

classification 1 Présentation R.animal
5. PRESENTATION DU REGNE ANIMAL.
5.1 Présentation des principes de la classification. ( Beaumont-Cassier).
Dès l’Antiquité, l’accroissement incessant du nombre des formes animales recensées suscita chez les naturalistes le désir d’en faire un inventaire complet et, soucis de tout collectionneur, d’en effectuer la classification .
Dans cet ordre d’idées, les premières classifications sont imparfaites, artificielles voire subjectives car elles n’utilisaient qu’un seul caractère pris au hasard. Cette démarche méritoire, si elle permettait de regrouper dans un ensemble tous les animaux possédant en commun les mêmes caractères, s’avérait impuissante à dégager les éventuels liens existant entre ces diverses entités juxtaposées qui constituent le règne animal et qui résultent en apparence de créations indépendantes .Cette simple constatation des faits était insuffisante,sans signification.Seul Aristote1 émit l’idée de l’existence de liens entre les différentes formes animales. Donc d’un ordre naturel.
Au XVIIIè siècle, le Botaniste Bernard de Jussieu2 faisant œuvre de systématicien, chercha à établir la classification des végétaux recensés à son époque en utilisant plusieurs caractères et constata qu’ils n’avaient pas tous la même valeur ; les uns dits dominateurs sont très importants et permettent de définir de grands groupes que les autres dits subordonnés permettent de scinder en groupes inférieurs. La découverte de ce « principe de subordination des caractères » permettant d’attribuer à chaque caractère un rang dans la hiérarchie, une place dans l’ordre naturel. De ce fait, la classification est en quelque sorte imposée par la nature elle-même. L’utilisation de ce principe de subordination des caractères permit au zoologiste Cuvier3 d’établir la première « classification naturelle »des formes animales. Il suggère clairement l’existence de liens entre les différents groupes, substituant ainsi à l’image d’une nature chaotique celle d’une nature ordonnée. Ses écrits contiennent en substance la notion d’EVOLUTION , alors qu’il est en quelque sorte le champion du fixisme.
L’existence de ces liens , l’idée d’une filiation possible entre les divers groupes zoologiques sont corroborés par l’étude des formes disparues et de leur enchaînement chronologique. Les formes actuelles ne sont que les derniers maillons de longues chaînes de formes différentes et disparues : ne vouloir envisager que les formes actuelles est , à l’échelle des temps géologiques, réaliser une coupe horizontale de l’arbre généalogique,en observer l’extrémité des rameaux. En fait, la nature vivante ne peut être comprise dans ses rapports, ses filiations, son agencement, qu’en suivant l’évolution des animaux au cour des âges : « La chronologie explique le monde animal tout entier ; les transformations des êtres vivants sont des phénomènes historiques » ( P.P GRASSE) . L’existence de ces filiations , de cet enchaînement naturel des formes animales connues de nos jours est également, en dépit d’un aperçu parcellaire, confirmée par les multiples données de l’anatomie comparée, de l’embryologie descriptive et comparée et de la chimie biologique.
BIBLIOGRAPHIE : Biologie animale, tome I : A.BEAUMONT, P. CASSIER / DUNOD UNIVERSITE.
1) Aristote (384-322 av. J.-C.) Important philosophe grec, disciple de Platon et fondateur d'une école de philosophie : le Lycée. Il fut le précepteur d'Alexandre le Grand.Ses écrits, dont les principaux sont la Physique, la Politique, la Poétique, la Rhétorique et Ethique à Nicomaque, couvrent tout le savoir de l'époque dans des domaines aussi variés que la philosophie naturelle, la psychologie, la morale, la politique, l'art, la zoologie, la métaphysique ou les sciences naturelles pratiques. La pensée aristotélicienne (encore appelée "l'aristotélisme") eut une influence considérable en Occident mais, devenue dogmatique et incontournable, elle finit par empêcher la progression des sciences et des connaissances.
Quelques citations : "L'homme est naturellement un animal politique."(Politique)
"Le commencement de toutes les sciences, c'est l'étonnement de ce que les choses sont ce qu'elles sont."(la Métaphysique)
2) Bernard de Jussieu (1699-1777). Le XVIIIe siècle voit l’activité botanique se développer en France, et le Jardin du Roi, actuel Jardin des Plantes, échanger plantes et graines avec des correspondants du monde entier. À la demande de Louis XV, un célèbre botaniste du Jardin, Bernard de Jussieu, crée en 1759 un jardin botanique dans le parc de Trianon, en limite de Chèvreloup. Ce jardin, organisé par Jussieu selon ses conceptions de la classification des plantes, tout à fait nouvelles pour son temps, disparaîtra après la mort de Louis XV.
3) Pierre-Paul Grassé. Zoologiste et biologiste français (Périgueux,1895 — Carlux, 1985). Il se consacra d'abord à la cytologie des Flagellés parasites et symbiotiques qu'il aborda, l'un des premiers, au microscope électronique. Il étudia également la biologie des termites et des insectes sociaux en général. Ancien titulaire de la chaire d'évolution des êtres organisés de la Sorbonne, il a adopté une attitude critique à l'égard du néodarwinisme et cherché à fonder sur les données de la biologie moléculaire une théorie de l'évolution qui admette un certain effet direct du milieu (Évolution du vivant, 1973).
5.2. L’arbre phylogénétique ou généalogique.
5.2.1. Petite historique des classifications.
Beaucoup de classifications ont vu le jour au fil des années, elles se sont détrônées mutuellement au fur et à mesure du développement des sciences, de l’histoire et des philosophies.
ARISTOTE ( 384 – 322 av J.C. )
Il propose :Environ 400 espèces d’animaux réparties dans deux classes.
-les animaux à sang rouge ou vertébrés.
- les animaux dépourvus de sang rouge ou invertébrés.
LINNE1 (1707-1773 ap J.C.)
Il propose : La notion d’Espèce est énoncée et de cette façon une base nouvelle est apportée à la classification . Il cherche les
affinités existant entre les êtres et pour ce , se base sur des caractères anatomiques, les animaux sont répartis en 6 classes
Vers – Insectes – Poissons – Amphibiens – oiseaux – mammifères.
LAMARCQ2 ( 1744- 1829 ap J.C. )
En 1806, Lamarcq établi son « tableau du règne animal »
Notion de base : présence ou absence de vertèbres.
VERTEBRES : mammifères, Oiseaux, Reptiles, Poissons.
INVERTEBRES : Mollusques, Annélides, Crustacés, Arachnides, Insectes, Vers intestinaux, Radiaires , Polypes
CUVIER3 ( 1769-1832 ap J.C.)
Classification basée sur l’anatomie comparée
Il reconnaît quatre grands plans structuraux dans le monde animal :
- Les Vertébrés, les mollusques, les articulés et les zoophytes.
Tous les animaux peuvent être accrochés à un de ces plans de base.
Ces quatre classifications n’ont plus actuellement qu’un intérêt historique
Arbre généalogique de Cuénot (1940)
Cet arbre présente la forme caractéristique d’un arbre, la complication des formes est croissante dans le temps, c’est – à - dire du pied vers la cime.
Sur le tronc,s’attachent les divers phylums, embranchements ou clades
Un embranchement se divise normalement en classes, les classes en ordres, les ordres en familles.
Un phylum est autonome, il correspond à un pallier évolutif caractérisé par un type original d’organisation.
Seules les parties terminales des phyla sont en vie .
Les écarts existant entre les différents phyla sur le tronc indiquent le degré d’affinité qui les unit.
Actuellement, les biologistes sont unanimes à reconnaître que les bactéries et les cyanophycées ou algues bleues sont les organismes dont la structure est la plus simple.
On les réunit dans un grand ensemble , les Schizophytes ou Acaryotes ou Monères. Ce premier palier sera suivi d’ un second caractérisé par l’apparition de la cellule, c’est – à – dire naissance des Eucaryotes. Ces cellules vont dans un premier stade mener une vie indépendante, ce sont les Protistes .Les Protistes constituent le départ d’un côté vers les clades végétaux et de l’autre côté vers les clades animaux, avec comme clade de base des PROTOZOAIRES.
Il faut remarquer qu’actuellement on considère l’Embranchement des Protozoaires comme dérivé des Schizophytes ( bactéries et algues bleues acaryotes) qui sont parmi les plus vieilles formes vivantes connues.
