
Spongiaires/Cnidaires
Parmi les métazoaires nous commençons avec les diploblastiques : spongiaires, Cnidaires et Cténaires
1.Métazoaires primitifs /.SPONGIAIRES

1 Position systématique
L'embranchement des Spongiaires est actuellement représenté par 3 classes :
- Les Démosponges
- Les Hexactinellides
- Les Éponges calcaires
Les Spongiaires ont été considérés, par le passé, comme un stade intermédiaire entre les Protozoaires et les Métazoaires ( Parazoaires ). Cette hypothèse semble infirmée par les indications collectées grâce à la phylogénie moléculaire, qui les a fermement inclus dans le groupe des métazoaires.
A nouveau cette conception semble changer car il dériveraient de protozoaires particuliers appelés choanoflagellés qui ressemblent étrangement au choanocytes de l'assise cellulaire interne de l'organisme (voir paragraphe suivant) .
2 Définition
Métazoaires diploblastiques
Métazoaires archaïques ne présentant pas de symétrie et dont le corps est constitué de deux feuillets. Organismes diploblastiques. actuellement on hésite à les dire " diploblastiques" car le rassemblement des cellules est fait sans membrane basale, ce ne sont pas des tissus au sens strict du terme;
La cavité ou gastrique ou archentéron est tapissée par l’endoblaste constitué de choanocytes,cellules à collerette.
Les deux feuillets embryonnaires sont séparés par une couche gélatineuse, la mésoglée, envahie secondairement par différents types de cellules.
Le système nerveux est diffus et rudimentaire.
La fécondation indirecte.
Pas d’organes définis mais plusieurs sortes de cellules.
Animaux aquatiques essentiellement marins
La paroi du corps est perforée et parcourue par de nombreux canaux dits inhalants.
Ce sont des organismes filtreurs,l’eau s’introduit par les pores inhalants, parcourt les canaux inhalants et ressort par l’oscule.Cette eau est filtrée grâce au passage à proximité des choanocytes.
Animaux sessiles dont les seuls mouvements sont des contractions et l’occlusion des pores et oscules.
Squelette formé de spicules calcaires, siliceuses et organiques.
Reproduction sexuée, asexuée et faculté de régénération à partir de fragments, voire de cellules isolées.
Ces organismes peuvent se présenter sous trois formes différentes que l’on apelle stades :
- Stade Ascon
- Stade Sycon
- Stade Leucon.
Ces stades ne sont pas nécessairement présents chez toutes les espèces, mais un des trois stades peut être caractéristique d’une espèce donnée. Ces stades correspondent à une organisation de plus en plus ramifiée en cavités gastrales secondaires et finalement en corbeilles vibratiles.

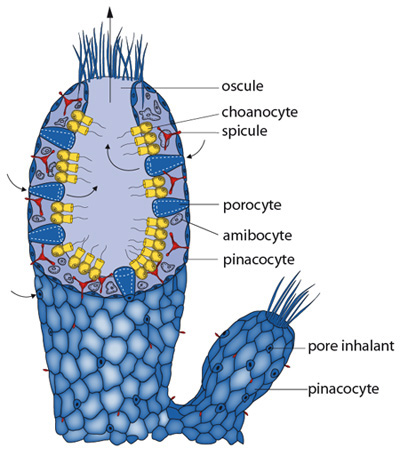
3 Morphologie-anatomie
Description faite sur le stade Ascon. Coupe longitudinale de paroi didermique
a) Feuillet externe ou ectoblaste :
Epithélium formé de cellules pavimenteuses appelées pinacocytes , entre ces cellules s’ouvrent les pores inhalants .
b) Feuillet interne ou endoblaste
Feuillet tapissant la cavité gastrale ( archentéron) , formé de choanocytes ( cellules à collerette). Les mouvements des flagelles apportent à l’éponge l’oxygène et les particules alimentaires ( bactéries, algues unicellulaires).
c) Mésoglée
Plusieurs variétés ont migré . Aucune cohésion entre elles , donc pas d’organes, on distingue :
- Les porocytes, cellules drains tubulaires
- Les amoebocytes, cellules arrondies donnant naissance au gonocytes, aux phagocytes etc…
- Les cellules nerveuses, cellules fixes généralement filiformes, elles réalisent un relais entre pinacocytes et choanocytes.
- Les scléroblastes, d’origine ectodermique, ils élaborent les spicules. Formation d’un squelette interne.

4 Classification
Celle-ci se base sur la nature des éléments squelettiques, on peut diviser cet embranchement en trois classes :
4.1 Les éponges calcaires
La classification des éponges calcaires est basée sur la distribution des choanocytes qui peuvent tapisser entièrement la cavité gastrale : éponges calcaires homocoeles,ou se répartir dans des diverticules de cette cavité : éponges calcaires hétérocoeles.
4.1.1 éponges calcaire homocoeles
La cavité axiale est à le fois cavité gastrale et corbeille vibratile, il peut y avoir une légère complication. On a un ou peu d’oscules. Ce sont des formes typique du stade Ascon
Ex : Olynthus sp, Asceta sp, Leucosolenia sp, Clathrina…
4.1.2 éponges calcaire hétérocoeles
Le stade Ascon est transitoire ou escamoté, développement caractérisé par une complexité croissante.

