de cette multiplication asexuée constitué par la séparation d'un individu en métamères isolés qui redonnent chacun un individu complet a été observé chez Ctenodrilus monostylis. Chez Dodecacena caulleryi la régénération d'une tête peut avoir lieu avant la séparation des fragments. Les individus qui se séparent ensuite sont appelés Zoïdes. Chaque métamère bourgeonne un individu nouveau vers l'avant et un autre vers l'arrière. Les deux individus se détachent puis le métamère donne deux autres individus qui se détachent. Le métamère dégénère enfin.
Souvent la multiplication asexuée interfère avec la reproduction sexuée. Il y a alors Schizogamie.
Chez Eunica viridis seule la région postérieure se détache à maturité sexuelle et elle porte les produits génitaux tandis que la région antérieure régénère une queue. Chez d'autres espèces le fragments postérieur sexué peut régénérer une tête : il y a donc superposition de la multiplication asexuée.
CLassification
- Les Errantes
Le corps est allongé et constitué de très nombreux segments. La tête est généralement bien individualisée avec des organes sensoriels. Ils vivent dans le sable, sous les pierres, dans les fentes de rochers. Ils sont souvent carnassiers.
Le genre Nereis vit sur dans les crampons de laminaires, tel Nereis pelagica. Il est siège de l'épitoquie.
Le genre Nephthys vit dans le sable. Les mâchoires cornées sont toujours internes. La trompe ne présente plus de paragnathes mais des papilles molles.
Le genre Aphrodite, taupes de mer vit dans les rochers. Son corps est trapu et aplati. Les cirres dorsaux sont aplatis, transformés en lamelles et se couvrent comme les tuiles d'un toit.
Le genre Phyllodoce vit sous les pierres et dans les rochers. Les cirres dorsaux et ventraux sont en forme de bouclier.
- Les Sédentaires
Les Tubicoles vivent le plus souvent à l'intérieur d'un tube. La tête est petite et peu distincte. Ils possèdent des branchies dans une région branchiale antérieure et sont généralement microphages. Leur tube peut être composé de différentes substances.
Chez Sabella et Spirographis, il est constutué de mucus imprégné de vase. Les branchies forment un panache pouvant se rétracter à l'avant.
Chez Serpula le tube est Arénacé car constitué de mucus imprégné de grains de sable.
Chez serpula, Spirorbis, Pomatoceros, le tube est calcaire. L'animal ferme l'entrée grâce à un Opercule.
Les Fouisseurs vivent dans un sable vaseux où ils creusent un tube en U, comme Arenicola marina. Certains fouisseurs perforent les roches, comme Polydora ciliata. Les parties antérieures et postérieures émergent.
3. LES OLIGOCHETES
Ces animaux sont surtout terrestres ou dulçaquicoles. Ils dérivent des Polychètes par simplifications anatomiques concernant les parapodes.
Le prostomium est réduit. Il ne porte aucun organe de sens externe. En arrière de la trompe se trouve le péristomium qui ne porte pas de soies. Sur les autres segments le nombre de soies est réduit à quatre paires par métamère. Elles sont petites et implantées dans le tégument. Il n'existe aucun parapode.
Les Oligochètes sont hermaphrodites, leurs organes génitaux sont localisés dans un petit nombre de segments, les conduits génitaux sont indépendants des néphridies.
Leur plan d'organisation est réalisé simplement par Lombricus herculeus ou Ver de Terre.
Morphologie
L'animal est blanc-rosé, plus coloré sur sa face dorsale et composé de 108 à 180 anneaux. La région antérieure effilée porte la bouche, la région postérieure aplatie dorso-ventralement porte l'anus.

Il existe quatre paires de soies ressemblantes à de petits crochets par anneau. C'est l'équivalent en réduction des rames de parapodes.
Depuis le 32e segment jusqu'au 37e les anneaux sont dorsalement plus renflés et forment le Clitellum surtout visible au moment de l'accouplement.
Au niveau de chaque segment, sauf dans le cas des trois premiers, se trouve une paire d'orifices excréteurs.