On émet également l’hypothèse suivante : la séparation entre le Règne animal et le Règne végétal se ferait au niveau du Phylum des Protozoaires qui présente des organismes à tendance animale et à tendance végétale.
Tendance végétale : Organismes autotrophes, aptitudes les dispensant habituellement de la faculté de mouvement et de sentir.
Tendance animale : Organismes hétérotrophes obligés de chercher leur nourriture . Ils ont du évoluer dans le sens de l’activité locomotrice, et par conséquent dans le
sens d’une conscience de plus en plus ample.
Mobilité et conscience.
Il est à remarquer que chez la plante , l’impressionnabilité toute spéciale de la chlorophylle par la lumière pourrait être mise en parallèle avec la sensibilité de l’animal.Or , un système nerveux est avant tout un mécanisme qui est l’intermédiaire entre l’organisme et le milieu de vie. On en déduirait donc que le véritable système nerveux de la plante serait le chimisme qui sert d’intermédiaire entre l’impressionnabilité de la chlorophylle à la lumière et la production d’amidon . Le même plan qui a porté l’animal à se donner des centres nerveux et des nerfs à dû aboutir dans la plante à la fonction chlorophyllienne.

Autre question capitale : Le passage de l’état unicellulaire à l’état pluricellulaire .
Ce sont les METAZOAIRES .
Un métazoaire est un organisme creux ; d’où deux conséquences :
Une différenciation cellulaire apparaît , ce qui est logique puisque certaines cellules resteront en contact avec le milieu extérieur, d’autres tapisseront la cavité interne et certaines migreront entre les précédentes.
Nécessité d’une ontogénèse : de l’œuf , il se formera d’abord un agglomérat de cellules qu’il faudra arranger, structurer, : morula, blastula, gastrula, etc...

BLASTULA ET GASTRULA, segmentation de l’œuf. (Houillon, Coll méthodes)

On distingue :
-les métazoaires diploblastiques :Sacs formés de deux feuillets cellulaires accolés, l’endoderme et l’ectoderme .
- Les métazoaires triploblastiques
Corps formés de trois feuillets cellulaires : endoderme, mésoderme, ectoderme.
A. Les métazoaires diploblastiques
Premier niveau : LA CLASSE DES SPONGIAIRES.
- Peu évolués.
- Pas d’élément de symétrie .
- Système nerveux diffus constitué de cellules éparses peu différenciées, pas de synapses.
- Structure à l’état de feuillets diversement plissés et agencés.
- Cellules différentiées ( choanocytes, pynacocytes amoebocytes, ).
Deuxième niveau : LA CLASSE DES CNIDAIRES
- Symétrie généralement axiale
- Système nerveux en forme de plexus à la base de chacun des deux feuillets, coordination réelle de l’activité grâce aux informations reçues par des organes sensoriels différentiés (chémorécepteurs, statorécepteurs, photorécepteurs).
- Mésoglée épaisse pourvue de cellules migratrices.
Ce sont les méduses, les anémones de mer, les coraux
LA CLASSE DES CTENAIRES (Cténophores)
Longtemps placés avec les cnidaires dans l’ensemble des coelentérés, s’en distinguent par leur symétrie bilatérale, l’absence de cnidoblastes, la présence constante de cellules spécialisées aux propriétés adhésives , localisées sur deux tentacules symétriques plus ou moins ramifiés , les colloblastes utilisés à la capture des proies .leur mésoglée épaisse contient de nombreuses cellules formant un mésenchyme qui peut être considéré comme l’ébauche d’un troisième feuillet .La condensation peu marquée du système nerveux , la présence quasi constante d’un statocyste, la localisation de la bouche et des tentacules annoncent la différentiation d’un axe antéro-postérieur.

B. Les métazoaires triploblastiques
L’étape ultime dans l’évolution des formes animales est caractérisée par l’apparition d’un troisième feuillet embryonnaire, le mésoderme qui se substitue à la mésoglée des organismes diploblastiques, cette apparition permet de définir des organismes triploblastiques qui ,comparativement aux animaux précédents , atteignent un niveau plus élevé de complexité.Ce troisième feuillet donne naissance au tissu squelettique, au tissu conjonctif, aux muscles à contraction rapide, au sang, aux organes excréteurs et reproducteurs et participera à la formation d’une cavité corporelle .
Tous ces animaux posséderont des organes individualisés assurant une fonction précise.Les animaux en devenant triploblastiques ont franchi un grand pas vers une haute complexité.A part les échinodermes, nous allons rencontrer le phénomène de céphalisation : concentration de récepteurs sensoriels et de centres nerveux dans la région antérieure du corps.
Le mésoderme participant à la formation des cavités du corps peut prendre trois aspects :
Le mésoderme reste massif entre les deux feuillets , il forme un parenchyme de remplissage entre la paroi et le tube digestif, seule cavité .Ce sont les métazoaires triploblastiques acoelomates . ( Vers plats, Némertes, Rotifères…)
Le mésoderme ne donne pas de parenchyme , entre paroi et tube digestif, grande cavité contenant des massifs musculaires , les organes génitaux et excréteurs, cette grande cavité correspond à la blastula et sera appelée blastocoele .Ce sont les métazoaires triploblastiques pseudocoelomates.(Vers filamenteux, Némathelminthes… )
Le mésoderme forme un parenchyme qui se creuse, les cavités qui se forment s’appelleront coelome, dans ces cavités se formerons les viscères.Ce sont les métazoaires triploblastiques coelomates. (Mollusques, Arthropodes, Annélides, Lophophoriens vertébrés…)
Dans la phylogénie récente, la distinction entre coelomates et acoelomates tend à s'effacer, le rynchocoele des Némertes étant considéré comme un coelome et les plathelminthes considérés comme un stock ancestral de protostomiens qui ont évolués d'une part vers le groupe des Ecdysozoaires et d'autre part des Lophotrochozoaires. Les protostomiens lophotrochozoaires possèdent soit un lophophore ( couronne ciliée bucale ), ce sont les Lophophorates ou Lophophoriens, soit une larve trochophore avec un mode de division de l'oeuf spiralé, constituant le groupe des Eutrochozoaires ou Trochozoaires . On a donc parmi les Lophotrochozoaires deux groupes, celui des Trochozoaires et celui des Lophophoriens. Au sein des Trochozoaires deux groupes s'individualisent encore : les Syndermates dont la division de l'oeuf ne serait pas spiralée mais la larve trochophore et les organismes à développement de l'oeuf spiralé, ce sont les Spiralia ou Spiraliens.à larve trochophore. Les Protostomiens Ecdysozoaires sont caractérisés eux comme présentant des mues successives au cours de leur développement et dont le corps est recouvert de 3 couches de kératine alpha.
Pourquoi un bifurcation au niveau des Métazoaires triploblastiques ?
Traditionnellement, deux grands plan structuraux imposent cette bifurcation aux évolutionnistes.

Caractéristiques et apparition du coelome.
Les modalités qui aboutissent à la formation du mésoderme sont à voir dans la rubrique embryologie, les mouvements cellulaires qui y aboutissent regroupent divers mécanismes nommés : embolie, épibolie, délamination...
Chez les organismes triploblastiques coelomates, les cellules mésodermiques constituent de part et d’autre du tube digestif, des massifs cellulaires pairs, symétriques, qui s’organisent , au moins au cours du développement embryonnaire en vésicules closes ou vésicules coelomiques. L’ensemble de ces vésicules nées par entérocoelie ( super phylum des Echinodermes) ou par schyzocoelie ( super phylum des Annélides ) représente le coelome.
Formation par schizocœlie : des fissures se forment au sein du mésoderme et convergent pour former une cavité unique de part et d’autre de l’embryon.
Formation par entérocœlie : de chaque côté de l’endoderme se créent des dépressions qui détachent le mésoderme et forment ainsi la cavité cœlomique.
Cette segmentation ou fragmentation du mésoderme (mésomérie) est accompagnée d’une distribution périodique des masses musculaires (myomérie), nerveuses (neuromérie),excrétrices ( néphromérie) ou génitale, ce qui correspond à une division du corps en unités anatomiques et fonctionnelles, les métamères, qui extérieurement concrétisent leur individualité par des constrictions antérieures et postérieures correspondant au plan d’affrontement des cavités coelomiques successives. La coordination de l’activité des métamères successifs est assurée par les centres nerveux antérieurs.