On passe généralement au stade Sycon avec cavités gastrales secondaires, la cavité axiale s’appelle atrium ( voir schéma).
Finalement, stade Leucon avec fort développement du cortex.
Ex : Grantia sp, Sycon sp, Leucandra sp.
Organismes communs des plateaux continentaux.
4.2 Les Desmosponges
Spicules siliceux et fibres de spongine ( scléroprotéine riche en ponts disulfure et en tyrosine halogénée) reliant les éléments squelettiques.
Cette classe reprend des animaux intéressant soit sur le plan économique, soit sur le plan biologique :
- Cliona qui perfore les coquilles de Mollusques lamellibranches ( Huîtres ).
- Spongia : éponge de toilette.
- Certaines vivent dans les eaux douces.
4.3 Les Hexatinellides
Spicules siliceux
5 Reproduction et régénération
5.1 La reproduction sexuée
Dans la mésoglée, les amoebocytes donnent des gonocytes qui engendrent des ovogonies et des spermatogonies. Pas encore de véritables gonades.
Les éponges sont gonochoriques ou hermaphrodites.
5.1.1 La gamétogénèse
- Spermatogénèse : normale donnant naissance à des spermatozoïdes flagellés qui seront libérés dans le milieu extérieur.
- Ovogénèse : l’ovocyte I s’associe à deux cellules , cellule satellite et cellule nouricière.
5.1.2 La fécondation
Celle - ci est indirecte, elle s’opère après que le spermatozoïde ait fusionné avec un choanocyte, ce qui donne une cellule « charriante » qui
transporte son noyau et celui du spermatozoïde. Le noyau du spermatozoïde associé à une vacuole forme le spermiokyste.
Lorsque la cellule charriante entre en contact avec un ovocyte, elle émet un prolongement qui pénètre dans cet ovocyte et forme un « tunnel » par lequel le spermiokyste pourra migrer dans le cytoplasme ovocytaire. Il y a ensuite caryogamie et formation du zygote.

Les spermatozoïdes proviendraient de choanocytes qui migrent dans la mésoglée, s'y dédifférencient en perdant leur flagelle et leur collerette, puis se différencient en spermatogonies. Chez d'autres espèces, les spermatogonies se différencient directement à partir d'archéocytes.
Les spermatogonies se multiplient puis subissent la méiose et la spermiogenèse. Les spermatozoïdes quittent l'éponge-père avec le courant exhalant.
Les ovules proviennent d’archéocytes, qui se différencient et accumulent des réserves. Les ovules restent en place dans la mésoglée.
La fécondation est très particulière: un spermatozoïde entre avec le courant d'eau inhalant; il est capté par un choanocyte qui fait office de cellule transporteuse au sein de la mésoglée. Arrivé près d'un ovule, le choanocyte lui transmet le spermatozoïde.
Le développement du zygote se produit à l'intérieur de l'éponge. Il existe plusieurs types d’embryons, notamment :
Parenchymula
Embryon couvert d'une assise externe de cellules flagellées, et qui contient à l'intérieur la plupart des types cellulaires de l'adulte, sauf les choanocytes.
Amphiblastula
Embryon creux dont un des hémisphères est composé de petites cellules flagellées, et l'autre de grosses cellules non flagellées.
Ces embryons quittent leur éponge-mère et mènent une brève existence libre. Puis, après leur fixation sur un support, ils subissent une réorganisation.
Chez la parenchymula, les cellules externes flagellées perdent leur flagelle, se déplacent vers l'intérieur et s'y différencient en choanocytes.
Chez l'amphiblastula, les petites cellules flagellées s'invaginent et les grosses cellules finissent par les entourer.
Ces modes de ségrégation des feuillets sont assez différents de ceux rencontrés chez les autres Métazoaires.
5.1.3 Le développement embryonnaire
Les éponges sont vivipare, l’embryon se développe dans la mésoglée. A la fin du développement, une larve traversera la couche des choanocytes et gagnera , via l’atrium, le milieu extérieur ( amphiblastula).
Certaines espèces d’éponges ( Desmosponges ) émettent des larves plus évoluées, véritables pupes appelées parenchymula.


5.1.4 Le développement larvaire
La jeune larve nage librement grâce à des flagelles, elle finira par se fixer. Une fois fixée, le blastopore se ferme et au pôle opposé, s’ouvrira l’oscule.
5.2 La reproduction asexuée
Chez les métazoaires, la reproduction asexuée procède selon ; le bourgeonnement et la schizogénèse.
Chez les Spongiaires , on rencontre le bourgeonnement.
Les cellules qui donneront le bourgeon par prolifération peuvent appartenir à des tissus différenciés, mais elles devront subir , au préalable, une préparation, la blastogénèse : c’est le pendant de la gamétogénèse de la reproduction sexuée.
La blastogénèse conduira à la formation de blastozoïtes
Chez les éponges, on rencontrera le bourgeonnement actif ou externe, dans lequel les blastozoïtes entrent immédiatement en prolifération et un individu se sépare de la souche mère.
On rencontrera également le bourgeonnement de protection ou interne , par lequel le précurseur d’une forme de résistance est générée à l’intérieur de l’organisme mère, par exemple des spongilles ( schéma ) qui vont se libérer et se fixer à l’extérieur. Ces spongilles contiennent des gemmules formés dans leur mésoglée. Ces gemmules sont un amas de cellules indifférenciées limité par une coque percée d’un micropyle. Les spongilles sont libérées à la mauvaise saison par l’organisme qui meurt. Au printemps suivant les gemmulles germent et donnent naissance à de nouveaux individus.

