
Loph/Ectoproctes
INTRODUCTION : EMBRANCHEMENT DES LOPHOPHORIENS
DEFINITION ET SUBDIVISIONS
L'EMBRANCHEMENT DES LOPHOPHORIENS comporte trois catégories de Coelomates protostomiens ( traditionnellement quatre avec les Endoproctes ou Kamptozoaires ) auxquels certains auteurs attribuent la valeur d'Embranchement donc de phylums distincts : les Ectotropes ou Bryozoaires, les Phoronidiens et les Brachiopodes.Les représentants de ces divers groupes sont caractérisés par la présence autour de la bouche de tentacules, disposés en couronne ou en U , couverts de cils, donc les battements déterminent un courant d'eau portant les particules alimentaires vers la bouche ; ce dispositif ou lophophore est constituéde tentacules creux, simples évaginations de la paroi du corps contenant un diverticule coelomique La nutrition de ces organismes est du type muco-ciliaire. Chez les Lophophoriens l'axe antéro-postérieur est très raccouci par suite du rapprochementde la bouche et de l'anus; le tube digestif , comme celui des Sipunculiens, forme une anse digestive étendue dans une longue dilatation du corps qui représente le métasome ; l'anus est dorsal et presque toujours en dehors de l'aire lophophorienne L'appareil excréteur, lorsqu'il existe est composé de métanéphridies
Ces organismes sédentaires, généralement fixés, parfois bourgeonnant donnent naissance à des larves libres, ciliées, planctoniques qui par certains caractères rappellent la larve trochophore des Annélides, ce qui témoignerait d'une parenté entre ces embranchements. De plus, les Lophophoriens présentent par les caractères de leur coelome des affinités avec les Deutérostomiens. En effet, comme chez de nombreux Deutérostomes tels que les Hémichordés et les Echinodermes, le corps est divisé, au moins au stade larvaire, en 3 régions; le protosome, le mésosome et le métasome, et chaque région contient une cavité coelomique : protocoele, mésocoele et métacoele. Quoique le protocoele ne soit pas très développé chez les Lophophoriens par suite peut-être d'une réduction de la tête, le coelome est nettement divisé par un septum transverse , un mésocoele et un métacoele; il est parfois difficile d'affirmer que le coelome est typiquement trimère. Le mésocoele est situé dans la région antérieure du corps et se prolonge dans les tentacules; le métacoele occupe la partie troncale du corps.
D'un point de vue phylogénétique l'embryologie des Lophophoriens est particulièrement intéressante car des modalités de type protostomien et deutérostomien peuvent être décelées. Chez les Phoronidiens et les Brachiopodes la bouche dérive du blastopore et la larve est une trochophore modifiée; ces groupes sont donc de véritables protostomiens mais la segmentation n'est véritablement spirale que chez les Phoronidiens. Chez les Ectotropes et surtout les Brachiopodes le clivage est radiaire et plus, chez quelques Brachiopodes , le mésoderme et le coelome se forment par entérocoelie comme chez de véritables deutérostomiens .
En plus de l'existence d'un lophophore, d'un coelome compartimenté, presque tous les membres de ce groupe sont sessiles, ont une tête réduite et secrètent une enveloppe protectrice.
Selon Hyman les Lophophoriens seraient issus d'une forme ancestrale commune, détachée du tronc conduisant aux Annélides, et qui aurait également donné naissance à la lignée des deutérostomiens. Ainsi les Lophophoriens représentent peut-être un groupe charnière à partir duquel les 2 tendances évolutives principales de Règne animal auraient divergé.
12 Ectoproctes
I. CLASSE DES ECTOPROCTES OU BRYOZOAIRES
Triploblastiques coelomates bilatérien hyponeuriens protostomiens Lophotrochozoaires Lophophorates
Définition et caracteres généraux
Les Ectoproctes ou Bryozoaires sont des organismes métazoaires triploblastiques, coelomates, protostomiens, non segmentés dotés d'une symétrie bilatérale . Ces animaux microscopiques ( 1 mm environ ), aquatiques ( marins ou dulcicoles ),généralement sédentaires sont doués d'un intense pouvoir de bourgeonnement et constituent des colonies qui prennent souvent l'aspect de Mousses ce qui leur a valu l'appellation de Bryozoaires.