Les orifices génitaux entourés par deux lèvres épaisses sont au nombre de deux pour les voies sexuelles mäles et situés face ventrale du 15e anneau. Au moment de la reproduction ces deux orifices sont prolongés par un sillon spermatique qui va jusqu'au clitellum.
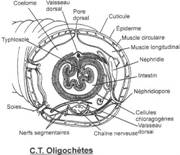
Les téguments
Ils présentent un épithélium simple recouvert d'une cuticule où sont présentes des cellules à mucus favorisant la reptation et des cellules neuroépithéliales sensorielles. Il existe des Pores coelomiques dorsaux assurant les communications entre les vésicules coelomiques et la surface épidermique.
Sous l'epiderme, plus profondément, il existe deux couches musculaires de muscles circulaires et muscles longitudinaux.
Les métamères
Il y en a autant que d'anneaux visibles. Dans chacun les vésicules coelomiques ont fusionné grâce à la disparition du mésentère dorsal et ventral. Les dissépiments subsistent.
Le système nerveux
Il ressemble à celui des Polychètes et est composé par une paire de ganglions cérébroïdes supra-oesophagiens, un collier périoesophagien, un ganglion sous-oesophagien et une chaîne nerveuse ventrale portant une paire de ganglions par métamère. Le cerveau est relégué dans le troisième métamère.
Les animaux présentent un Phototactisme négatif dont le centre se situe au niveau de l'acron. Ils sont en général aveugles mais certaines espèces présentent des yeux rudimentaires.

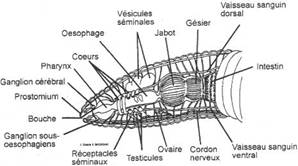
Le tube digestif
Il est rectiligne. Après la bouche s'ouvre un pharynx musculeux dont la dilatation provoque une dépression pneumatique qui facilite l'aspiration des particules de terre et de feuilles dont se nourrit l'animal.
Ensuite il y a un oesophage animé de mouvements péristaltiques dans lequel débouchent trois paires de Glandes calcarifères ou Glandes de MORREN secrétant des concrétions de calcite. Leur rôle est important car elles sont bien vascularisées mais elles sont peu connues et constitueraient une réserve de calcium ou seraient chargées d'effectuer l'évacuation du gaz carbonique sous forme de carbonates. Le ver de terre neutralise ainsi l'humidité de l'humus.
En arrière de l'oesophage sont présents un Jabot et un Gésier, deux parties dilatées. Ce dernier possède une musculature importante, des crètes obliques recouvertes d'une cuticule épaisse et assure le malaxage de la terre et des feuilles ingérées. Son rôle est également filtrant, les particules non-assimilables sont rejetées.
L'intestin présente un repli dorsal en forme de gouttière, le Typhosolis, dont le rôle est d'augmenter la surface de contact avec les aliments. Le ver utilise les particules organiques de ses aliments et rejette les résidus sous forme de tortillons très volumineux car ses aliments n'ont que peu de valeur nutritive.
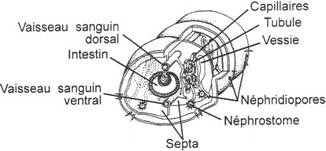
L'appareil excréteur
On trouve une paire de néphridies par métamères sauf dans les trois premiers segments. Le pavillon cilié ou Néphrostome s'ouvre dans un métamère n et un canalicule très fin et contourné s'ouvre dans le métamère n+1. Dans la splanchnopleure qui entoure l'intestin sont présentes des cellules brunâtres accumulant les déchets (guanine) et nommées Cellules chloragogènes. Leur extrémité apicale tombe dans le coelome et les déchets sont alors phagocytés par les cellules amiboïdes qui s'amassent surtout dans la région postérieure formant des nodules brunâtres à structure syncitiale rappellant un plasmode, élimination par les néphridies.