 Détail de la gemnule
Détail de la gemnule
5.3 La régénération
- Grandes facultés de régénération.
- Les cellules isolées sont capable de régénérer totalement un individu. Il s’agit donc d’un type de régénération à potentialités totales.
6 Biologie
L’eau pénètre par les pores et ressort par l’oscule.
L’eau absorbée contient des substances dissoutes et de très fines particules
Les choanocytes captent les principes nutritifs qui seront transmis aux différentes cellules par les amoebocytes.
7 Écologie
Les éponges habitent généralement les zones littorales et sublittorales où la nourriture (phytoplancton,bactéries) est abondante. Elles filtrent d'énormes quantités d'eau et contribuent à réduire la turbidité de l'eau. Leur abondance est souvent limitée par la disponibilité de silice ou de calcium. Certaines éponges peuvent
décomposer les roches ou coquilles calcaires et jouent un rôle important dans le cycle biogéochimique du calcium dans les océans. Cette capacité de dégrader
les coquilles calcaires les pousse parfois à décimer les populations d'huîtres et de palourdes. Les éponges servent d'abris pour de multiples animaux et sont
mangées par certains poissons.
8 Une question de phylogénie
Que penser d’une ressemblance choanocytes – Choanoflagellés ?
Il s’agirait plus d’une convergence toute fortuite que d’une parenté directe. ( affirmation remise en question actuellement, il y aurait un lien )
Le caractère primitif des Spongiaires ne doit pas être recherché au niveau de la spermatogénèse ou de l’ovogénèse, mais bien dans
les points suivants :
- Sytéme nerveux diffus.
- Absence d’organes sensoriels définis.
- Absence de région céphalique.
Mise à jour :
Quand on analyse la diversité des Spongiaires, qui sont représentés par 4 classes entre lesquelles les relations taxonomiques restent incertaines (Calcisponges, Démosponges, Hexactinnelides et Homoscléromorphes), on voit apparaître des variations de structure qui ont pour conséquence d'augmenter la surface du revêtement de choanocytes par rapport au volume de l'éponge. On distingue :
Type Ascon
Cette structure en forme de tube avec vaste cavité centrale n'autorise qu'une petite taille : une grande cavité centrale contiendrait trop d'eau pour qu'elle puisse être mobilisée par les choanocytes. Le type Ascon se retrouve chez les éponges les plus petites et les plus simples.
Type Sycon
En cours de développement, la paroi du corps de l’Eponge peut se plisser, formant des évaginations en doigts de gant dans lesquelles sont confinés les choanocytes et des canaux inhalants. Des fusions de canaux inhalants et une couverture générale par les pinacocytes, ainsi que le développement du squelette dans le mésenchyme renforcent la cohésion de l'ensemble.
Type Leucon
Les canaux à choanocytes eux-mêmes peuvent se plisser et se fragmenter en chambres à choanocytes sphériques. La cavité centrale, proportionnellement plus réduite, subsiste sous forme de canaux qui mènent à l'oscule ou aux oscules dans la forme coloniale. La filtration des particules alimentaires est ainsi optimisée par l’augmentation de surface des choanocytes par rapport à la surface totale du corps.
4. Origine et évolution
On dispose d’une série d’informations qui nous permettent de proposer un scénario pour les trois questions qui nous préoccupent, à savoir :
- Quelle est l’origine évolutive des Eponges ?
- Quelle est la diversité existante au sein des Eponges ?
- Quelles sont les relations de parenté entre les Eponges et les autres groupes étudiés ?
L’étude des couches géologiques anciennes à la surface de la terre nous révèle que la première faune complexe pour laquelle on a de bonnes données paléontologiques est la faune d’Ediacara, présente dans des roches qui ont entre 630–540 millions d’années, période qu’on appelle le Précambrien. On y retrouve des traces de fossiles des traces de fossiles apparentés aux Spongiaires. Cette faune nous révèle que les Spongiaires étaient donc déjà présents longtemps avant le début de la période cambrienne (540 à 490 millions d’années).
Quand on s’intéresse aux spongiaires vivant à l’heure actuelle, on remarque une similarité structurelle remarquable entre les cellules nourricières des Spongiaires, les choanocytes, et un groupe de protistes appelés Choanoflagellés. Les deux types de cellules possèdent un flagelle unique entouré par un collier de fins tentacules (la collerette). On a donc émis l’hypothèse que les Eponges auraient évolué à partir d’un ancêtre de type protiste Choanoflagellé.
Etant donné le faible nombre de caractères morphologiques présents chez les Spongiaires, d’autres chercheurs se sont tournés vers des méthodes de biologie moléculaire, dont le séquençage d’un grand nombre de gènes. Le séquençage de différents gènes conservés au sein des Métaozaires et parmi les Spongiaires a montré que ce groupe est formé de quatre groupes distincts :
- les Calcisponges (Calcarea) qui partagent un squelette de calcaire, tels que Grantia sp ;
- les Hexactinellides tels que Euplecetella sp qui produisent des spicules à 6 pointes formant une construction complexe mais légère, et sont formés d’un choanosyncitium plutôt que d’un choanoderme, et ne présentent pas de pinacoderme continu ;
- les Démosponges comme Halichondria et Haliclona qui possèdent toutes une structure de type leucon et des spicules constitués de silice organisée autour d’une matrice de spongine ;
- les Hémoscléromorphes, le groupe le plus récemment identifié, représentés par une seule famille, les Plakinidae.
Ces différents groupes sont monophylétiques
 .
.
Par ailleurs, le premier génome complet d’un Porifère a été séquencé en 2010, c’est celui d’Amphimedon queenslandica. Sur base de l’arbre phylogénétique qui en découle, on constate que les Porifères sont à la base des Métazoaires, et que l’organisme le plus proche des Eponges est un bien un Protiste, Monosiga sp. Le séquençage du génome complet d’A. queenslandica a permis de monter que cette éponge comprend la plupart des gènes utilisés dans la formation des tissus épithéliaux (adhérence cellulaire, communication intercellulaire, protéines kinases,…). Ce type de tissus est un élément essentiel pour l’élaboration de plans de structures morphologiques complexes et le développement d’organismes de grande taille. Les tissus épithéliaux permettent en effet de créer une compartimentalisation entre organes et entre le corps et le milieu extérieur. On peut maintenant tester l’existence d’epithelia dans un organisme en analysant la présence et l’expression des gènes impliqués dans la formation de ces epithelia. En effet, ces gènes sont conservés (i.e. sont les mêmes) pour les insectes, les nématodes et les vertébrés, et sont donc vraisemblablement les mêmes pour tous les animaux.

Comme nous l’avons vu, les éponges ne possèdent pas de tissu épithélial à proprement parler et pourtant, possèdent de nombreux gènes qui les produisent chez d’autres groupes taxonomiques. Par contre, on ne retrouve pas la plupart de ces gènes dans les génomes de Protistes. Ces gènes caractéristiques des tissus épithéliaux sont donc apparus au sein de l’ancêtre des Métazoaires.
2.Eumétazoaires / CNIDAIRES