Chez ces Lophophoriens qui comptent environ 4000 espèces le cercle tentaculaire ou lophophore entoure uniquement la bouche alors que l'anus est situé à l'extérieurdu cercle. Ce sont donc des Ectoproctes autrefois réunis au Endoproctes dans le groupe des Bryozoaires maintenant démembré. Ils se distinguent nettement des Endoproctes par leurs tentacules lophophoriens qui peuvent se rétracter dans une gaine tentaculaire, l'absence de néhridies et d'appareil circulatoire, la production de larves ciliées, libres, pélagiques, leur reproduction asexuée par bourgeonnement. Ce sont des organismes microphages dont le tube digestif décrit une boucle. Leur degré d'organisation est supérieur à celui des Endoproctes.
Les Ectoproctes sont répandus dans toutes les mers ; ils prolifèrent dans la zone littorale et adhèrent à tous les substrats ; algues, rochers, coquilles, autres organismes. Ils forment des colonies étalées, encroûtantes ou dressées, arborescentes, foliacées, rigides ou gélatineuses. Ces colonies ne dépassent guère en général quelques centimètres ; exceptionnellement elles sont de grande taille, par exemple chez les Eschara de notre littoral.
Les Bryozoaires constituent un des groupes dominants de la faune fixée des fonds rocheux ; ils s'installent partout où un support solide le permet. A cet égard ils jouent dans les peuplements des mers un rôle équivalent à celui des Spongiaires, des Cnidaires , des Ascidies. Ils sont connus à l'état fossile depuis l'ère primaire.
II. ORGANISATION GENERALE
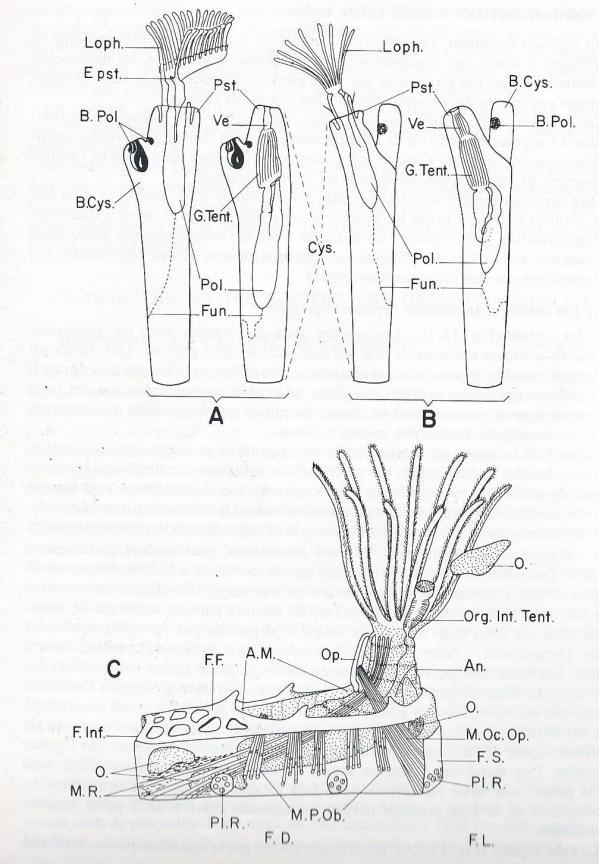
Un individu fondateur l'oozoïte ( ou oozoïde ) est là l'origine de toute la colonie de Bryozoaires. Il donne par bourgeonnement 1 ou plusieurs individus, les blastozoïtes qui restent unis les uns aux autres par leurs parois contigües. Par la suite ils bourgeonnent eux-même de nouveaux blastozoïtes et ainsi de suite.
Les diverses individualités ( oozoïte, blastozoïtes et 1er, 2ème , nième génération ) reçoivent l'appellation générale de zoÏtes ou zoécies dont la constitution fondamentale est identique et complète. Chaque zoécie comporte une loge ou cystide et l'animal proprement dit ou polypide. La colonie est appelée zoarium.
Chez certaines espèces, tous les individus d'une colonie sont identiques alors que chez d'autres espèces il existe un polymorphisme plus ou moins marqué traduisant une spécialité fonctionnelle ; on distingue alors des autozoécies ( ou autozoïtes ) comparable au zoïtes des colonies sans polymorphisme et des hétérozoïtes ( ou hétérozoécies ) qui s'en distinguent nettement.
A. Les zoécies et autozoécies : cystide et polypide.
1. La cystide.
Les cystides sont des logettes dont les dimensions varient de quelques dixièmes de mm à 3 mm pour les plus grandes. Leur forme est également variable mais le moulage réciproque des différents individus leur donne le plus souvant une forme polyédrique. Dans les espèces encroûtantes une des faces adhère au support, une autre est au contact du milieu extérieur, tandis que les autres faces sont contigües à celles des zoécies voisines.