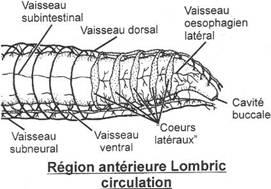
L'appareil circulatoire
Il est clos. Le sang est rouge car il contient une chromoprotéine voisine de l'hémoglobine et il contient aussi des globules blancs. Il n'existe pas d'hématies.On trouve un vaisseau dorsal portant des renflements contractiles dans chaque segment et un vaisseau ventral situé sous le tube digestif. Dans la région antérieure ces vaisseaux sont réunis par cinq anneaux transverses contractiles nommées Coeurs latéraux plus gros et contractiles. Il existe également un vaisseau sous-neural.


L'appareil respiratoire
La respiration s'effectue au niveau de la peau dont le tégument doit toujours être humide sinon la mort est rapide.
Certaines espèces limicoles ou vivant dans la vase respirent par des branchies filamenteuse. Le respiration est normale tant que la pression d'oxygène n'est pas trop basse, sinon elle diminue. Les Oligochètes peuvent supporter assez longtemps une privation totale en oxygène : ce sont donc des organismes Aérobies facultatifs qui oxydent les produits de l'anaérobiose quand l'oxygène revient.
L'appareil génital
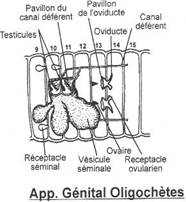

Les animaux sont hermaphrodites et leurs organes génitaux sont situés dans la région antérieure avec les ovaires en arrière des testicules.
L'appareil mâle
Deux pares de testicules sont situés dans les 10e et 11e métamères. Il existe trois paires de vésicules séminales volumineuses qui sont des excroissances du coelome. Les deux premières dépendent du 10e segment et la troisième du 11e. Dans ces vésicules s'effectuerait la spermatogénèse : les gonilles y migrent. Si expérimentalement on enlève les vésicules séminales la spermatogénèse se produit dans le coelome. Deux paires de pavillons ciliés captent les spermatozoïdes qui tombent dans des canaux déférents se jetant dans un spermiducte, un à droite et un à gauche et qui s'ouvrent dans le 15e segment.
L'appareil femelle
Une paire d'ovaires est située dans le 13e métamère contre le dissépiment antérieur. Les ovules tombent dans le coelome et sont recueillis par les pavillons ciliés prolongés chacun par un court oviducte qui s'ouvre dans le 14e segment.
En plus il existe deux réceptacles séminaux en forme de poches dans les 9e et 10e métamères. C'est là que s'accumule le sperme émis par le conjoint.
L'accouplement
L'accouplement est réciproque entre deux vers quand les organes mâles sont mûrs : l'hermaphrodisme est qualifié de Protérandrique. Le clitellum est alors bien visible, saillant, épais et glandulaire. Les orifices génitaux mâles ont leurs lèvres bien dévoloppées, les deux gouttières spermatiques ventrales apparaissent nettement jusqu'au niveau du clitellum. La castration ne modifie pas le cycle d'évolution de ces caractères sexuels secondaires dont le déterminisme est neuroendocrine.
L'ablation des ganglions cérébroïdes produit l'involution des caractères sexuels secondaires. Après deux semaies l'animal prend un aspect de ver en repos sexuel. De quatre à cinq semaines après il peut à nouveau se reproduire, les ganglions cérébroïdes ont régénéré.
L'accouplement s'effectue ventre-à-ventre et tête-bêche, le clitellum de l'un en face des réceptacles séminaux de l'autre. Chaque clitellum secrète un anneau de mucus qui durcit et unit étroitement les conjoints. Le sperme émis par l'un suit les crêtes séminales et s'accumule dans les réceptacles séminaux de l'autre et récoproquement. Il existe une partie commune où les deux spermes se rencontrent mais cette partie est fermée, interdisant tout mélange : les quatre gouttières sont indépendantes. Ensuite les deux vers se séparent.
