
Physiologie animale 6
VI CELLULES SECRETRICES ET SECRETION
1. L'Activité sécrétoire
1.1 Définition
L'activité sécrétoire d'une cellule est un processus par lequel elle élabore un ou des produits spécifiques, ensuite exporté(s) vers d'autres cellules ou vers l'extérieur de l'organisme . La sécrétion est très répandue dans l'organisme et elle est apparue très précocement lors de l'évolution des espèces : les Spongiaires possèdent en effet des cellules spécialisées dans la sécrétion de mucus , à usage collectif, et d'autres cellules, les spongioblastes , qui élaborent la spongine, une substance apparentée au glycogène.
1.2 Importance quantitative de la sécrétion
Ce critère doit retenir l'attention car il permettra de rattacher ou non une activité cellulaire à la sécrétion de type glandulaire. Certaines cellules ont une activité sécrétoire très développée : dans l'espèce humaine, les sécrétions du pancréas exocrine ou des glandes salivaires se chiffrent en litres de liquide et en grammes de protéines par 24 heures ; les glandes mammaires peuvent également produire des dizaines de grammes de protides, lipides et glucides par jour. Par contre les sécrétions de certaines cellules endocrines ou neuroendocrines sont parfois très peu abondantes , voire moins abondantes que la libération de certains neuromédiateurs par des neurones très sollicités.
La sécrétion ne semble pas être l'apanage d'une catégorie cellulaire particulière mais correspond à l'exagération des capacités de synthèse , et à la libération de substances qui sont destinées à la communauté cellulaire ou aux relations avec l'écosystème dont elle dépend.
1.3 Les processus sécrétoires
Le choix de quelques modèles de cellules sécrétrices envisagées ci-après a pour but d'illustrer les divers aspects de la sécrétion :
- phases de synthèse
- libération du produit
- couplage entre stimulus et sécrétion
- aspects électrophysiologiques ...
2. La cellule acineuse du pancréas exocrine
2.1 Le pancréas exocrine de Mammifère
2.1.1 Structure
Le pancréas est une glande digestive mixte, qui sécrète des hormones dans le sang ( composante endocrine ) et des sucs digestifs dans l'intestin ( composante exocrine ). La partie exocrine du pancréas est de loin la plus volumineuse : 99 % de la masse pancréatique chez le Rat. La partie endocrine est représentée par les îlots de Langerhans .
Le pancréas exocrine est une glande acineuse composée ; un réseau de canaux draîne la sécrétion de nombreux acini formant la glande.
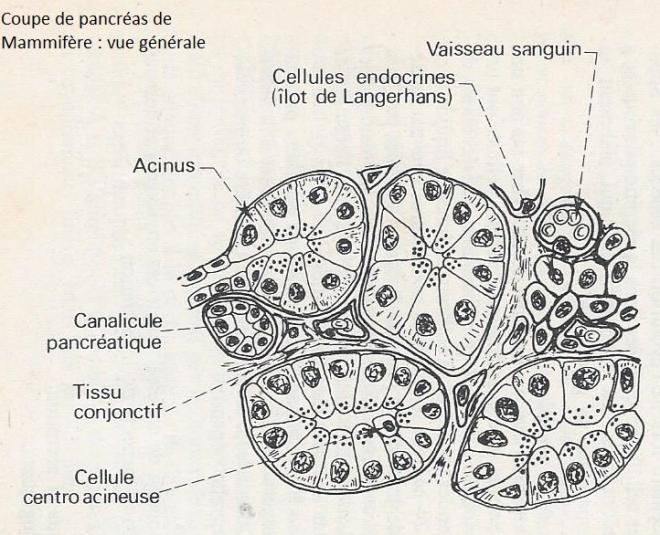
fig 1
Chaque acinus est formé par quelques dizaines de cellules de forme grossièrement pyramidale , groupées autour d'une cavité "virtuelle" ( elle est rarement visible sur les coupes de pancréas ) en ralation avec des canaux collecteurs.