1 Position systématique
Embranchement des Cnidaires
2 Définition
- Métazoaires diploblastiques à organisation relativement simple.
- Le corps est une gastrula dont l’unique orifice cumule les fonctions anale et buccale et est entouré d’un nombre variable de tentacules creux chargés de la capture.
- Présence d’une mésoglée anhiste contenant des cellules diverses d’origine ectoblastique.
- Par rapport aux Spongiaires, les Cnidaires présentent une symétrie radiaire d’ordre pair ( 4 ou 6 ) pouvant être altérée par une symétrie bilatérale.
- Pas d’appareil respiratoire, circulatoire, excréteur mais des organes des sens : les cnidoblastes
- Animaux marins vivant isolés ou en colonies.
1° les formes benthiques ou polypes sont fixées.
2° les formes nageuses ou méduses sont planctoniques.
- Cycle vital : forme polype – forme méduse sexuée et gonochorique – œuf – larve planula – polype.
- Reproduction sexuée et asexuée et faculté de se régénérer.
3 Morphologie - anatomie
Exemple choisi : L’hydre d’eau douce ( brune,grise ou verte ).
Cas aberrant puisqu’elle est dulcicole, n’existe que sous forme de polype, n’a pas de squelette et est hermaphrodite, mais intéressant pour des
études histologique
3.1 Anatomie
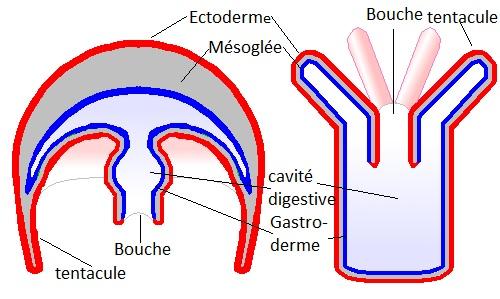
Ils présentent tous une Cavité gastrique ou gastrale ou gastrovasculaire dérivant de l'archentéron et possédant un orifice, la bouche,qui apparaît au stade gastrula.
Leur symétrie est radiaire par rapport à leur axe apico-basal.
Ils sont libres ou fixés. Souvent dans la même espèce existe une alternance de deux types : le Polype, fixé, donne naissance par voie asexuée à la forme libre ou Méduse qui se reproduit par voie sexuée pour redonner un polype. Ce phénomène, nommé Polymorphisme, n'est connu que depuis peu : les naturalistes avaient autrefois donné des noms différents à ces deux phases.
.1.1 Anatomie de la phase polype.
Un polype est un petit sac dont l'ouverture, la bouche, est entourée d'une couronne de tentacules dans lesquels se prolonge la cavité gastrovasculaire. La mésoglée est mince chez le polype.


3.1.2 Anatomie de la phase méduse
La face Aborale ou opposée à la bouche est fortement élargie et prend une forme convexe : c'est l'Ombrelle.
La face orale est concave, la bouche s'ouvre à l'extrémité d'une pseudotrompe appelée Manubrium.
L'ouverture de la face orale peut être rétrécie par une sorte de diaphragme, le Vélum, constitué d'un repli d'ectoderme rempli de mésoglée.
Cette mésoglée, épaisse dans l'ombrelle, réduit la cavité gastrovasculaire à un réseau de canaux qui partent du manubrium vers
l'extrémité des tentacules.Il existe un appareil, le Canal circulaire, qui relie les canaux radiaires entre eux.

3.2. Structure histologique
3.2.1 Les cellules myo-épithéliales
Ces cellules appartenant à l'ectoderme ou à l'endoderme sont également appelées cellules épithéliomusculaires. Elles sont pourvues de myofibrilles du côté de la mésoglée.
L'orientation des fibrilles musculaires n'est pas quelconque : elle est longitudinale dans l'ectoderme. Leur contraction provoque un raccourcissement de l'animal ; dans l'endoderme elle est circulaire et la contraction des fibres provoque un rétrécissement ou un allongement de l'animal.
3.2.2 Les cellules nerveuses
Ces cellules forment deux plexi nerveux situés pour l'un à la base de l'ectoderme, pour l'autre à la base de l'endoderme. Elles transmettent
l'information à des cellules sensorielles réparties dans les deux feuillets.
3.2.3 Les cellules ectodermiques spécifiques
3.2.3.1 Les cellules intersticielles
Ce sont de petites cellules à caractère embryonnaire qui produisent les cellules germinales et capables de se différencier pour remplacer d'autres cellules : les Cnidoblastes.

3.2.3.2 Les cnidoblastes
Ce sont les cellules caractéristiques des Cnidaires.Elles sont abondantes dans les tentacules, urticantes et servent à capturer les proies.Elles sont constituées par un noyau, un cytoplasme et une capsule spéciale, le Cnidocyste. Un filament cytoplasmique immobile dépasse à l'extérieur : c'est le Cnidocil à structure microfibrillaire classique accompagnée d'une fibre dont les caractéristiques peuvent être rapprochées des fibres nerveuses.Le cnidocyste renferme un long filament invaginé portant sur sa face interne, à forte section, des épines et, à faible section,des barbes ; un liquide urticant contient des paralysants musculaires dont l'un est l'actinocongestine. Le tout est fermé par un Opercule.
Lorsque certains corps étrangers touchent le cnidocil, il y a une contraction violente des cellules myo-épithéliales proches, ce qui induit une forte augmentation de pression à l'intérieur du cnidocyste. Le premier ensemble d'épines du filament fait alors fonction de percuteur et éjecte l'opercule. Le filament se dévagine alors en doigt de gant, colle à l'intrus grâce à ses barbillons ; les épines perforent sa chair et les toxines sont injectées. Ce phénomène se déroule à l'échelle de la milliseconde.
Le cnidoblaste se différencie à l'intérieur de l'ectoderme à partir d'une cellule intersticielle et ne parvient à la surface que lorsqu'il est mûr. Il ne sert qu'une seule fois.

3.2.4 Les cellules endodermiques spécifiques
3.2.4.1 Les cellules épithéliales
Ces cellules possèdent de deux à cinq flagelles et ont une activité phagocytaire. Entre les cellules myo-épithéliales existent des cellules glandulaires dont les enzymes sont déversées dans la cavité gastrovasculaire et digèrent partiellement les particules alimentaires avantnleur phagocytose.
3.2.4.1 Les cellules basales
Ces cellules sont de type embryonnaire, souches des cellules glandulaires et situées à la base de l'endoderme.

4 Reproduction
4.1 La voie sexuée
Comme exemple, un hydrozoaire,Obélia , qui forme des colonies à la surface des stipes des laminaires.
4.1.1 Larve Planula : Après développement embryonnaire, larve ciliée nageuse, didermique, qui se fixe sur un support
.1.1.1 Le développement embryonnaire
La segmentation de l'oeuf est totale, égale et conduit à une blastula ciliée, ovoïde et nageuse.