Cette métamérisation est accompagnée par une condensation, voire même une céphalisation, des éléments nerveux antérieurs , phénomènes qui atteignent leur apogée chez les Céphalopodes, les Insectes et les Vertébrés. L’ensemble des organismes triploblastiques coelomates est très hétérogène , l’observation de la destinée du blastopore permet de disjoindre deux lignées évolutives naturelles, les protérostomiens et les deutérostomiens qui par leurs autres caractères présentent une parenté phylétique. pour distinguer ces modalités sous forme de schéma, voir notre page

a) Les protérostomiens
Le blastopore qui, chez les diploblastiques assurait la double fonction de bouche et d’anus, constitue la bouche et ne donne jamais l’anus . Il peut aussi complètement disparaître ; bouche et anus étant dans ce cas des néoformations. Il peut également s’étirer et former une fente longitudinale dont les bords se soudent dans la partie moyenne, délimitant ainsi une ouverture antérieure à l’origine de la bouche et une ouverture postérieure à l’origine de l’anus.
Chez les protérostomiens, la métamérisation reste très nette chez les formes inférieures, même à l’état adulte ; elle s’estompe fréquemment chez les formes supérieures. Le système nerveux de ces Métazoaires est condensé, métamérisé et comprend une paire de ganglions cérébroïdes antérieurs et dorsaux associés à une chaîne nerveuse ventrale comportant une paire de ganglions par métamère, située sous le tube digestif ; cette disposition qui leur vaut le qualificatif d’Hyponeuriens se rencontre chez les Annélides, les Mollusques, les Arthropodes et dans quelques autres embranchements de moindre importance ( Sipunculiens, Echiuriens,Onychophores, Lophophoriens). De manière résumée, l'on a :
LOPHOTROCHOZOAIRES
Trochozoaires syndermates : Rotifères, Acantocéphales,
Trochozoaires spiraliens : Entoproctes, Plathelminthes, Némertes, Mollusques, Sipunculiens, Annélides.
S'opposant aux Trochozoaires nous avont le second groupe des Lophotrochozoaires constitué par les Lophophorates : Brachiopodes, Phoronidiens, Chaetognathes, et les Ectoproctes.
La parenté phylétique des organismes protostomiens spiraliens est confirmée par l’identité des modalités de leur développement embryonnaire ( type Spiralia) , par la similitude de leurs formes larvaires ( type Trochophore ) et par l’équivalence de leur croissance antéro-postérieure ( individualisation de paires de sacs coelomiques à partir d’une zone postérieure de prolifération ).
ECDYSOZOAIRES
Dans ce groupe, s'individualise le sous groupe des
Panarthropodes avec les Onychophores, les Tardigrades et les Euarthropodes.A côté du groupe des Panarthropodes s'individualise encore le groupe des :
Introvertés : Ils possèdent un introvert dévaginé au repos et leur système nerveux présente un cordon nerveux et un anneau péripharyngial = cycloneuralia ; ce dernier sous groupe comprend les Kynorinches, les Priapuliens et les Nématodes.
et deux groupes libres, les Loricifères et les Nématomorphes.
b) les Deutérostomiens
Chez ces organismes triploblastiques coelomates, le blastopore de la gastrula devient l’anus ou marque l’emplacement de cet orifice ; la bouche est une néoformation. La segmentation n’est jamais spirale ; le mésoderme naît fréquemment par entérocoelie. Le niveau de différenciation et la position du système nerveux permettent de scinder cet ensemble en Epithélioneuriens et Epineuriens (voir plus bas).
5.2 ETUDE DE LA BRANCHE DE GAUCHE
1) Acoelomates
Chez les métazoaires acoelomates, le mésoderme reste compact, il ne s’organise jamais en vésicules closes, ce feuillet joue un rôle effacé ou participe à la constitution d’un tissus diffus, le parenchyme qui comble plus ou moins la cavité viscérale, et à celle de divers organes nouveaux : muscles pariétaux et viscéraux, organes génitaux etc.
L’ensemble des Acoelomates, organismes triploblastiques primitifs comprend les Plathelminthes, les Némertiens, les Némathelminthes, les Rotifères, quelques autres embranchements de moindre importance ( Orthonectides, Dyciémides) voire les Mollusques.
La structure des Métazoaires hyponeuriens est schématiquement la suivante : Répartition le long de l’axe longitudinal du corps de sacs coelomiques, d’où formation de segments ou métamères. Chaque métamère possède une paire de ganglions et une paire d’organes excréteurs. Cette structure archaïque ne se retrouve que chez les Annélides de la classe des Polychètes. Le long de cette branche , les principaux embranchements rencontrés sont les suivants :
LES PLATHELMINTHES
Ce sont des vers plats acoelomates ne possédant pas d’anus et souvent parasites
Ex : le Taenia, la Douve du foie.
LES NEMATHELMINTHES
Vers cylindriques,filamenteux, pseudocoelomates, ce qui fait que le tube digestif, les organes génitaux et excréteurs baignent librement dans le liquide péri viscéral.
Formes libres et parasites.
Ex : les Nématodes, les Ascaris.
LES ROTIFERES
Animaux acoelomates, microscopiques vivant dans les eaux douces et les mousses.
LES LOPHOPHORIENS
Avec cet embranchement nous entrons dans les coelomates
Ce sont les Bryozoaires, les Brachiopodes, etc…
Tous sont des animaux microphages pourvus d’un cercle de tentacules péribuccaux ( Lophophore) ciliés qui acheminent les proies vers la bouche.
2) Coelomates
LES ANNELIDES
Vers supérieurs annelés, conservant, du moins chez les Polychètes, la structure archaïque des Protérostomiens. Trois classes sont à distinguer :
Les Polychètes, vers marins ; ex : Serpula, Néréis
Les Oligochètes, vers terrestres ; ex : le Lombric
Les Achètes ; ex : la Douve
LES MOLLUSQUES
Animaux non métamérisés et formant une clade homogène, malgré les aspects parfois très différents de leurs représentants. On distingue trois grandes classes :
Les Lamellibranches ( Moules, Huitres).
Les Gastéropodes ( escargots).
Les Céphalopodes ( Seiches, pieuvres).
LES ARTHROPODES ( Euarthropodes )
Embranchement immense chapeautant cette branche de droite, riche en espèces et en formes.
On en est arrivés à la spécialisation des appendices segmentaire et à une céphalisation poussée à l’extrême ( fusion de plusieurs segments), accompagné d’une régionalisation des parties du corps ( tête, thorax, abdomen).
La métamérie est bien conservée au niveau de l’abdomen ( voir entre autre la chaîne nerveuse ventrale).
Actuellement, on les divise en deux sous embranchement matérialisant les deux tendances évolutives de ces animaux .
Les Chélicérates : Pas d’antennes, des Chélicères comme pièces buccales ; Principale Classe, les Arachnides.
Les Mandibulates ou Antennates : Antennes, Mandibules.
Trois grandes Classes :
- Les Crustacés
- Les Myriapodes
- Les Insectes
5.3 ETUDE DE LA BRANCHE DE DROITE
Deutérostomiens épineuriens qui peuvent se diviser en deux groupes si on se base sur la différentiation et la position du système nerveux.
a) Les Deutérostomiens épithélioneuriens
Le système nerveux est imparfaitement dégagé de l’ectoderme dont il dérive, la céphalisation est nulle, pas de métamérie.
Trois Embranchements :
LES ECHINODERMES
Animaux caractéristiques par leur symétrie axiale d’ordre 5 se subdivisant à la symétrie bilatérale.
Etoiles de mer, Oursins etc…
LES STOMOCHORDES
Organismes vermiformes présentant un long pharynx portant de nombreuses fentes branchiales.
Animaux marins vivant enfoncés dans la vase.
Ex : les Balanoglosses.
La stomochorde, diverticule pharyngien , dirigée vers l’avant et placée dorsalement ne serait pas ( ? ) l’équivalent de la corde des Chordés.
LES POGONOPHORES
Animaux vermiformes découverts tardivement par les zoologistes, ils vivent en eaux profondes dans des tubes chitineux.