Les ovaires arrivent à maturité, chaque ver secrète à nouveau un manchon au niveau de son clitellum et l'animal en sort à reculons. Le manchon passe devant les orifices mâles et femelle. A ce moment l'animal pond ses ovules puis au niveau des réceptacles séminaux remplis de spermatozoïdes la fécondation se produit dans le manchon. Finalement le manchon est libéré et devient un Cocon de ponte. Il se ferme aux deux extrémités et s'affaisse.
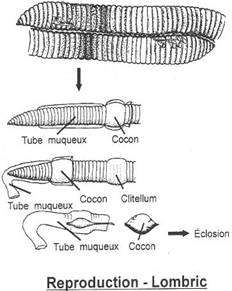
Le développement
Les oeufs riches en vitellus se développent directement sans passer par un stade larvaire libre. La segmentation est de type spirale mais moins régulière que chez les Polychètes. Du cocon sortent de minuscules lombrics
Biologie
Les limicoles présentent des capacités de régénération importantes, moins marquées chez les terricoles.
Si on coupe un ver en deux on obtient deux individus complets.
La multiplication asexuée est très fréquente chez les limicoles et en général avancée quand l'individu se fragmente en zoïdes qui peuvent rester longtemps attachés avant de se séparer. Chez certaines espèces elle constitue le seul mode de reproduction. Chez Tubifex on observe des bourgeonnements latéraux qui produisent de nouveaux individus.
Lorsqu'il se déplace, il s'allonge et se raccourcit grâce à ses muscles. Il peut s'accrocher au sol à l'aide de ses soies.
Lorsqu'il se nourrit de débris végétaux ou de matière organique de la terre dont il ingurgite de grosses quantités, il consomme plusieurs fois son poids en une journée. Il peut se nourrir de viande. Il distingue une feuille d'oignon d'une feuille de chou grâce à ses cellules olfactives.
Il creuse des galeries dont il ne sort que la nuit car il a un phototactisme négatif. Il a un rôle important dans l'ameublissement de la terre dont il ramène les parties profondes en surface, laissant l'air circuler plus facilement.
Classification
On distingue deux grands groupes.
Les limicoles
Ils vivent dans la vase, les mares, les ruisseaux et les étangs. Quelques-uns sont marins.
Ils sont de petite taille, grêles, leurs soies sont plus nombreuses que celles des terricoles, comme chez Nais.
Tubifex présente une teinte rouge et vit la tête implantée dans la vase. Ils peuvent être indicateurs biologique d'une pollution car ils ne vivent que dans une pression d'oxygène faible.
Les terricoles
Ils sont plus grands, atteignent 2 à 3m. En France 80 cm pour 1 à 1,5 cm de long.
Les vers de terre épigés
Ils vivent à la surface du sol, au niveau de la litière et dans les matières organiques en décomposition. On les trouvera également dans les excréments des grands herbivores ou dans le bois humide en cours de décomposition. Peu protégés ils subissent une forte prédation qu'ils compensent par une fertilité élevée 42 à 106 cocons par adulte et par an. Quand la nourriture est abondante et les conditions climatiques favorables, ils peuvent se multiplier très rapidement. Leurs cocons, qui résistent à la sécheresse, assurent la survie de l'espèce. Les vers de terre épigés jouent un rôle important dans le recyclage de la matière organique. On les utilise parfois de façon industrielle pour produire du « lombricompost » et pour traiter les ordures ménagères. Certaines espèces sont élevées pour servir d'appâts pour la pêche.
Les vers de terre endogés :
Ils représentent 20 à 50 % de la biomasse des terres fertiles et vivent en permanence dans le sol où ils creusent des galeries horizontales. Ils ne sont pas pigmentés. Ils ont une fécondité moyenne 8 à 27 cocons par adulte et par an. Ils se nourrissent de terre plus ou moins riche en matière organique. En période de sécheresse ils tombent en léthargie et on les trouve enroulés sur eux-mêmes. Les vers endogés présentent des modes de vie assez différenciés. Certains sont filiformes et s'installent le long des racines, d'autres forment des pelotes dans les couches profondes du sol, à proximité des drains, et filtrent l'eau dont ils séparent les particules organiques. On en connaît aussi qui sont prédateurs d'autres vers de terre.