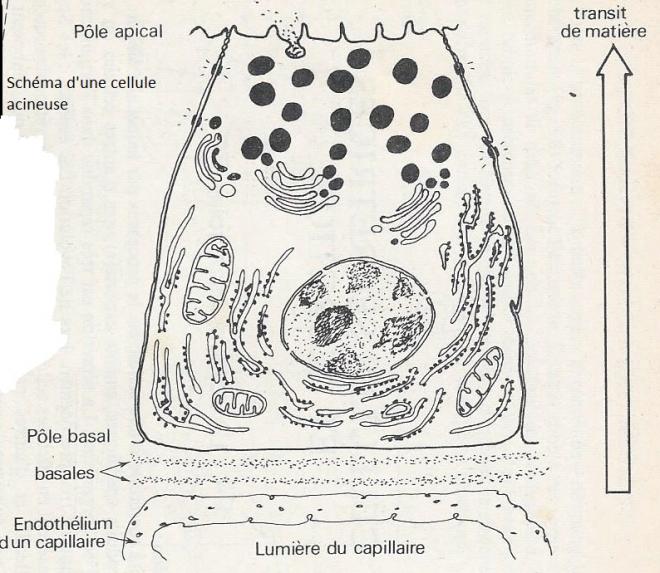
fig 2
2.1.2 Rappel physiologique
Les cellules acineuses du pancréas sécrètent diverses enzymes ou proenzymes ( précurseurs inactifs ) ayant un rôle digestif : trypsinigène, chymotrypsinogène, carboxypeptidase, ribonucléase, désoxyribonucléase, lipase, amylase ... Ces enzymes sont des protéines de masse moléculaire élevée. Les cellules des canaux pancréatiques, ainsi que les cellules centro-acineuses, sécrètent l'eau et les sels minéraux ( en particulier les bicarbonates ) qui forment la partie liquide de la sécrétion. Ces diverses sécrétions sont soumises à un contrôle multihormonal et nerveux dont la complexité commence seulement a être perçue.
2.1.3 Polarité cellulaire
Les cellules acineuses du pancréas ont une polarité extrêmement marquée sur les plans structural et physiologique. La partie de la cellule qui s'appuie sur la membrane avoisinant les capillaires , et par conséquent la région par où se font les apports nutritionnels, est appelé pôle basal, alors que la partie diamétralement opposée de la cellule, tournée vers la lumière de l'acinus forme le pôle apical.
La membrane orientée vers la lumière a une surface qui représente 1/20 de la surface membranaire totale d'une cellule . La polarité cellulaire est maintenue par les diverses formations membranaires qui unissent les cellules d'un même acinus.
A la partie basale de la cellules se trouvent le noyau, les mitochondries, le réticulum endoplasmique rugueux, dont les cavités sont larges et orientées parallèlement à l'axe apicobasal de la cellule. Le réticulum endoplasmique représente environ 60 du volume cytoplasmique et le contenu en ARN de chaque cellules est l'un des plus élevé de l'organisme , ce qui atteste d'une très grande activité de synthèse protéique. Au dessus du noyau, le complexe de Golgi est également très développé. Des vésicules contenant un matériel dense aux électrons semblent s'en détacher. Leur contenu est de plus en plus dense lorsque ces vésicules sont plus proches de la membrane apicale ; elles forment des graines de zymogène ( environ 3 du volume cytoplasmique contenant les enzymes digestives ).
2.1.4 Intérêt de ce type cellulaire
La cellule acineuse du pancréas présente plusieurs caractéristiques favorables à l'étude de la sécrétion.
- activité quantitativement très importante;
- polarité cellulaire marquée, ce qui facilite les études morphophysiologique : la ségrégation des produits de synthèse s'effectue de la base vers l'apex de la cellule.
- possibilité de déclencher l'activité des cellules de manière synchrone , après avoir provoqué leur déplétion par un stimulus approprié ( pilocarpine par exemple ): toutes les cellules libèrent leur contenu en même temps et un nouveau cycle sécrétoire peut commencer.
3. Le cycle sécrétoire
3.1 Etapes de la sécrétion
Lorsque la cellule acineuse passe de l'état de repos apparent à l'état sécrétoire le processus peut se décrire en le partageant en quaztre périodes, bien que les phénomènes se déroulent de manière continue :
- captation des acides aminés et début de synthèse protéique.
- ségrégation des protéines.
- stockage.
- extrusion des granules.
3.1.1 Captation des acides aminés et synthèse protéique
Les acides aminés présents dans les capillaires sont captés au pôle basal de la cellule acineuse. Pour cela ils doivent traverser successivement :
- L'endothélium vasculaire
- La lame basale du capillaire
- La lame basale de l'acinus
- La membrane basale de la cellule acineuse.
Le transfert des acides aminés ne demande toutefois que quelques secondes par suite de l'existence d'un transport par transporteur très efficace.
La synthèse des protéines débute immédiatemment au niveau des ribosomes, groupés en polysomes ( ou polyribosomes ) associés aux membranes du réticulum endoplasmique.
3.1.2 Ségrégation des protéines
Les diverses protéines enzymatiques synthétisées au niveau des polysomes sont immédiatemment ségrégées vers l'intérieur des saccules du réticulum endoplasmique . Cette ségrégation présente un intérêt pour la cellule, dans la mesure où les produits élaborés sont des enzymes ou des proenzymes susceptibles de pouvoir interagir avec certains éléments cytoplasmiques. Les protéines sont ensuite transportées, par les saccules du réticulum, vers l'appareil de Golgi. Le mécanisme par lequel s'effectue le transit des protéines depuis le réticulum vers les cavités des divers dictyosomes du système de Golgi est encore mal connu. Il se produirait un bourgeonnement de petites vésicules issues du réticulum, qui fusionnerait alors avec la face convexe des dictyosomes. La phase de ségrégation des protéines, jusqu'à l'appareil de Golgi, s'effectue dans les 15 à 20 min suivant le début d'incorporation de leucine marquée.