Cet organisme possède un coelome divisé en trois portions,il présente une condensation de plexus nerveux dans la partie antérieure. Absence d’appareil digestif
b) Les Deutérostomiens épineuriens : tube nerveux complètement dégagé de l’ectoderme, céphalisation.
Ce sont les CORDES, (phylum)
Définition : Tous les centres nerveux au dessus du tube digestif.
Axe squelettique dorsal, la corde située entre le tube digestif et le système nerveux.
Présence de fentes branchiales ( au moins à l’état embryonnaire ).
Trois Embranchements :
Les PROCORDES OU TUNICIERS
Les Tuniciers ou procordés, sont de petits organismes vivant dans la mer. Le plan d’organisation de base est conservé chez les larves, et, à l’état adulte, uniquement chez les Appendiculaires . Le coelome est limité à la cavité péricardique , car la majeure partie des bandelettes mésodermiques se dissocie au cours de l’ontogénèse.
Les CEPHALOCORDES
( Amphyoxus, Asymmetron )
Prenons l’exemple de l’Amphyoxus , il s’agit d’un organisme marin fouisseur vivant dans le sable calcaire (Maerl). Ils conservent le plan type d’organisation des Chordés mais leur appareil excréteur est représente par une touffe de protonéphridies. Si l’Amphyoxus ne peut pas être considéré comme l’Archétype des Vertébrés, il présente un intérêt tout particulier car, par son organisation, les modalités de son ontogénèse, il constitue une copie parfaite de cet archétype.
Les VERTEBRES
Définition :
- Corps divisé en trois parties : Tête, siège de la vie consciente.
Tronc, siège des viscères.
Queue, siège de la locomotion.
- Système nerveux fortement céphalisé : cerveau.
- Cerveau enveloppé dans un crâne qui, au cours de l’évolution des vertèbres, passe du stade muqueux, cartilagineux et enfin osseux.
- Moelle épinière logée dans le canal rachidien, dépendance de la colonne vertébrale qui s’est substituée à la chorde primitive. Unité de cette colonne : la vertèbre.

Une partie du squelette branchial est à l’exception des Agnathes ou Cyclostomes, incorporée au crâne et constitue en particulier la mâchoire inférieure.
Le coelome n’est métamérisé que dans sa partie dorsale, ce qui isole des vésicules paires ou somites à l’origine de la masse musculaire pariétale, du squelette axial, de la majeure partie du tissus conjonctif et de l’organe adrénal. Sa partie ventrale indivise ou lame latérale constitue le péricarde, le péritoine et, chez les formes aériennes la cavité pleurale. Elle disparaît dans la région antérieure et dans la queue.
Le Pharynx est soudé aux parois du corps et, chez les formes aériennes, les fentes branchiales ne sont pas visibles, au moins à l’état d’ébauches non perforées au stade embryonnaire. Les Vertébrés primitivement inféodés au milieu aquatique ( Cyclostomes, poissons ), ont, avec l’apparition des Amphibiens, colonisé le milieu terrestre grâce au développement d’un appareil respiratoire aérien, les poumons, qui coexista avec les branchies puis se substitua totalement à elles ; parallèlement, la région caudale régresse, les nageoires paires ( disposition ichtyoptérigyenne) forment des membres dont le plan d’organisation est identique des batraciens à l’homme ; tous les Vertébrés à partir des Batraciens sont des Tétrapodes. Le squelette branchial perd sa vocation respiratoire et participe à la formation du squelette céphalique ventral ou splanchnocrâne ; aux néphrons primitifs se substitue un nouveau rein, le métanéphros ; la céphalisation s’accentue ; la pharyngotrémie ne persiste que chez l’embryon.
Les Cyclostomes ( Lamproies, Myxines) , formes archaïques molles ne représentent à l’heure actuelle que des reliques d’un groupe florissant, riche en espèces et en individus généralement de grande taille et pourvus d’un exosquelette très développé , les Ostracodermes Ils sont généralement connus sous le nom d’Agnathes ( ou Agnathostomes) car il sont dépourvus de mandibule. Le premier arc viscéral n’est pas différentié en mâchoire.
Les Poissons, forment un ensemble très vaste, hétérogène, vraisemblablement polyphylétique où les Chondrichtyens (= Elasmobranches=Sélaciens ) au squelette cartilagineux se distinguent naturellement des poissons osseux ou Ostéichtyens (Dipneustes, Crossoptérygiens, Actinoptérygiens, Brachioptérygiens).
Les Amphibiens réalisent la première étape de la colonisation du milieu terrestre, étape incomplète, puisque leurs larves et les adultes restent inféodés ou font retour, en période de reproduction au milieu aquatique.
A partir des Amphibiens qui leur donnent naissance, divergent deux grandes lignées évolutives : les Sauropsidés ( Reptiles, Sauropsidés, oiseaux ) et les Thérapsidés ( Reptiles,Thérapsidés, Mammifères) qui se distinguent par la constitution de l’oreille moyenne, le mode de suspension de la mâchoire, par l’acquisition de systèmes les rendant indépendants du milieu extérieur ( homéothermie, viviparité, constance du milieu intérieur) et par diverses particularités anatomiques.L’apparition de ces grands groupes au niveau historique correspond à un degré de complexité croissante. L’ère primaire contient en abondance poissons puis batraciens ; l’ère secondaire est l’ère des reptiles ; les oiseaux et les mammifères connus dès la fin du secondaire se diversifient au cours des les Eres Tertiaire et Secondaire.

Nous étudierons en détail le Phylum des Chordés et en particulier l’Embranchement des Vertébrés plus loin dans ce modeste recueil, établissons cependant dès à présent ses grandes divisions.
Evolution de l’Embranchement des vertébrés
A) Point de départ : les Agnathes
Tous les arcs du squelette branchial sont identiques, pas de mâchoire différentiée.
Classe des CYCLOSTOMES
B) Etape Gnathostome
Les premiers arcs branchiaux se transforment en mâchoires supérieures et inférieures.C’est le cas de tous les autres vertébrés.
B.1 Appendices pairs du type nageoire
Poissons cartilagineux : CHONDRICHTYENS
Poissons osseux : OSTEICHTYENS
B.2 Appendices pairs du type pattes : Etape tétrapode
La colonisation du milieu terrestre débute . tous ces vertébrés possèdent des pattes construites sur le même modèle et présentant des adaptations à divers modes de vie.
Apparition de novations structurales fonctionnelles :
Les poumons
Les annexes embryonnaires
B.2.1 Embryon dépourvu d’annexes extra-embryonnaires, anamiotes
Classe des AMPHIBIENS
Seuls les adultes mènent une vie terrestre, les larves continuent à mener une vie aquatique ( pas besoins d’annexes embryonnaires ).
Sous classe des Anoures : Les Grenouilles, les Crapauds.
Sous classe des Urodèles : Les Tritons, les Salamandres.

B.2.2 Embryon possédant des annexes embryonnaires : Etat amniote
B.2.2.1 Mâchoire inférieure articulée sur le crâne par l’intermédiaire du carré et de l’articulaire
Deux crosses aortiques : Classe des REPTILES
Une crosse aortique : Classe des OISEAUX
B.2.2.2 Mâchoire articulée directement sur le Squamosal
Une crosse aortique : classe des MAMMIFERES
Ce paragraphe sur l’Embranchement des Vertébrés n’est provisoirement qu’a parcourir attentivement, il est incomplètement défini et sera largement étoffé de schémas et d’explications plus loin dans cette rubrique.
A partir des Amphibiens, deux grandes lignées évolutives sont à distinguer : les SAUROPSIDES et les THERAPSIDES.
Les Sauropsidés, reptiles actuels, reptiles fossiles ailés et, fer de lance de cette poussée évolutive, les OISEAUX.
Les Thérapsidés, reptiles mammaliens fossiles et, fer de lance, les MAMMIFERES.
La recherche en matière de classification, d’évolution et de phylogénie du règne animal, comme végétal d’ailleurs, continue, si bien
que ce document ne constitue qu’un texte de base qu’il faut régulièrement adapter en fonctions des avancées et découvertes.
Il faut également pouvoir disposer de documentation sérieuse (revues scientifiques) en plus d’une bibliographie de base. Cette documentation permettra d’étoffer vos connaissances et d’exercer vos sens de la comparaison de la critique et de la synthèse .