La paroi de la logette est constituée par une cuticule ou ectocyste continue, parfois calcifiée, doublée intérieurement par un épithélium mésodermique pariéto-pleural. Ces deux épithéliums sont séparés par une couche de fibres musculaires longitudinales dont la contraction, par augmentation de la pression des liquides internes, assure la dévagination de la gaine tentaculaire.
2. Le polypide.
Suspendu dans la cavité cystidienne, peut en faire saillie partiellement à l'extérieur au niveau d'un orifice circulaire visible à la face externe de la logette ( orifice cystidien ). Son organisation est très simple . En effet, il est constitué par une anse digestive dont l'orifice buccal est entouré par une couronne de tentacules ciliés sur leurs faces internes et latérales , supportés par une crête également ciliée. Les tentacules peuvent se rétracter dans une gaine ( gaine tentaculaire ) qui s'invagine en doigt de gant à partir de l'orifice cystidien et le prolonge à l'extérieur lorsqu'elle est dévaginée . La gaine tentaculaire limite un espace atrial ou atrium ; elle est surmontée d'une paroi vestibulaire également invaginée, circonscrivant un vestibule séparé de l'atrium par un diaphragmme et s'ouvrant à l'extérieur par l'orifice cystidien. Une membrane circulaire souple forme une sorte de colerette à la base de la gaine ; elle ferme l'ouverture de la loge lorsque les tentacules sont rétractés. L'ouverture de la loge présente parfois un opercule pourvu de 2 petits muscles adducteurs.
Le tube digestif , entièrement revêtu par la sphanchnopleure, est diviséen 3 régions par deux constrictions transversales. Le pharynx est cilié dans sa partie antérieure. L'estomac encadré par la valvule cardiaque et la valvule pylorique est composé d'un cardia terminant la branche descendante et d'un pylore amorçant la branche ascendante ; il est en relation avec le diverticule caecal ou caecum stomacal. La région postérieure correspond à l'intestin et au rectum qui aboutit à l'anus. L'intestin présente des rangées ciliaires.
La digestion est extracellulaire. Les courants d'eau créés par les battements des cils tentaculaires entraînent les particules alimentaires ( microphagie vers la bouche et la partie ciliée du pharynx . Ses particules sont littéralement dégluties par la partie postérieure non ciliée du pharynx qui présente des mouvements de dilatation et de contraction réguliers
La cavité générale des Ectoproctes est un véritable coelome dont la partie antérieure séparée par un véritable septum constitue une sorte de canal annulaire qui se prolonge dans les tentacules ( coelome annulaire lophophorien). Cette cavité remplie de liquide lymphatique contient des cellules amoebocytaires et des leucocytes à inclusion; elle ne contient aucun tissus conjonctif.
Le funicule . L'anse intestinale est reliée à la paroi interne de la logette par une lame plus ou moins épaisse , située dans le plan sagittal, le funicule constitué par une assise mésodermique en continuité avec celle qui double la paroi de la zoécie et le tube digestif. Le funicule relie la splanchnopleure à la somatopleure.
La musculature formée exclusivement de fibres lisses est représentée par des muscles rétracteurs longitudinaux et des muscles pariétaux longitudinaux dont l'activité assure la dévagination de la partie antérieure du polypide. La dévagination est lente, l'invagination est rapide. Autour du tube digestif les fibres musculaires surtout transversales , quelquefois longitudinales , assurent les mouvements péristaltiques.

Le système nerveux : comporte un nganglion cérébroïde appliqué contre la paroi de la cavité buccale, sous le lophophore. Il en part un cordon péripharyngien, lophophoral, en connexion avec les nerfs qui desservent la paroi et les viscères. Il n'existe pas d'organe des sens spécialisé.
L'appareil génital est hermaphrodite le plus souvent. L'ovaire est inséré sur le funicule et les gonades mâles sont attachées ça et là sur les parois du corps. A maturité, les produits génitaux rompent les membranes enveloppantes des gonades et tombent dans la cavité générale.