Les vers de terre anéciques
Ce sont eux que l'on trouve lorsqu'on retourne la terre du jardin. S'il n'y a pas de vers de terre dans votre jardin ce n'est pas bon signe.... Ils vivent dans des galeries verticales et viennent « faire leurs provisions » à la surface du sol tout en restant prudemment accrochés par la queue à l'entrée de leur terrier. Les feuilles et les débris organiques qu'ils peuvent entraîner dans leurs galeries sont ingurgités avec de la terre. Les excréments sont déposés à la surface du sol sous forme de tortillons appelés aussi turricules. Des trois groupes ce sont eux qui ont la fécondité la plus réduite 3 à 13 cocons par adulte et par an. En Europe tempérée les vers anéciques représentent 80 % de la masse totale des lombrics. En période estivale ils tombent en léthargie. Le plus grand vers anécique de la faune française dépasse 1 mètre de long.
SCHEMAS COMPLEMENTAIRES



4. LES ACHETES OU HIRUDINEES

Ils sont également connus sous le nom d'Hirudinés.
C'est une classe d’ectoparasites, souvent aquatiques.
Ils ne portent ni soies ni parapodes mais une ou deux ventouses fixatrices.
Leur nombre de métamères est constant.
Les cavités coelomiques ont disparu et sont comblées par un tissu mésenchymateux qui provient de la différenciation des parois coelomiques dissociées qui ne sont représentées que par deux sinus longitudinaux : un sinus ventral et un dorsal.
Ces animaux sont hermaphrodites, le clitellum est réduit au niveau des segments génitaux.
Ils sont généralement aquatiques, surtout en eau douce. Il en existe quelques marins. La plupart sont ectoparasites de vertébrés. Quelques-uns sont terrestres et vivent en milieu humide.
Pas de croissance pygidiale
Le plan d'organisation des Achètes est représenté simplement par Hirudo medicinalis ou sangsue médicinale. Elle a été exploitée pour faire des saignements et a tendance à disparaître.
Morphologie
L'animal vit dans les mares et les étangs. Il mesure de 10 à 12 cm de long, 1,5 cm de large et est aplati dorso-ventralement. Son poids est de 2 à 3g. La face dorsale est foncée de couleur gris-vert, la face ventrale est vert-clair et porte deux bandes longitudinales jaunes-orangées.
L'avant porte la bouche entourée par une ventouse et l'arrière se termine par une ventouse subcirculaire plus grande.
On observe des sillons transversaux nombreux et des anneaux rapprochés mais ils ne correspondent pas à de vrais métamères. Il existe 101 anneaux visibles pour 33 métamères.
Les premier et second métamères correspondent chacun à un anneau. Le troisième métamère correspond à deux anneaux. Les quatrièmes, cinquièmes et sixièmes métamères correspondent à trois anneaux. Enfin à partir du septième, chaque métamère est représenté par cinq anneaux.
La face dorsale comporte cinq paires d'yeux situés sur le bord du premier anneau de chacun des cinq premiers métamères.
Le premier anneau de chaque métamère porte huit taches sensorielles pigmentées constituées de cellules neuroépithéliales.
Sur la face ventrale l'orifice génital mâle est présent sur le 10e segment. Un pénis filamenteux et blanchâtre peut faire saillie. L'orifice femelle est porté sur le 11e segment.
Dans la région moyenne existe une paire de néphridies par métamère. En tout il y a 17 paires de néphridies avec une paire d'orifices ventraux sur le premier anneau de chaque métamère.

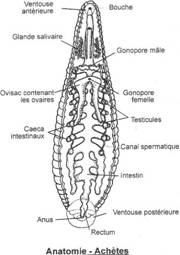
Les téguments
Il présente de nombreuses glandes à mucus, possède une assise de muscles circulaires extérieurs et longitudinaux à l'intérieur. Il y en plus des muscles dorso-ventraux et obliques.