Lire par exemple, parmi des publications un peu datées :
Pour la science, juillet 1995 : l’essor des mammifères
La Recherche, décembre 1993 : la crise Crétacé Tertiaire
La Recherche, juillet août 1993 : les dinosaures primitifs
Pour la Science, avril 1999 : l’évolution des poissons
Science & vie, décembre 2000 n° 213 hors série : La grande histoire du vivant
Je me permettrai, en humble passionné de sciences naturelles, de considérer notre environnement ( la vie et son décor ) comme une merveille sans borne qui offre, à qui sait observer, attendre et réfléchir, des satisfactions durables et un sens instinctif du bonheur et des choses fondamentales.
Liste des phyla
En général on admet les 19 phyla suivants et parfois quelques autres comme nous l'avons vu ci-dessus :
PROTISTES
FUNGI
BRYOPHYTES
TRACHEOPHYTES
PORIFERES (SPONGIAIRES ).
CNIDAIRES
CTENAIRES
PLATHELMINTHES
NEMERTES
GASTROTRICHES
KINORHYNCHES
PREAPULIDES
ENTOPROCTES
NEMATODES
ROTIFERES
ANNELIDES
ARTHROPODES
MOLLUSQUES
SIPUNCULIDES
ECHIURIDES
POGONOPHORES
ECTOPROCTES
PHORONIDES
BRACHIOPODES
HEMICORDES ( STOMOCORDES )
CAETOGNATHES
ECHINODERMES
UROCORDES
CORDES
5.4 LA PHYLOGENIE
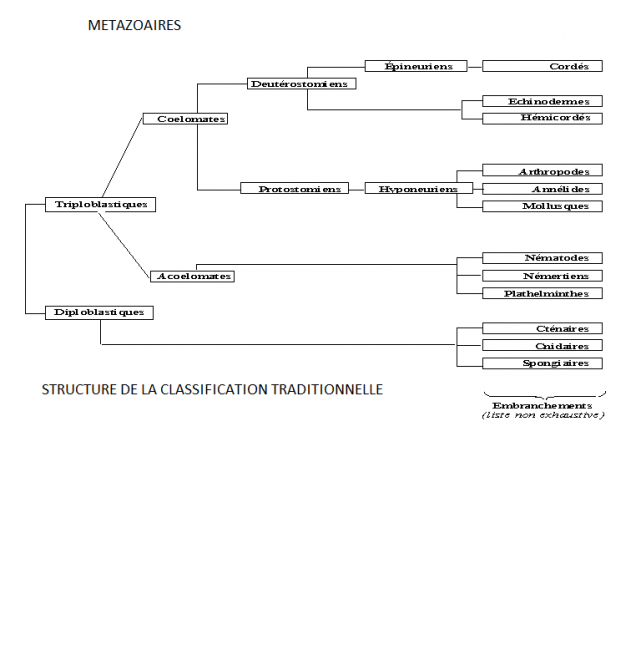
La phylogénie animale présente un paradoxe apparent: des groupes animaux constituant de grands ensembles dénommés "phyla" sont assez faciles à reconnaître, mais les relations entre phyla ont été très difficiles à établir. Les phylas ont été décrits au 19ème siècle et ont été affinés depuis, sur la base d'un "plan d'organisation" de chaque phylum.
La phylogénie traditionnelle
Toutes les espèces appartenant à un phylum donné partagent un ensemble de caractères permettant sans ambiguïté leur assignation à ce phylum. Les exemples les plus caractéristiques sont les arthropodes avec un corps segmenté, une cuticule comportant de la chitine, des appendices articulés, les mollusques avec une radula (une bande cuticulaire rigide utilisée pour la prise des aliments), un manteau (sécrétant habituellement une coquille) et un pied, les échinodermes avec une symétrie pentaradiaire et les vertébrés avec leur notochorde, des fentes branchiales et une colonne vertébrale. Contrairement aux divisions claires des groupes d'animaux en phyla, les relations entre ces phyla ont donné lieu à des controverses pendant les dernières décades. La raison fondamentale en est l'absence de caractères homologues entre phyla, empêchant leur regroupement en noeuds successifs, chacun correspondant à un lot de caractères dérivés (synapomorphies dans la terminologie cladistique). En l'absence de tels caractères chez les adultes, les zoologistes se sont basés récemment sur d'autres critères inspirés par l'idée de la récapitulation, hypothèse populaire que les stades embryologiques successifs des organismes récapitulent leur phylogenèse. Ils ont recherché les grandes similitudes, spécialement pour les stades embryonnaires précoces. Ceci leur a permis de constituer de très grands groupes. Ils ont organisé ces groupes dans un ordre reflétant ce qu'ils considèrent une "complexité croissante", avec des animaux simples à la base de l'arbre et les plus complexes au sommet.
En ce sens, la première démarcation parmi les métazoaires a été placée entre les éponges (perte de l'organisation tissulaire, peu de types cellulaires différenciés, absence de cellules nerveuses) considérées comme les animaux les plus simples, et tous les autres. Les autres métazoaires sont ensuite subdivisés selon le nombre de feuillets. Les Cnidaires (méduses, anémones de mer, coraux, hydres) ont apparemment seulement deux feuillets tissulaires: un ectoderme externe, donnant principalement un épiderme et des cellules nerveuses; un endoderme interne formant la cavité digestive. Les Cnidaires sont quelquefois appelés animaux diploblastiques. Tous les autres animaux ont un troisième feuillet, le mésoderme, situé entre les deux autres et permettant la différenciation de muscles et d'organes internes. Ils sont ainsi appelés animaux triploblastiques. De façon peut-être plus significative, les Cnidaires ont une symétrie radiaire, tandis que tous les animaux triploblastiques ont une symétrie bilatérale primitive (Bilateria est un synonyme de Triploblastica). Le statut des cténophores, des placozoaires (une espèce unique, Trichoplax) et des mésozoaires (petits acoelomates vermiformes constitués de très peu de cellules) est ambigu, mais ils sont le plus souvent placés avec les Cnidaires à la base de l'arbre des métazoaires.
Chez les Bilateria, la division la plus importante a été entre acoelomates et coelomates. A partir de ce point, un poids important a été attribué à la présence ou l'absence de coelome et à son mode de formation (particulièrement dans le travail fondamental de Libbie Hyman). Chez les acoelomates comprenant essentiellement les vers plats (et souvent les némertes), la cavité interne entre ectoderme et endoderme est entièrement remplie par un réseau dense de cellules mésodermiques. Chez les coelomates, le mésoderme est "creux", constitué d'un compartiment empli de liquide, de volume variable, délimité par un épithélium mésodermique, nommé le coelome. Les coelomates vrais ont été ultérieurement en deux grands clades, les protostomiens et les deutérostomiens selon la manière dont la bouche se forme lors de l'embryogenèse, mais également selon un ensemble convergent d'autres caractéristiques. De nombreux protostomiens ont une segmentation embryonnaire spirale tandis que les deutérostomiens présentent un mode de clivage radial. Le coelome est souvent formé par le creusement d'une masse mésodermique chez les protostomiens (schizocoelie), à partir de poches issues de l'intestin primitif chez les deutérostomiens (entérocoelie). Les cordons nerveux sont typiquement ventraux chez les protostomiens tandis qu'il y a un tube nerveux dorsal chez de nombreux deutérostomiens. Les protostomiens comprennent les arthropodes, les Annélides, les mollusques et un ensemble de phyla vermiformes plus obscur (sipunculiens, pogonophores, tardigrades, onychophores). Les deutérostomiens comprennent les échinodermes, les hémichordés (ptérobranches et entéropneustes comme le balanoglosse) et les chordés. Ces derniers sont subdivisés en urochordés (ascidies), céphalochordés (amphioxus) et vertébrés. Les choses sont légèrement pour un groupe de petits Bilateria,apparemment dépourvus de vrai coelome, mais possédant néanmoins des cavités internes souvent dérivées d'un blastocoele embryonnaire. Ces organismes, connus collectivement comme pseudocoelomates (ou aschelminthes), comprennent les nématodes, les rotifères, les nématomorphes, les priapulides et quelques autres
petits groupes. Le point de vue dominant a été de les placer quelque part entre les acoelomates et les vrais coelomates, habituellement dans un super-phylum unique.L'arbre phylogénétique que l'on trouve dans les principaux manuels est comparable à celui de la figure 1, des éponges à la base aux deux branches terminales au sommet, l'une correspondant aux arthropodes, l'autre aux vertébrés. Dans l'intervalle, de nombreuses branches latérales émergent successivement, correspondant aux degrés d'un complexité croissante. Les premiers auteurs ont souvent été plus prudents que ne le suggère cette simplification. Mais il était plus facile d'exposer l'évolution animale comme une saga glorieuse d'une augmentation progressive de la complexité que des générations d'étudiants en zoologie ne pouvaient pas comprendre différemment. Des traitements cladistiques plus récents de la phylogénie animale ont atténué ce biais, mais ne l'ont pas complètement éliminé. La phylogénie moléculaire a incité récemment à une réévaluation vigoureuse de ce scénario.