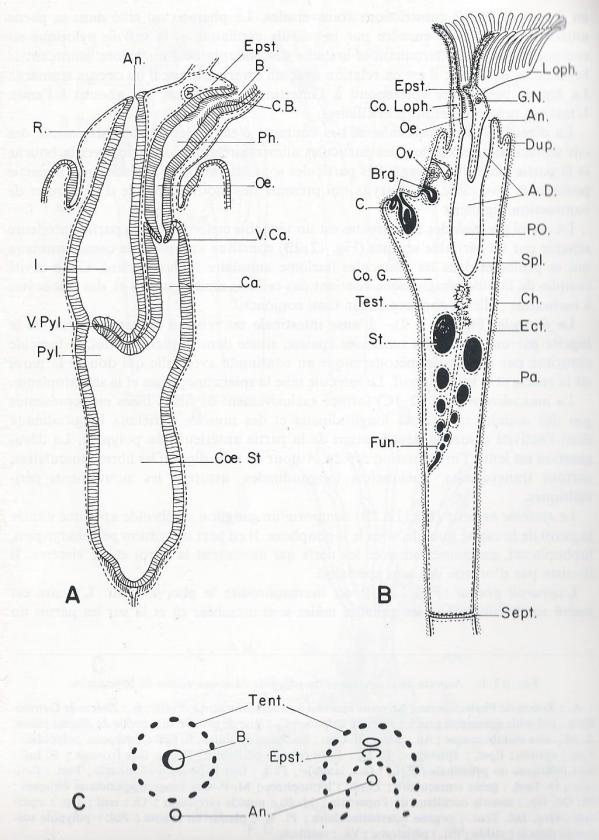

Il n'existe pas d'appareil circulatoire, ni d'appareil respiratoire . Les tentacules épanouis assument les échanges gazeux , il n'existe pas non plus d'appareil excréteur ; les cellules de la paroi intestinale se chargent de produits d'excrétion , constituant un rein d'accumulation. Cette surcharge entraîne la mort du polype et seule la logette demeure vivante ; le tube digestif, les muscles rétracteurs se "ratatinent" et forment un "corps brun" alors qu'un nouveau polypide se constitue à partir de la paroi de la logette. Le corps brun est généralement englobé par l'estomac du nouvel individu; il est digéré et ses résidus rejetés par l'anus .
A la vérité, cette rénovation du polype est un phénomène normal et périodique chez beaucoup d'Ectotropes et il est peu vraisemblable qu'il soit uniquement déterminé par l'accumulation des excréta puisque, en l'absence de néphridies, l'excrétion peut s'effectuer par diffusion vers l'extérieur, au travers des téguments.
Chez Alconydium et chez d'autres Ectoproctes, il apparaît au moment de la maturité sexuelle un organe intertentaculaire cilié qui met en communication le coelome antérieur et le milieu ambiant. Il est situé entre les te,ntacules , en arrière de la bouche et du ganglion nerveux et sert, chez certains, au rejet des ovules.
Dans beaucoup de formes marines ( Cténostomes, Chilostome ) les communications entre les individus de la colonie se font par l'intermédiaire de plaques perforées de petits orifices ou plaques en rosette. Ces plaques sont réunies, notamment chez les chilostomes, par des tractus mésenchymateux de nature funiculaire ou cordons latéraux.
En conclusion, la zoécie présente une symétrie bilatérale nette, Le plan sagittal passe par la bouche , l'anus, les deux branches de l'anse digestive , le ganglion cérébroïde et coupe le lophophore en deux moitiés symétriques ; il est donc possible de déterminer une face gauche et une face droite, une face ventrale correspondant à la bouche et une face dorsale correspondant à l'anus.
B. Dualité de la zoécie.
Pour des raisons d'ordre embryologique il est possible de reconnaître dans chaque zoécie 2 entités distinctes : la cystide formée de la paroi zoéciale avec sa doublure extomésodermique, le polypide constitué par l'anse digestive et la gaine tentaculaire. Cette notion est justifiée par le fait que lors de la formation et du développement des colonies cystide et polypide ont une évolution pratiquement indépendante. La partie polypidienne de la zoécie peut dégénérer, disparaître et être remplacée par un nouveau polypide sans altérer l'individualité de la cystide.
III.PARTICULARITE BIOLOGIQUE : LE BOURGEONNEMENT
Le bourgeonnement se fait en deux temps . Le premier temps consiste en la formation de la nouvelle cystide ; c'est une excroissance en un certain point de la paroi du cystide initial, le bourgeon cystidal. Cette hernie cystidale , à un moment de sa croissance édifie un nouveau polypide par prolifération de sa paroi ventrale ou frontale ; le bourgeon polypidien aboutit à la constitution des organes fondamentaux : anse digestive, lophophore, ganglion cérébroïde, système nerveux . La nouvelle cystide reste attachée à la cystide qui l'engendre , ect. Le bourgeonnement assure la formation de la colonie dont la croissance est continue par ses bords et ses rameaux les plus distaux. Dans chaque groupe le bourgeonnement colonial abéit à des lois précises.