L’épiderme est mono-stratifié, recouvert par une cuticule plus ou moins épaisse, selon la région du corps et selon l’espèce. La cuticule s’interrompt au niveau d’un petit orifice correspondant aux glandes cutanées.
Le derme est formé d’un tissu conjonctif. Sous le derme, on trouve une musculature très développée, présente sous trois formes : circulaire, longitudinale et dorso-ventrale.
Le système nerveux
Il est représenté par une paire de ganglions cérébroïdes, un collier périoesophagien, une masse nerveuse sous-oesophagienne correspondant à cinq paires de ganglions fusionnés, une chaîne nerveuse ventrale avec une paire de ganglions par métamère et un gros ganglion terminal, coalescence des sept dernières paires.
De A à C, on observe une fusion des ganglions puis des cordons nerveux.
En D et E, on voit les ganglions cérébroïdes qui reculent en arrière du prostomium, vers M3. Les ganglions sous-œsophagiens sont repoussés dans M4. On a aussi un développement des prolongements des chaînes nerveuses vers l’avant pour les organes sensoriels et les muscles.
De F à H, on observe un enfoncement de la chaîne nerveuse à l’intérieur de l’animal. Primitivement (en F), la chaîne nerveuse est sous-épidermique ; elle s’enfonce ensuite (en G) dans les muscles longitudinaux (sous-dermiques) et finie (en H) dans la chaîne de la cavité cœlomique.
Le tube digestif
Il est rectiligne. La bouche est au fond de la ventouse buccale et présente deux lèvres dorsales let une lèvre ventrale. Elle st entourée par trois mâchoires à 120° l'une de l'autre. Ce sont des lames chitineuses semi-circulaires portant de nombreux denticules de calcite sur le bord arrondi. Quand les dents coupent la peau la blessure est typique en Y.
Entre les denticules débouchent des canaux reliés au glandes salivaires qui produisent l'Hirudine, une substance anticoagulante. Toutes les sangsues n'ont pas cette mâchoire, certaines ont une trompe exsertile.
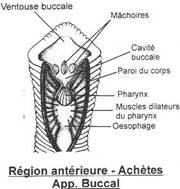

A la suite de la cavité buccale, un court oesophage puis un estomac composé de onze paires de caecums accumule le sang ingéré. Chaque caecum est pourvu d'un sphincter.
L'estomac débouche dans un intestin rectiligne. L'anus débouche sur la face dorsale de la ventouse postérieure.
Le coelome et les cavités coelomiques
Chez la larve chaque métamère possède une paire de sacs coelomiques avec mésentère et dissépiment. Chez l'adulte tout est comblé par du tissu mésenchymateux qui provient des parois coelomiques. Au terme de ce processus le coelome n'est plus représenté que par deux sinus principaux, un ventral et un dorsal, qui entourent la chaîne nerveuse. Dans les sinus se trouve un liquide plasmatique rouge. Ils sont réunis par de fins sinus transverses anastomosés. De petits sinus coelomiques entourent également testicules et ovaires.
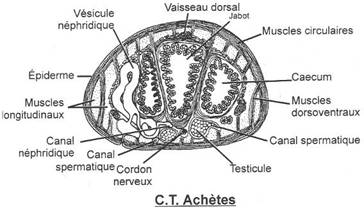
L'appareil circulatoire
Il est constitué par deux vaisseaux latéraux contractiles réunis par 17 paires de vaisseaux transverses dorsaux.
Le sang est rouge : il contient une chromoprotéine proche de l'hémoglobine.
Les vaisseaux latéraux émettent des ramifications vers les néphridies, les organes génitaux, le tube digestif et le tégument.
L'interprêtation des appareils circulatoires et coelomiques est difficile. Les vaisseaux dorsaux et ventraux des Polychètes et Oligochètes ont disparu et sont remplacés par les deux sinus
dorsaux et ventraux et le système circulatoire latéral est en fait indépendant. Or il existe des communications entre vaisseau ventral et vaisseau latéral.
L'appareil respiratoire
Il n'est pas différencié, les échanges se faisant au niveau des téguments.