Phylogénie moléculaire
Evolution biochimique
La notion d’évolution biochimique vient maintenant épauler les zoologistes dans leurs travaux de classification. L’étude de l’évolution des macromolécules informationnelles est entreprise, et, malgré l’étendue des recherches a accomplir a déjà apporté de
précieux renseignements.
Ce genre d’étude repose sur l’universalité du code génétique et permet la comparaison de différents organismes quels que soient leurs degrés de spéciation.
Macromolécules informationnelles.
Les macromolécules informationnelles sont de grosses molécules formées de l’enchaînement
1) D’acides aminés que l’on appelle protéines , ces protéines pour être actives portent des sites actifs, ce sont les enzymes ou les hormones.
2) D’acides nucléiques que l’on appelle ADN et formant les gènes.
La structure des protéines permet par le biais du code génétique de déduire les séquences d’acides nucléiques correspondants.
Phylogénie moléculaire
La phylogénie moléculaire correspond à la phylogénie par comparaison de gènes.
Les gènes utilisés doivent être choisis avec soin , il faut que cela soit des gènes subissant de fortes contraintes fonctionnelles, donc ayant un taux de mutation faible.Un bon exemple concerne les cytochromes , par exemple le cytochrome b intervenant dans les chaînes d’oxydation cellulaire de tous les êtres vivants ( les êtres vivants actuels l’ont sans doute hérité d’un ancêtre commun il y a trois milliards d’années).
La comparaison des séquences d’un enzyme assurant une même fonction dans des organismes différents la fait apparaître des écarts plus ou moins importants selon que les organismes en question sont plus ou moins rapprochés phylogénétiquement.
Les premières études ont porté sur le cytochrome c transporteur d’électrons dans les chaînes respiratoires.
Si l’on compare le cytochrome de l’homme et celui de la drosophile, on constate que sur respectivement 104 et 107 motifs, 79 seulement coïncident.
Par contre, la comparaison des séquences homme singe rhésus ne se différencient que par trois mutations ponctuelles aux sites 24-29-69.
Cela signifie que ces deux espèces sont très proches l’une de l’autre et qu’elles possèdent un ancêtre commun direct. Il n’y a pas entre elles et l’ancêtre de ramifications vers l’une quelconque des espèces ayant d’autres séquences connues.
Cet ancêtre aura ce que l’on appelle une séquence nodale que devra en principe porter tous les motifs communs aux séquences filles sauf cas de mutation fortuite.
Pour les différences , reste à savoir si, par rapport à la séquence nodale , c’est celle de l’homme qui s’est trouvée modifiée ou celle du singe.
On constate que sur les sites 24 et 29 , les motifs de la séquence humaine sont les mêmes que pour l’ensemble des mammifères et pour la plupart des autres espèces . Par contre, sur le site 69 , la même situation apparaît en faveur du singe.
La séquence nodale sera donc la suivante : Motifs communs : sites 24 et 29 de l’homme, site 69 du singe rhésus.
De études similaires ont été entreprises sur des myoglobines et des hémoglobines, actuellement, on travaille sur des ARN ribosomiaux, sur du matériel génétique, sur des mitochondries, etc… et petit à petit se constituent des arbres phylogénétiques , ces travaux viendront, espère t’on étayer ou préciser la théorie de l’évolution.
l'horloge moléculaire
En résumé, on constate que le taux d'accumulation des mutations dans le génome d'organismes différents est du même ordre de grandeur dans des régions homologues (régions soumises à la même pression de sélection).
L'accumulation sera maximale pour des régions qui ne sont pas soumises à la pression de sélection naturelle (ne codant pas pour des gènes) et minimale dans les parties du génome soumises à une forte pression (c'est à dire les régions codant pour des fonctions essentielles à la survie de l'organisme).
Chaque séquence accumule les mutations à un rythme qui lui est propre et qui est dicté par l'intensité de la pression de sélection à laquelle elle est soumise. Pour reconstituer des phylogénies (dater la divergence entre deux espèces), on peut utiliser différentes molécules comme on utilise les aiguilles d'une montre pour calibrer l'horloge :
- la trotteuse des secondes (taux de mutation important, par exemple un pseudogène) pour des évènements récents (études des sous populations au sein d'une espèce).
- l'aiguille des minutes (taux de mutation moyen, par exemple le cytochrome C) pour l'analyse d'un passé proche.
- l'aiguille des heures (taux de mutations faible : les histones) pour l'étude d'un passé lointain.
La vitesse d'évolution de la séquence est du même ordre de grandeur au sein d'une même classe fonctionnelle de protéines et elle est différente pour des protéines qui ont des fonctions différentes : la vitesse d'évolution de la sérum albumine est toujours plus importante que celle du cytochrome C. Ces différences de vitesse dépendent à la fois de la probabilité qu'une substitution apparaisse et de sa compatibilité avec la survie de l'organisme. Si l'on admet cette théorie, et que l'on connaît le taux d'accumulation des mutations, il est possible d'estimer le temps de divergences d'espèces en comparant leur diversité moléculaire.
Arguments contre l'horloge moléculaire
La théorie de l'horloge moléculaire est remise en cause et plusieurs arguments ont été développés :
- L'horloge moléculaire ne serait pas constante (Goodman): les mutations avantageuses se fixeraient plus rapidement lors de la formation de nouvelles espèces.
- L'horloge moléculaire serait épisodique (Gillepsie) et les mutations ne se produiraient pas de façon indépendante au cours de l'évolution: il y aurait des épisodes d'accumulation suivis d'arrêts évolutifs.
Conclusion
Bien que le débat persiste, il semble que l'horloge moléculaire fonctionne assez bien sur de longues périodes évolutives, pour des
gènes ayant un taux de mutation relativement faible où même si l'horloge ne bat pas très régulièrement, les ralentissements et les accélérations se compensent.Il faut également se méfier des estimations de temps de divergence basées sur un petit nombre de gènes.
Acquis de la phylogénie moléculaire (ARNr 18S)
Notons en premier lieu que la tendance actuelle ( non traditionnelle) tend à vouloir organiser un arbre pourvu de groupes d’animaux monophylétique ce qui amènerait à une simplification des extrémités
buissonnante. Cette démarche serait intéressante au niveau des Vertébrés, ou le groupe des poissons semble particulièrement polyphylétique.
---------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------
Quelques acquis de la phylogénie moléculaire :
- Les éponges ( Spongiaires,Porifères) fermement attachées à l’ensemble des Métazoaires.
- Confirmation du caractère monophylétique des coelomates stricts, qui ont ,comme attendu, les Plathelminthes acoelomates comme groupe sœur.
Quatre grandes Clades chez les coelomates : 1) les arthropodes 2) les protostomiens Spiralia coelomates 3) les échinodermes 4) les chordés (cordés).
Ce buisson des phyla coelomates pourrait être le reflet de l’explosion cambrienne.
La branche des deutérostomiens apparaît tout à fait solide et la résolution que l’on a pu obtenir à l’intérieur du groupe confirme les vues existantes depuis longtemps, par exemple les échinodermes apparaissent comme groupe frère des cordés.
La situation est plus confuse au niveau des protostomiens ( prostomiens, protérostomiens…).
Eclatement du groupe des Articulata ( Annélides + Arthropodes), les Annélides sont solidement reliés aux mollusques et à d’autre phyla protostomiens partageant le clivage spiral des œufs et la larve trochophore.