3.1.3 Stockage de la sécrétion
Une à deux heures après le début de l'expérience, la radioactivité est insignifiante dans le réticulum ou dans l'appareil de Golgi alors que les granules situés au pôle apical de la cellule sont fortement marqués. Les grains de zymogène , ou vésicules sécrétrices, ont pris naissance à partir des dictyosomes de l'appareil de Golgi. Ce dernier concentre les enzymes et fournit l'emballage membranaire nécessaire à leur stockage. Le renouvellement des membranes des dictyosomes est extrêmement rapide : une trentaine de minutes suffisent à leur régénération complète.
3.1.4 Extrusion des grains de sécrétion.
La mobilisation des enzymes contenues dans les divers grains de sécrétion est possible après deux heures, sous l'influence de divers sécrétagogues ( c'est à dire de substances qui stimulent la sécrétion ) . La libération du contenu des granules est réalisée par exocytose : la membrane des granules fusionne avec la membrane apicale de la cellule et leur contenu est déversé dans la lumière de l'acinus.
L'excédent de surface membranaire apicale, provoqué par la fusion des membranes des granules avec la membrane plasmique, est rapidement compensé par endocytose de fragments membranaires qui vont alors s'incorporer au réticulum ou à l'appareil de Golgi ( recyclage membranaire ).
3.2 Principaux régulateurs de la sécrétion des cellules acineuses du pancréas
3.2.1 Les modulateurs de la sécrétion
Parmi les diverses molécules susceptibles d'induire l'exocytose des granules de zymogène, on retiendra les trois qui sont les mieux caractérisées .
- acétylcholine
Médiateur libéré par les terminaisons nerveuses parasympathiques, l'acétylcholine simule la libération des enzymes du pancréas exocrine. Cette action est inhibée par l'atropine alors qu'elle est reproduite par les parasympathomimétiques comme la pilocarpine.
- Cholecystokinine-pancréozymine ( ou CCK-PZ ou encore CCK)
Cette hormone produite par certaines cellules de la muqueuse de l'intestin grêle ( mais aussi par des cellules situées en diverses autres régions de l'organisme ) doit son nom à divers travaux anciens . Les auteurs qui avaient isolé l'hormone active sur la vidange des voies biliaires , l'avaient nommée cholécystokinine . Quinze années plus tard , une hormone active sur la sécrétion d'enzymes par le pancréas était nommée pancréozymine. Ce n'est encore que 20 ans plus tard qu'il a été montré qu'une seule et même molécule avait les deux actions, la CCK-PZ. La CCK-PZ est un polypeptide de 33 acides aminés . On lui connaît plusieurs formes dérivées, soit de poids moléculaire supérieur, soit ne comprenant qu'une partie de la séquence.
- Bombésine
La bombésine est un tétradécapeptide ( polypeptide de 14 acides aminés) isolé d'abord de la peau des Amphibiens . Elle a été retrouvée depuis dans des cellules de muqueuse gastrique ou duodénale et dans le cerveau de diverses espèces de Mammifères.
Chez le chien et chez l'Homme , les dosages radio-immunologiques de bombézine mettent en évidence une augmentation de la concentration plasmatique après le repas. Cela suggère une action modulatrice de ce peptide sur la libération des enzymes pancréatriques accompagnant la prise d'aliments.
REMARQUE : La gastrine et le peptide intestinal vasoactif ( VIP) deux autres hormones gastro-intestinales ont probablement aussi un rôle physiologique dans le contrôle de la sécrétion d'enzymes par les acini, alors que la sécrétine est inactive sur les cellules acineuses; elle agit par contre sur les cellules des canaux pancréatiques et stimule fortement la sécrétion de liquide et d'électrolytes du suc pancréatique .
3.2.2 Le couplage stimulus-sécrétion.
Lorsque l'un ou l'autre des trois modulateurs mentionnés, acétylcholine, CCK-PZ ou bombésine est ajouté au milieu qui irrigue le pancréas, ou appliqué, par micro-ionophorèse, à la périphérie cellulaire, on enregistre immédiatemment une modification du potentiel de membrane des cellules acineuses. La variation du potentiel membranaire est toujours faible ( quelques mV) mais divers arguments expérimentaux montrent bien que la dépolarisation n'est pas liée aux perturbations de perméabilité membranaire qui pourraient être postérieurs aux remaniements de la membrane apicale accompagnant l'extrusion des granules.