La gastrulation se fait par Migration unipolaire.

Des cellules tombent dans le blastocoele et le remplissent, ce qui amène l'organisme au stade de larve Parenchymula à ectoderme cilié et endoderme plein.

Les cellules endodermiques s'organisent autour d'une cavité appelée Archentéron, l'organisme passe alors au stade de larve Planula creuse, à vie pélagique.

La planula se fixe sur un support, s'aplatit, puis s'allonge en prenant la forme d'un polype. La bouche se perce à la partie supérieure, les tentacules se forment à partir de cet orifice.

4.1.2 La phase polype
Croissance de la planula et formation du premier hydranthe bientôt suivi d’autres par bourgeonnement. Extension de la colonie par allongement et ramifications.
4.1.3 Apparition des gonanges
Polypes dépourvus de bouche et de tentacules et se composant d’un axe fertile blastostyle et d’une enveloppe gonothèque. Le blastostyle va engendrer par bourgeonnement la forme méduse . Les gonocytes émigrent au niveau du blastostyle.
4.1.4 La phase méduse
petites méduses à vélum ou méduses craspédotes portant les gonades ( ensemble de gonocytes ).
Ces méduses s’échappent par l’ouverture de la gonothèque. Toutes les méduses issues d’une même colonie ont le même sexe.
La fécondation a lieu dans l’eau, formation de la Planula.

4.2 La voie asexuée
Chez certaines espèces où la phase méduse est majoritaire, il y a un phénomène de Strobilisation, multiples étranglements du polype permettant de libérer les petites méduses. C’est une forme de bourgeonnement
4.3 La régénération
Le pouvoir de régénération est considérable, a condition que le morceau envisagé contienne de l’ectoderme et de l’endoderme.
Une hydre coupée en morceau redonne des animaux complets et la polarité primitive est respectée.
Ce pouvoir s’explique par la présence de cellules intersticielles.
CYCLES
Fondamentalement le cycle de développement des Cnidaires passe par une alternance entre phase polype et méduse. La phase sexuée étant la méduse. Mais à partir de ce schéma il y a de nombreuses variations possibles et une des phases, aussi bien l'une que l'autre, peut être escamotée.
a) Cycle général

b) Exemple de cycle sans polype

c) Exemple de cycle sans stade méduse

d) Cycle de reproduction d'un Hydrozoaire,avec alternance méduse- polype (La forme polype constitue des colonies à la surface de certaines algues, elle donne des axes de 2 à 3 cm de long. Les tubes ramifiés qui courent à la surface de l'algue constituent l'hydrorhise, les axes dressés, l'hydrocaule, les polypes sont les hydrantes.
L'ectoderne périphérique secrète un fin squelette ectodermique le périsarc la partie charnue est le coenosarc. Au niveau des hydrantes le périsarc donne une logette l'hydrothèque portée par un pédoncule articulé..A maturité cette colonie forme des gonanges ou gonozoïdes, ce sont des polypes dépourvus de bouche et de tentacules qui vont bourgeonner des méduses. Les bourgeons médusaires apparaissent progressivement de haut en bas à la surface du blastostyle. Quand l'organisation de la méduse est parfaitement définie il y a étranglement du pédoncule de fixation et libération de la méduse (1mm de diamètre). Toutes les méduses issues de la même colonie ont le même sexe.
La méduse libre possède sauf exception un velum , elle est dite craspédote, les gonocytes sont logés dans la parois des canaux radiaires. Ces méduses au terme de leur croissance mesurent environ 5 mm de diamètre, elles assurent la dissémination des produits génitaux. Toutes les médusess issues d'un même gonozoïde sont du même sexe. La fécondation a lieu dans l'eau de mer ou dans la cavité sous ombrellaire. Le développement embryonnaire conduit à une larve didermique la planula qui est ciliée et nageuse, elle se fixe après quelques heures, elle s'étale sur le support en étoile dont les branches constituent les ébauches des hydrorhizes.

c) Cycle d'Aurelia avec alternance de stade méduse dominant et stade polype

.5 . Biologie – physiologie
5.1 Respiration et circulation
Les Cnidaires dépendent principalement de la diffusion pour obtenir l'oxygène dont ils ont besoin. Leur corps étant très souvent formé de deux couches de cellules, l'une à l'extérieur, l'autre tapissant la cavité gastro-vasculaire, ils n'ont pas vraiment besoin de système circulatoire. Les cellules amiboïdes de la mésoglée s'occupent du transport des éléments nutritifs des cellules du gastroderme vers celles de l'épiderme.
5.2 Alimentation et digestion
Les Cnidaires sont, la plupart, des carnassiers qui se nourrissent de plancton, de protozoaires, et de petits poissons. Ils capturent et immobilisent leurs proies à l'aide des cnidoblastes de leurs tentacules. Souvent ces cnidoblastes contiennent des toxines qui paralysent la proie. La proie est transportée vers la bouche à l'aide des tentacules.
La digestion est à la fois extracellulaire et intracellulaire chez les Cnidaires. Les cellules du gastroderme sécrètent du mucus et des enzymes digestives qui sont relâchées dans la cavité gastro-vasculaire. Les cellules du gastroderme sont flagellées, et le battement des flagelles permet de mélanger l'eau, la nourriture, et les enzymes contenues dans la cavité. Les particules alimentaires partiellement digérées sont ensuite absorbées par phagocytose, et la digestion est complétée à l'intérieur des vacuoles digestives.