Confirmation du vaste groupe de Spiralia rebaptisé groupe des Eutrochozoaires
Les Némertes ne sont plus considérés comme des acoelomates, mais doivent faire partie des Eutrochozoaires ( leur rynchocoele est un coelome ).
Création du groupe monophylétique des Lophotrochozoaires, confirmant l’appartenance des trois phyla de Lophophoriens ( Brachiopodes, Bryozoaires, Phoronidiens) au groupe des protostomiens avec les Trochozoaires ( Annélides et Mollusques).
L’arbre commence à se simplifier avec deux grands groupes monophylétiques : les protostomiens et les deutérostomiens.
Le problème des pseudocoelomates reste épineux, il semble que ceux-ci ne constituent pas un groupe naturel mais est constitué de groupes indépendants dont certains aboutissent aux arthropodes et d’autres aux nouveaux Lophotrochozoaires ( cas des nématomorphes et des priapuliens qui émergeraient avec les arthropodes).
Création du groupe des Ecdyzoaires comportant les nématodes et les arthropodes.
Les Plathelminthes seraient très proches des Lophotrochozoaires, ce qui annoncerait la fin des groupes coelomates et acoelomates comme degrés précoces des métazoaires précédent la séparation prostomiens deutérostomiens.
Annélides et mollusques n’apparaissent pas comme groupes monophylétiques.
La phylogénie intra lophotrochozoaire et intraecdyzoaires reste incertaine.
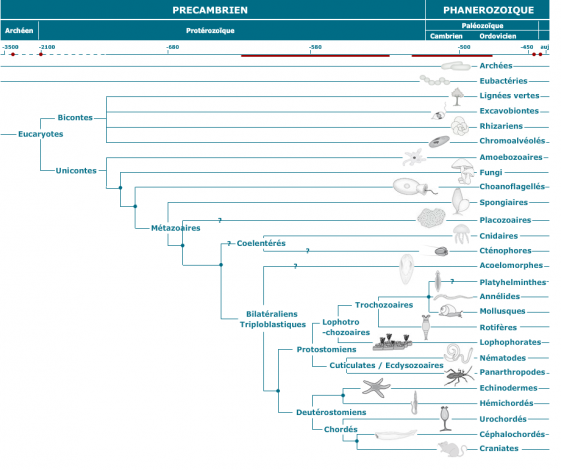
Mise à jour 18/10/2012 : modification de la classification
--------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------
DIPLOBLASTIQUES
Les éponges : de vrais Métazoaires ------> Spongiaires, Cnidaires, Cténaires
TRIPLOBLASTIQUES
1. Le groupe des Acoelomates ( Plathelminthes..) = groupe soeur des Coelomates .
2. Le groupe des Coelomates stricts est monophylétique
+ Coelomates stricts <------------Explosion cambrienne
+ Coelomates = Arthropodes, Echinodermes, Chordés, Potostomiens coelomates ( annélides, mollusques ........),les protostomiens comprenant des acoelomates, des pseudocoelomates et des coelomates est un groupe complexe. Ils sont actuellement considérés comme tous coelomates : Coelomates protostomiens est un groupe complexe
+ Coelomates deutérostomiens est un groupe solide ( echinodermes, chordés)
- Plus de groupe d'Articulata ( Annélides et Arthropodes )mais un groupe Eutrochozoaires de clivage spiral de l'oeuf ( Annélides, Mollusques ....), les Némertes ( leur rhyncocoele est un celome) ne sont plus des acoelomates mais font partie de ce groupe des eutrochozoaires -----------> ( Annélides, Mollusques, Némertes )
- Création du groupe des Ecdyzoaires ( Arthropodes et Nématodes )
- Création du groupe des Lophotrochozoiares , Eutrochozoaires + Groupe unifié monophylétique des Lophophoriens ( Brachiopodes, Bryozoaires, Phoronidiens ) .
- Les Plathelminthes seraient des Lophotrochozoaires, donc coelomates protostomiens ------------> plus de distinction coelomates/acoelomates mais une seul groupe des Coelomates.
3. Les pseudocoelomates ne constituent pas un groupe naturel mais est constitué de groupes indépendants dont certains aboutissent aux arthropodes et d’autres aux nouveaux Lophotrochozoaires ( cas des nématomorphes et des priapuliens qui émergeraient avec les arthropodes).
Mise à jour de 25/10/2013 : synthèse cette nouvelle classification est maintenant totalement acceptée  zoo-cours-oviv-2010-zoologie-n-2.pdf
zoo-cours-oviv-2010-zoologie-n-2.pdf
Effacement de la différence coelomate acoelomates, les némertes sont des coelomates ( Rhyncocoele) , les plathelminthes sont des lophotrochozoaires donc coelomates. Les pseudocoelomates ne constituent pas un groupe naturel mais est constitué de groupes indépendants dont certains aboutissent aux arthropodes et d’autres aux nouveaux Lophotrochozoaires ( cas des nématomorphes et des priapuliens qui émergeraient avec les arthropodes).
Ajout 16/06/2015 La nouvelle classification des Protostomiens
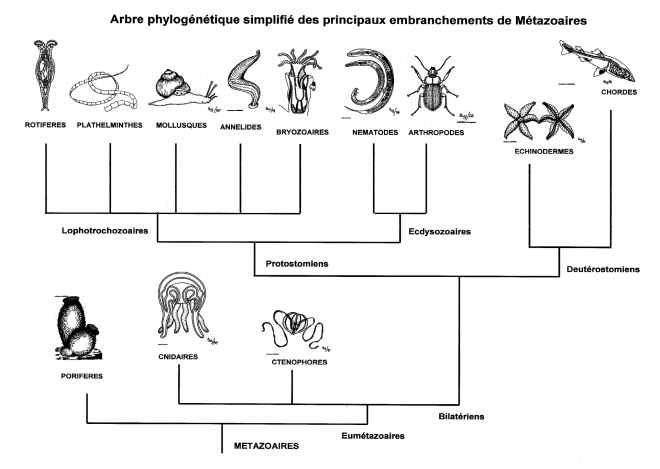
Les animaux bilatériens se divisent en deux groupes : les Protostomiens et les Deutérostomiens. Les Protostomiens sont caractérisés par :
Ø la bouche formée à partir du blastopore,
Ø le coelome schizocoelique,
Ø le système nerveux ventral,
Ø le squelette externe (à l’exception du squelette hydrostatique).
Selon la classification traditionnelle fondée sur la présence ou l’absence du coelome, on divise les Protostomiens en Acoelomates, Pseudocoelomates et Eucoelomates. Au cours de ces dix dernières années, les études moléculaires, basées principalement sur l’analyse des séquences d’ARN ribosomique et les caractéristiques de certains gènes du développement (Hox), ont démontré que cette classification ne reflète pas les relations réelles existant entre les différents phyla (embranchements) des bilatériens. Les données moléculaires ont montré que les
Acoelomates, les Pseudocoelomates et les Eucoelomates n’étaient pas des taxa valables. Le groupe des lophophoriens (ectoproctes, branchiopodes et phoronidiens) a été rapproché des mollusques et des annélides, en créant le groupe des Lophotrochozoa. Les études moléculaires ont montré également que les nématodes et certains autres pseudocoelomates sont proches des arthropodes, et clairement séparés des mollusques et des annélides. Le terme Ecdysozoa fut crée pour ce groupe ayant comme principal point commun la présence d’une cuticule plus ou moins rigide rendant obligatoire le phénomène de mue au cours de la croissance (ecdysis =mue). Les Protostomiens sont donc divisés en deux grands groupes : les Lophotrochozoa et les Ecdysozoa.