IV SYSTEMATIQUE
Nous ne nous étendrons pas sur la diversité des Ectoproctes ( Bryozoaires ) qui font, je le rappelle partie de l'Embranchement des Lophophoriens avec les Brachiopodes et les Phoronidiens.
En fait, ils se divisent en deux groupes :
- Les phylactolèmes.
- Les Gymnolèmes.
Les Phylactolèmes sont exclusivement dulcicoles et forment des colonies ramifiées ou massives, fixées aux rives ou sur les plantes aquatiques des ruisseaux ombragés à eau riche et courante ( ex Paludicella ) . Ils sont cosmopolites et comptent un nombre limité d'espèces ( 8 à 10 en France ). Les polypes d'une même colonie , plus grands que ceux des Gymnolèmes, sont plongés dans une sorte de coelome commun par suite de séparation incomplète des zoécies formées par bourgeonnement. Les tentacules, réunis à la base par une membrane continue ( membrane intertentaculaire ), sont disposés en fer à cheval sauf chez Fredericella dont le lophophore est circulaire. Le lophophore entoure la bouche qui peut être obturée par une petite lèvre ou épistome mais il n'existe pas d'opercule . Les deux branches du " fer à cheval " , dirigées dorsalement , encadrent l'anus. Cette disposition en fer à cheval évoque celle du lophophore de Phoronis. Les cystides sont longues, tubuleuses , cylindriques et naissent les unes des autres par bourgeonnement ; elles sont en principe séparées par des septa mais le plus souvent ces cloisons n'existent pas ( Plumatella ) . Ces cystides représentent les rameaux de colonies puissantes , mais il arrive que les cystides se racourcissent , confluent et constituent des colonies globuleuses où seuls les polypides restent distincts ( Lophopus ) ; la colonie acquiert alors une véritable individualité et une remarquable ordonnance des polypides, elle est libre et glisse sur le substrat.
- Les Gymnolèmes sont essentiellement marins mais quelques espèces vivent en eau saumâtre ( Victorella) ou sont devenues dulcicoles ( Paludicella ).Les tentacules lophophoriens ( 8 à 34 ) sont indépendants les uns des autres et disposés en cercle périoral . La zoécie a la structure fondamentale décrite ci-dessus mais il n'existe pas d'épistome . L'anse digestive est parfois compliquée par suite d'une dilatation de la région cardiaque en un volumineux jabot ou en un gésier musculeux dont les cellules surélevées et coiffées de formations cuticulaires constituent des dents masticatrices . L'ectocyste cuticulaire sécrété par l'ectoderme de la cystide se renforce par calcification ; sa paroi est alors rigide et souvent ornée de ciselures et d'épines. La musculature pariétale est alors inefficace quant à la dévagination du polypide et il se différencie un sac compensateur hydrostatique qui, en se remplissant d'eau, exercera une pression sur le polypide et le forcera à se dévaginer . La cystide est primitivement tubuleuse et cylindrique ; l'orifice cystidien est apical . Généralement elle se raccourci et prend une forme ovoïde ou même celle d'un petit coffret parallélépipédique . Dans ce cas , les cystides sont étroitement accolées dans la colonie comme des dalles juxtaposées ; l'orifice cystidien est alors reporté sur la face ventrale ou frontale. A son niveau, un repli de la paroi frontale forme un clapet ou opercule remarquablement ajusté à l'orifice cystidien ; cet opercule en basculant sur sa base s'ouvre alors que le polypide évagine son lophophore et se referme lorsque le polypide rentre dans sa cystide . Cet opercule est mû par des muscles occulteurs issus des muscles vestibulaires et diaphragmatiques ( Chilostomes ) . Le funicule des zoécies des Gymnolèmes s'attache par son extrémité distale à la paroi dorsale et non à la paroi ventrale comme chez les Phylactolèmes.
On différencie encore au sein des Gymnolèmes 3 groupes :
1) Les cténostomes
2) Les Chilostomes ---------------1)+2) = Eurystomes.
3) Les cyclostomes
Date de dernière mise à jour : 24/08/2018
Ajouter un commentaire