Chez les Anthozoaires, il y a une structure unique, le siphonoglyphe, qui est une gouttière ciliée à chaque extrémité de la bouche allongée. Cette structure permet de faire circuler l'eau vers l'intérieur de la cavité gastro-vasculaire. Les cellules du siphonoglyphe sécrètent également du mucus qui sert à lubrifier la bouche et le pharynx et ainsi faciliter le passage de la proie vers la cavité gastro-vasculaire. La présence des septa, chez les anémones de mer augmente la surface de contact entre le contenu de la cavité et le gastroderme.
Notez que la bouche est à la fois l'anus chez les Cnidaires. Il n'y a qu'une seule ouverture au tube digestif, et on dit de cet arrangement que c'est un tube digestif incomplet. Cet arrangement n'est pas très efficace car la nourriture partiellement digérée, les rebuts de digestion, et les proies nouvellement ingérées sont mélangées dans la cavité gastro-vasculaire. Il est donc difficile à ces animaux de digérer parfaitement leurs proies car les gradients de diffusion des substances nutritives et des déchets métaboliques de chaque coté des membranes des cellules du gastroderme ne peuvent être maintenus.
5.3 Excrétion et osmorégulation
Les Cnidaires marins sont isotoniques à l'eau de mer, et n'ont donc pas de problèmes d'osmorégulation. Les déchets azotés sont éliminés sous forme d'ammoniac. L'hydre, qui est dulcicole, est hypertonique par rapport à son milieu. Les surplus d'eau sont éliminés activement par le gastroderme, et les protéines de la membrane des cellules du gastroderme transportent activement les ions pour compenser la diffusion.
5.4 Sens et système nerveux
Il n'y a pas de tête ni de cerveau chez les Cnidaires. Le mode de vie sessile ou planctonique implique que l'animal peut venir en contact avec des proies ou de prédateurs dans toutes les directions. Une concentration des fibres nerveuses dans une région du corps n'est donc pas particulièrement avantageuse.
Les Cnidaires ont un système nerveux primitif (réseau nerveux) composé d'environ 100,000 neurones en réseau qui est en contact avec les cellules contractiles de l'épiderme et du gastroderme, et qui forment des extensions au travers de l'épiderme et du gastroderme. Ces cellules nerveuses transmettent les messages aux cellules contractiles. Il y a deux types de réponses musculaires, des réponses lentes et des réponses rapides qui sont obtenues par des nerfs de diamètres différents.
Les Cnidaires possèdent des cellules sensorielles qui réagissent aux stimuli chimiques et tactiles dans l'épiderme et le gastroderme. Les polypes n'ont généralement pas d'organes sensoriels, mais les méduses ont souvent des cellules photoréceptrices et des groupes de cellules permettant de détecter la gravité: les statocystes. Ces structures sont, dans leur forme la plus simple, un petit sac de cellules ciliées contenant du liquide et des statolithes de sulfate de calcium. Les cellules ciliées sont sensorielles et permettent à l'animal de distinguer la direction du fond et celle de la surface.
5.5 Associations
Symbiose avec des Zoochlorelles, Zooxanthelles qui se fixent dans son endoderme ( avantage alimentaire). Association anémone, poisson clown, etc.
6 Écologie
Les Cnidaires sont typiquement carnivores, mais certaines anémones de mer produisent des cellulases qui leur permettent de digérer le matériel végétal. Ils sont à leur tour des proies pour certains poissons, mollusques et crustacés. Les polypes produisent une quantité phénoménale de mucus pour nettoyer leur surface des particules qui sédimentent, et de nombreux poissons coralliens se nourrissent de ce mucus. Certains vers plats digèrent des polypes sans affecter les cnidoblastes et arrivent à intégrer ces cnidoblastes à leur épiderme pour assurer leur propre protection.
L'anémone et le poisson-clown forment un exemple classique de symbiose. Le poisson s'enrobe du mucus de l'anémone, ce qui empêche l'anémone de le reconnaître comme une proie. L'anémone assure protection au poisson qui se nourrit des déchets rejetés par l'anémone. En contrepartie, le poisson sert de leurre pour attirer d'autres proies vers l'anémone.
Plusieurs coraux possèdent des algues photosynthétiques dans des vacuoles. Les algues produisent des sucres qui sont assimilés par les coraux, et débarrassent les coraux des déchets azotés.
7. Classification des Cnidaires
Il existe quatre classes.
Classe des Hydrozoaires
Les Hydrozoaires sont la seule classe qui possède des espèces d'eau douce. Ils présentent l'alternance typique polype-méduse.
Le polype ne possède pas de pharynx, la bouche est située à l'extrémité d'un cône saillant appelé Hypostome. La cavité gastrovasculaire n'est pas délimitée par des cloisons.
Les méduses possèdent un vélum et sont donc qualifiées de Craspédotes.
Les gonades sont d'origine ectodermique et disposées en position sous-épidermique.
Classe des scyphozoaires
La classe des Scyphozoaires rassemble des animaux en forme de coupe et la phase méduse y est largement prédominante.
Elles ne possèdent pas de vélum et sont dites Acraspédotes. Les gonades sont d'origine endodermiques en position sous-endodermique.

Classe des Cubozoaires
La classe des Cobozoaires rassemble des méduses de petite taille dont l'ombrelle présente une concavité en forme de cube
Classe des Anthozoaires
La classe des Anthozoaires n'existe que sous la forme de polype . Ces animaux peuvent être solitaires ou coloniaux. Ils possèdent un pharynx d'origine ectodermique , la cavité gastrovasculaire est divisée par des cloisons radiales . Au niveau du pharynx existent une ou deux gouttières ciliées appelées siphonoglyphes qui déterminent un plan de symétrie bilatérale . Les autres ont une symétrie radiaire
7.1 Classe des Hydrozoaires
a) Sous classe des stromatoporides
Ils sont généralement rattachés au Hydrozoaires et sont tous fossiles
Leurs colonies sont des formations calcaires massives , incrustantes, dont l'envergure et la hauteur peuvent toutes dépasser 1 m . Elles sont formées d'une superposition de fines couches horizontales de calcaire , les Laminae , parmi lequelles on on distingue des couches des couches claires et sombres , reliées entre elles par des piliers verticaux plus ou moins longs .
De nombreux stromatroporides sont ornés de réseaux étoilés , les Astrorhizes, fins canaux verticaux se terminant à la surface des lamelles par un dessin rayonnant . La surface externe des stromatoporides porte souvent des monticules verruqueux, parfois ornés d'astrorhizes. Chez les espèces les plus anciennes , les Laminae constituent l'élément principal de la structure , les piliers restant courts . Les spécimens plus récents ont des piliers plus grands qui traversent un nombre de plus en plus grand de lamelles. Enfin, les espèces qui datent de la fin du paléozoïque peuvent présenter une structure relativement irrégulière.
Cet ordre est resté durant l'Ordovicien assez peu représenté , mais il constitue un élément important répandu et caractéristique de la formation des récifs au Silurien et au Dévonien.
b). Sous classe des Hydraires
Le type des Hydraires est Hydra ou Hydre d'eau douce existant sous forme de polype isolé et présentant un pouvoir de régénération important . C'est un polype simple sans périderme protecteur et qui peut se multiplier par bourgeonnement .
Reproduction asexuée par bourgeonnement :
- Apparition sur la colonne gastrique d'une hernie formée d'ectoderme et d'endoderme, dont la cavité gastro-vasculaire communique avec celle de l'Hydre-mère.
- Allongement progressif du bourgeon.
- Perforation de la bouche et apparition des tentacules. Le bourgeon peut se détacher de l'Hydre-mère par un pincement de sa base. Plusieurs bourgeons peuvent coexister sur une même Hydre et former une colonie temporaire.
Chez l'Hydre, comme il n'existe pas de stade de méduse, la reproduction sexuée se fait aussi par le polype. Formation de gonades à partir de cellules interstitielles : les gonades mâles dans la partie supérieure de la colonne gastrique, les gonades femelles dans la partie moyenne.
Hydra viridis est donc hermaphrodite, mais présente le phénomène de fécondation croisée, parce que la maturation des deux types de gonades est différenciée temporellement.