I. PROTOZOAIRES
II METAZOAIRES
II.1 PRIMITIFS
Spongiaires ( lien interne vers spongiares et cnidaires )
II.2 EUMETAZOAIRES
II.2.1 Non bilatériens
Cnidaires ( lien interne vers spongiaires et cnidaires )
II.2.2 bilatériens protostomiens
Les Lophotrochozoa regroupent les animaux possédant un lophophore (couronne de tentacules entourant la bouche) ou une larve de type trochophore. 13 phyla y sont inclus :
2. Annelida
3. Briachopoda
4. Cycliophora
5. Ectoprocta
6. Entoprocta
7. Gnathostomulida
8. Mollusca
9. Nemertini
10. Phoronida
11. Platyhelminthes
12. Rotifera
13. Sipuncula
Les Ecdysozoa sont caractérisés par (1) la perte des cils locomoteurs des cellules de l’épiderme, (2) la cuticule formée de trois couches, et (3) la croissance par des mues sous le
contrôle d’hormones ecdystéroïdes. Ils regroupent 9 phyla :
1. Arthropoda et insectes en particulier
2. Gastrotricha
Les pages liées suivantes sont en travail
3. Kinorhyncha
4. Loricifera
5. Nematoda
6. Nematomorpha
7. Onychophora
8. Priapula
9. Tardigrada
II.2.3 Bilatérien deutérostomiens
Dans ce tableau de liens, je n'ai plus tenu compte de la différenciation entre coelomates, acoelomates et pseudocoelomates, rendue obsolète par les dernières recherches.
Conclusions
1) fin des Acoelomates, des pseudocoelomates : tous les triploblastiques sont des coelomates, Acoelomates et pseudocoelomates sont intégrés dans les protostomiens2) éclatement des protostomiens : les Lophotrochozoaires et les Ecdysozoaires.3) Eloignement des Annélides et des Arthropodesles Annélides sont proche des Mollusquesles Arthropodes proche des NématodesIndépendamment de la paléontologie, les données moléculaires obtenues récemment à partir du génome d'espèces actuelles (ex: le gène 18S ARN) ont permis de construire de nouveaux schémas de parenté entre phylums (phylogénie moléculaire) et de proposer une chronologie de divergence entre groupes. Selon ces méthodes, trois groupes majeurs, les ecdysozoaires (animaux à croissance par mues regroupant les arthropodes, tardigrades, onychophores, priapuliens et nématodes), les lophotrochozoaires (ex: mollusques et brachiopodes) et les deutérostomiens (ex: échinodermes, chordés) émergent d'une souche pré-bilatérienne (ex: éponges, cténophores, cnidaires) 22). La phylogénie moléculaire est globalement en accord avec la paléontologie concernant l'ordre d'apparition des phylums et le concept général d'Explosion Cambrienne. Cependant, les données moléculaires fixent des âges de divergences entre groupes souvent très antérieurs à l'Explosion Cambrienne(52-55), les estimations variant parfois de 800 millions d'années suivant les techniques utilisées (ex: la divergence protostomiens-deutérostomiens datée alternativement à 1200 MA et 670 MA). Outre certains problèmes méthodologiques (ex: taux de substitution variables suivant les taxons), le désaccord entre données paléontologiques et moléculaires vient probablement d'un découplage entre divergence moléculaire et acquisition des plans anatomiques.Remise en cause de l’origine annélidienne des Arthropodes : les mécanismes génétiques de mise en place de la segmentation sont différents
Metazoaires BILATERIENS PROTOSTOMIENS
I. .LOPHOTROCHOZOAIRES
A Clade Trochozoa (Trochozoaires )
A.1 Clade Phoronozoa ou Brachiozoa
- Embranchement Brachiopoda ( Brachiopodes )
- Embranchement Phoronida ( Phoronidiens )
A.2 Clade Eutrochozoa
----------------------------------------------------------------------------------------------------------------------------------
Les Eutrochozoaires (Eutrochozoa) sont un sous-clade putatif des Trochozoaires, parmi les Lophotrochozoaires.
Conçu à l'origine comme regroupant :
-les Mollusques,
-Annélides,
-Siponcles et
-Brachiopodes,
Il a été corrigé en 1998 comme regroupant:
-les Mollusques,
-Annélides,
-Siponcles (Sipunculiens) et
-Némertiens.
---------------------------------------------------------------------------------------------------------
Ce groupe peut encore précisément être divisé en :
- Syndermates
Embranchement Rotifères
Embranchement Acanthocéphales
- cycliophores
certains crustacés
- Spiraliens divisés en
Parenchymiens : Embranchement des Plathelminthes et Embranchement des Némertes ,
Embranchement Entoproctes,
Embranchement Mollusques,
Embranchement Sipunculiens
Embranchement Annélides
B.Clade Lophophorates ( Lophophoriens)
On y dénombre trois embranchements, tous considérés aujourd'hui comme protostomiens :
B.1.Les Phoronides, caractérisés par un quatrième feuillet : l'épistosome qui vient au-dessus du prosome. Ils vivent fixés sur le substrat, possèdent un tube digestif entouré par une membrane et s'enroulent dans de la chitine à la manière des arthropodes (ex. Phoronopsis hippocrepia).
B.2Les Brachiopodes, phylum ressemblant au groupe des lamellibranches (mollusques), ils se distinguent par leurs coquilles ventrale et dorsale et par les muscles qui les actionnent (ex : Laqueus).
B.3Les Ectoproctes (bryozoaires s.s.) caractérisés par un squelette externe (zoïde) où l'animal (le polypide) se cache dans une loge (cystide), ce squelette est une matière calco-chitineuse sécrétée par l'animal. Ils se développent en colonie, se reproduisent par bourgeonnement et ressemblent aux coraux (ex: Electra pilosa).
Certains pensent que le groupement est polyphylétique. Peterson & Eernisse (2001), réservent ce terme aux deux premiers phyla, qui constituent un super-embranchement plus connu sous le nom de Phoronozoa.
2.ECDYZOAIRES
Les Ecdysozoaires (Ecdysozoa) sont un clade d'animaux représentant la division des protostomiens dont le développement s'effectue par une ou plusieurs mues cuticulaires. Le nom de ce taxon vient du mot « ecdysis », qui désigne la mue.
Les groupes les plus connus chez les ecdysozoaires sont les arthropodes (comprenant notamment les insectes et les crustacés) et les nématodes.
Ce clade rend obsolète l'ancienne notion d'articulés (en) (classification de Georges Cuvier en 1817), qui regroupait arthropodes et annélides sur l'idée que la segmentation était un caractère ancestral. Les données génétiques montrent que la segmentation s'est développée de façon indépendante chez les ecdysozoaires et les lophotrochozoaires. C'est bien la mue qui constitue un caractère ancestral et justifie de regrouper des ecdysozoaires.
La mue des arthropodes est déclenchée par l'ecdysone, mais cette hormone n'est probablement pas présente chez tous les Ecdysozoaires : le nématode Caenorhabditis elegans utilise une autre hormone stéroïde.
Parmi les autres synapomorphies des ecdysozoaires, on peut mentionner :
- le pharynx à musculature triradiée,
- une cuticule composée de 3 couches : l'épicuticule, l'exocuticule (composée de chitine) et l'endocuticule,
- la perte des cils locomoteurs de l’épiderme.
Embranchements
Arthropoda Latreille, 1829
Kinorhyncha Reinhard, 1881
Loricifera Kristensen, 1983
Nematoda Diesing, 1861
Nematomorpha Vejdovsky, 1886
Onychophora Grube, 1853
Priapulida Delage & Hérouard, 1897
Tardigrada Spallanzani, 1777
Notons le cas incertain des Gastrotriches classés parmi les Ecdyzoaires par Le Guyadier-Lecointre . Des études plus récentes les classent parmi les Eutrochozoaires Spiraliens
SYNTHESE DE LA CLASSIFICATION MODERNE
Date de dernière mise à jour : 06/01/2019
Commentaires
-
 I definitely wanted to post a small message so as to express gratitude to you for some of the amazing tips you are placing on this site. My extensive internet investigation has at the end of the day been paid with incredibly good details to talk about with my relatives. I would declare that we website visitors actually are truly fortunate to dwell in a fabulous site with so many outstanding individuals with useful things. I feel rather fortunate to have come across your webpages and look forward to some more amazing moments reading here. Thank you again for all the details.
I definitely wanted to post a small message so as to express gratitude to you for some of the amazing tips you are placing on this site. My extensive internet investigation has at the end of the day been paid with incredibly good details to talk about with my relatives. I would declare that we website visitors actually are truly fortunate to dwell in a fabulous site with so many outstanding individuals with useful things. I feel rather fortunate to have come across your webpages and look forward to some more amazing moments reading here. Thank you again for all the details. -
 Merci pour vos aide a travers cet site
Merci pour vos aide a travers cet site -
 Tous les remerciements pour le contenu de ce document
Tous les remerciements pour le contenu de ce document -
 c'est formidable
c'est formidable
Ajouter un commentaire