L'appareil excréteur
17 paires de néphridies sont présentes dans les segments 6 à 22. Les pavillons ciliés au niveau des néphrostomes ont disparu car il n'y a plus de vésicules coelomiques.
Le néphrostome est suivi d'une masse blanchâtre, la Glande en fer à cheval constituée de cellules excrétrices entourant le canal excréteur très contourné, fermé au départ et qui se termine par une vessie dilatée qui s'ouvre par un pore excréteur dans le métamère n+1.
Dans les métamères testiculaires le néphrostome est en relation avec le sinus péritesticulaire qui est un reste de coelome et contient un organe cilio-phagocytaire.
Il existe des cellules capables d'accumuler les produits de déchets. L'ensemble de ces cellules forme le Tissu bothryoïde situé au niveau testiculaire.
L'appareil génital
L'appareil mâle
Il comprend neuf paires de testicules dans les segments de 12 à 20. Deux rangées parallèles de chaque côté de la chaîne au niveau ventral. Chaque testicule est entouré d'un reste de coelome.
De chaque testicule part un court canal déférent transversal se jetant dans un spermiducte longitudinal qui se dirige vers l'avant et se pelotonne au niveau du dixième segment en une masse irrégulière, l'Epididyme, un de chaque côté. Au sortir des épididymes les canaux spermiductes confluent et donnent un canal éjaculateur impair dont la paroi forme un pénis dévaginable et sa base est entourée d'une glande annexe, la Prostate qui élabore la coque du Spermatophore.
L'appareil femelle
Il est contenu en entier dans le 11e segment. Deux ovaires qui sont entourés d'un reste de coelome émettent chacun un petit oviducte. Les deux se rejoignent en un utérus entouré par la Glande de l'albumine, puis un vagin qui s'ouvre par l'orifice génital femelle sur le 11e segment.
Il y a accouplement réciproque tête-bêche lorsque l'appareil mâle est à maturité mais pas d'introduction du pénis dans le vagin. Le sperme est émis par petits paquets, les spermatophores. Ils sont implantés dans la peau, la fécondation est dite Traumatique.

La fécondation
L'enveloppe du spermatophore est digérée par les diastases issues de l'épiderme et des tissus sous-jacents. Les spermatozoïdes non-utilisés sont digérés par des amibocytes dans les mailles du tissu bothryoïde près des testicules. le clitellum est limité aux métamères 10 et 11. Il secrète un manchon de mucus qui durcit et sert à recevoir les ovules fécondés. comme chez le Ver de terre la manchon se ferme aux deux extrémités en une sorte de cocon rempli d'une secrétion albumineuse dans laquelle commence le développement indirect des oeufs.
La segmentation est spirale, mais modifiée. Il n'existe pas de multiplication asexuée, chose à mettre en relation avec l'absence de croissance métamérique post-embryonnaire, ni régénération mais les blessures cicatrisent.
Biologie
Elles avancent dans l'eau en nageant, ondulent latéralement ou peuvent arpenter une surface.
Elles peuvent absorber de cinq à six fois leur poids en sang.
Classification
On distingue trois groupes.
Les Acanthobdèles forment un groupe de transition avec les Oligochètes. Ils présentent une seule ventouse postérieure. Les segments deux à six portent quatre doubles rangées de soies. Les cavités coelomiques sont fereconnaissables. Ce sont des parasites de Salmonidés.
Les Rhyncobdèles possèdent une trompe exsertile. Ils sont souvent parasites d'invertébrés d'eau douce, comme Piscicola geometra ou Clepsine qui porte trois paires d'yeux.
Les Arhyncobdèles ne possèdent pas de trompe. Les Pharungobdèles possèdent un pharynx puissant et musculeux, pas de mâchoire. Les Gnathobdèles possède des mâchoires, comme Hirudo medicinalis et Haemoptis sanguisuga ou Sangsue de cheval.
SCHEMA COMPLEMENTAIRE


Date de dernière mise à jour : 12/01/2019
Ajouter un commentaire