b1) Hydraires calyptoblastiques
Chez Obélia la phase polype est coloniale , elle est formée par un stolon ou tige sur laquelle bourgeonnent d'autres individus
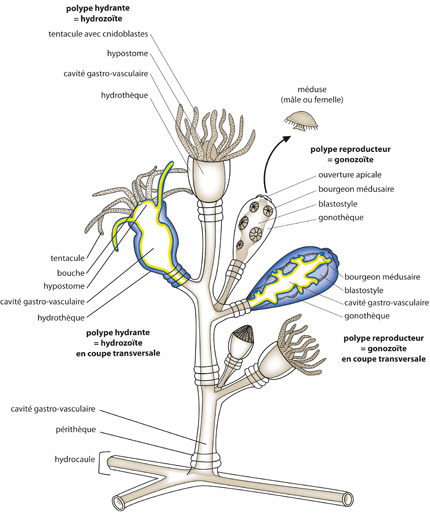
L'hydrorhize émet des stolons dressés et ramifiés appelés Hydrocaules. Chaque ramification se termine par un polype.
Hydrorhize et hydrocaule sont des tubes formés par deux feuillets cellulaires avec de la mésoglée. L'intérieur du tube est la cavité gastrovasculaire qui communique ainsi entre chaque individu . L'ensemble est appelé coenosarc et est entouré d'une membrane et est entouré d'une membrane chitineuse appelée ,périsarc .
Le périderme entoure les différents polypes et forme à leur niveau des loges en forme de coupes nommées hydrothèques ou Gonothèques en fonction du type de polype qu'elles protègent.
Il existe deux types de polypes .
- Les polypes nourriciers ou Hydrantes sont chargés de capter les particules alimentaires à l'aide d'une couronne de tentacules . Ils sont protégés par une hydrothèque.
- Les polypes reproducteurs ou gonantes sont formés d'un axe appelé blastostyle sur lequel bourgeonnent de petites méduses présentant ocelle et statocystes.
 b2 Hydraires gymnoblastiques.
b2 Hydraires gymnoblastiques.
Ce sont des hydraires dont les polypes ne sont pas protégés par un périsarc. Le type des hydraires gymnoblastiques est le genre Hydractinia.
Ils présentent trois types de polypes :
- Les polypes nourriciers sont appelés gastrozoïdes ( ïtes)
- Les polypes reproducteurs sont des gonozoïtes
- Type de polype protecteur ou dactylozoîtes












------------------------------------------------------------------------------------------------------------------------------------------
Mise à jour
La diversité chez Les Cnidaires
9,000 espèces décrites.
Nous n’allons pas voir ici en détail les données morphologiques et moléculaires qui permettent de retracer l’origine évolutive, la diversité, et les relations de parenté que le groupe des Cnidaires entretient avec les autres embranchements de Métazoaires. Ces questions seront abordées au cours de vos travaux de groupe, ou, pour les biologistes, dans la suite de votre cursus universitaire. Sachez cependant que la réponse aux trois questions qui nous préoccupent a changé, pour les Cnidaires comme pour la majorité des groupes taxonomiques vus dans ce cours, de manière importante au cours des dix dernières années. Ceci grâce au développement des techniques de séquençage d’ADN et d’expression génique. Il est donc essentiel de consulter des sources d’informations fiables et récentes (telles que celles que vous trouvez sur les sites de recensement des publications scientifiques en biologie Pubmed ou Scopus) si vous désirez, ou devez, approfondir la question. Le site « scopus » est accessible depuis le serveur de l’université tandis que le site pubmed est disponible de tout ordinateur gratuitement. Ces sites sont un des principaux moteurs de recherche au niveau mondial pour l’accès en ligne de l’information scientifique vérifiée (au contraire de sites tels que « wikipedia »). Servez-vous de « scopus » notamment pour votre travail de groupe.
On peut tenter de répondre aux trois questions :
1. Quelle est l’origine évolutive du groupe taxonomique ?
Une étape majeure de la complexification des Métazoaires concerne, après l’apparition de la multicellularité, la mise en place de la symétrie bilatérale qui caractérise la grande majorité des Métazoaires. Il est remarquable en effet que la diversité des plan de structure du corps ainsi que la diversité en espèces ait explosé à partir du moment où les animaux ont acquis la bilatéralité : parmi les Triblastiques, on retrouve plus de 30 phyla distincts correspondant chacun à un plan de corps différent, et plus de 41 millions d’espèces décrites alors que les Métazoaires plus primitifs (Eponges, Placozoaires, Cténophores et Cnidaires) ne regroupent qu’à peu près 30,000 espèces. On a les premières traces de méduses (siphonophores) dans la faune d’Ediacara, il y a 680 millions d’années, mais pas de traces d’organismes à symétrie bilatérale. On retrouve des traces de symétrie bilatérale chez les Anthozoaires, mais ces caractères n’étaient pas considérés autrefois comme des preuves de la bilatéralité des Cnidaires parce qu’on pensait que les Aanthozoaires étaient un taxa dérivé au sein des Cnidaires et donc la bilatéralité serait apparue secondairement dans ce groupe. Cependant, différentes phylogénies moléculaires ont maintenant montré que les Anthozoaires sont ancestraux par rapport aux autres groupes de Cnidaires et leurs caractères sont donc ancestraux (on parle de plésiomorphies ) pour le groupe, et potentiellement pour le reste des Métazoaires. Il y a aurait donc un ancêtre commun à la fois aux Cnidaires et aux Métazoaires plus évolués, qu’on appelle « Urbilateralia », qui présentait déjà une symétrie bilatérale.
Certains groupes de Cnidaires seraient donc revenus secondairement à une morphologie radiaire, sans doute parce que cette dernière est plus adaptée à un mode de vie sessile : pour un organisme fixe, la nourriture et les prédateurs peuvent venir de toutes les directions. En conséquence, on inclut donc maintenant les Cnidaires parmi les Bilateralia et ce terme devient donc un synonyme de « Eumétazoaire ».
2. Quelle est la diversité existante au sein du groupe taxonomique ?
Deux classes se distinguent parmi les Cnidaires, les Anthozoaires et Medusozoaires, que vous verrez en travaux pratiques. Les caractéristiques qui distinguent ce groupe des Eponges, et qui caractérisent les groupes plus évolués que les Eponges de façon générale, sont: véritables tissus avec lame basale sous l’epithélium, au moins deux véritables feuillets embryonnaires (ectoderme et endoderme), différenciation cellulaire poussée en rapport avec des fonctions variées (cellules épithélio-musculaires, cellules nerveuses et synapses, cellules sensorielles, …), cavité digestive différenciée avec cellules sécrétrices d’enzymes digestives dans le milieu extracellulaire, absence des choanocytes. Ces caractères sont communs à tous les Eumétazoaires (ou Bilateralia) ; on parle de synapomorphies pour les Eumétazoaires.
Par ailleurs, une série de caractères distinguent les Cnidaires de tout autre groupe animal ; ce sont les autopomorphies du groupe. Parmi celles-ci, les plus importantes sont les cnidocytes , la symétrie radiaire avec tendance à l’acquisition d’une symétrie bilatérale, la musculature d’origine ecto- et endodermiques (et non mésodermique), le stade larvaire de type planula à épiderme cilié avec ectoderme et endoderme et qui est issue de la reproduction sexuée, la présence d’organes rudimentaires tels que les rhopalies, le système nerveux très simple disposé en réseau multidirectionnel (pas de synapses chimiques), et l’existence de deux formes, méduse et polype, adaptées à leur mode de vie. Les Cnidaires se ressemblent aussi par l’absence d’appareil circulatoire ou excréteur et par la cavité gastro-vasculaire à une seule ouverture, la bouche.
Le cas des Anthozoaires est particulièrement intéressant. Etant donné leur position phylogénétique basale au sein des Cnidaires, leurs caractéristiques pourraient refléter celles présentes ancestralement dans le groupe. Il est par exemple remarquable qu’on retrouve des traces de symétrie bilatérale chez les Anthozoaires au niveau la bouche (en forme de fente), du siphonoglyphe et des mésentères internes de la cavité digestive, sans oublier la bilatéralité complète de la larve planula. Ces traces de symétrie bilatérale ont donc été vraisemblablement perdues chez les Cnidaires apparus plus tardivement.
Au sein des Anthozoaires, on peut distinguer les sous-classes des octocoralliaires et des hexacoralliaires qui diffèrent par le pattern de développement des septa mésentériques. Chez les Hexacoralliaires, les polypes présentent 6 septa au stade jeune, et ce nombre est variable en cours de vie des polypes et en fonction des espèces. Le nombre de septas peut s’accroître de façon cyclomérique (par ex. Actinia), ou métamérique. C’est le cas chez Cerianthus : la disposition des septa délimite une loge antérieure, une postérieure et 4 latérales. Les nouveaux septa apparaissent toujours dans la loge postérieure, et refoulent vers l'avant les septa préexistants. Chez les Octocoralliaires par contre, on compte 8 septa chez le jeune comme chez l'adulte.
D’autres caractères morphologiques distinguent les Hexacoralliaires des Octocoralliaires. Parmi les hexacoralliaires, les espèces solitaires (comme Actinia) sont généralement dépourvues de squelette, tandis que les espèces coloniales sont généralement pourvues d'un squelette calcaire ou corné très développé, appelé polypier.
Chez les Madrépores, le squelette calcaire est une sécrétion de l'ectoderme. Chaque polype sécrète une base et une muraille qui l'engaine à la partie inférieure. Des projections calcaires se développent également en cloisons vers l'intérieur du polype. Ces animaux ont ainsi édifié au cours des ères géologiques les récifs barrières, les îles coralliaires, les atolls. Actuellement, ils s'accroissent de 20 à 40 mm par an.
On distingue au sein des Medusozoaires de nombreux clades dont des représentants seront vus en travaux pratiques. Pour information, notamment pour ceux qui suivront le stage de biologie marine en deuxième quadrimestre, on peut retrouver toute la classification et la description des caractères propres à chaque famille de Cnidaires sur le site suivant : www.ucmp.berkeley.edu/cnidaria/cnidaria.html
La position des Cnidaires au sein des Métazoaires est encore l’objet d’âpres discussions. Il est cependant admis par la majorité des biologistes que les Cnidaires forment un clade avec le groupe des Cténophores, qu’on appelle « Cœlentérés ». Les Cténophores, qui contient les organismes que nous appelons en langage courant les « groseilles de mer », ne seront pas vus dans ce cours. Les Cœlentérés sont le clade sœur des Bilateralia, qui regroupent tous les animaux que nous verrons dans la suite de ce cours. Les Cnidaires sont donc à la base de la majorité des Métazoaires et cette position primitive explique l’intérêt qu’on porte à l’étude de leurs caractères principaux, tels que cellules musculaires, tissus, organes, épithelia,….
Date de dernière mise à jour : 12/01/2019
Ajouter un commentaire