On note une très bonne corrélation entre la concentration de l'agent stimulateur, la dépolarisation de la membrane, l'augmentation de la conductance membranaire et la sécrétion des enzymes.
La séquence des mécanismes qui interviennent n'est pas totalement connue . On a pu noter cependant :
- très rapidement après la stimulation, une augmentation de la conductance des membranes basolatérales à plusieurs ions : Na+, K+,Cl-;
- immédiatemment après, une diminution de la conductance ionique au niveau des jonctions lacunaires , avec un découplage immédiat des cellules voisines.
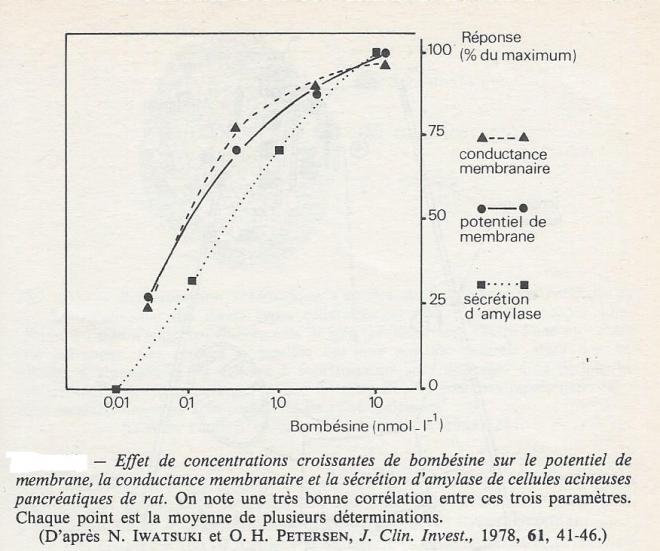
fig 3
L'augmentation de la concentration de Ca2+ dans le cytosol a très probablement le premier rôle dans le processus de couplage entre la stimulation et la réponse cellulaire, car les effets sur les perméabilités ioniques sont exactement reproduits , en absence de stimulation par les modulateurs, si l'on introduit du Ca2+ dans la cellule par ionophorèse.
Il est également probable qu'un nucléotide cyclique ( le GMPc ) soit impliqué dans les mécanismes qui mènent à l'extrusion membranaire.
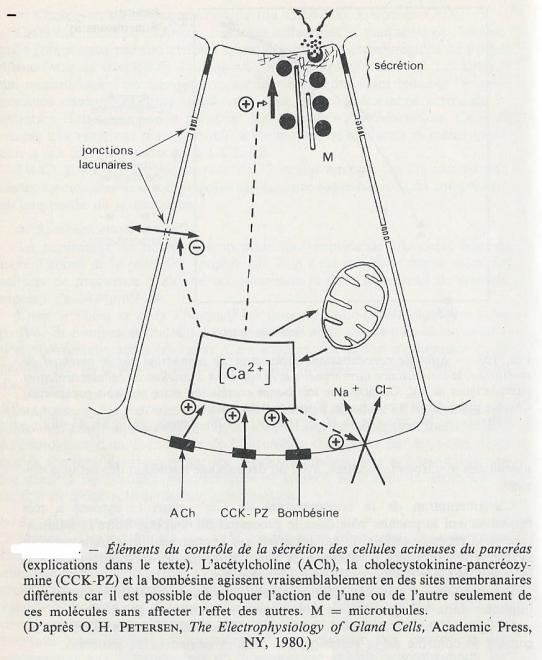
fig 4
4. La cellule B du pancréas endocrine
4.1 Les ilôts de Langerhans
Les îlots de Langerhans sont des enclaves de cellules de petite taille qui se trouvent disséminées au sein du pancréas exocrine chez les Vertébrés les plus évolués. Les Poissons cartilagineux possèdent des îlots de Langerhans alors que dans les espèces plus primitives les cellules endocrines sont encore disséminées dans l'épithélium digestif. Les cellules ne sont pas organisées autour de canaux collecteurs, mais sont richement irriguées et les sécrétions des diverses cellules sont d'abord déversées dans les espaces extracellulaires avant d'être drainées par le sang. La quantité de tissus représentée par les ilôts de Langerhans ne dépasse guère 1 à 1,5 % de la masse totale du pancréas.
L'histologie classique permet de révéler trois types cellulaires principaux en fonction de leurs affinités tinctoriales.
- Les cellules A, représentant environ 10 à 20 % des cellules sont surtout présentes à la périphérie des îlots. Elles sécrètent le glucagon, polypeptide de 29 acides aminés, dont l'effet global est hyperglycémiant.
- Les cellules B forment la catégorie la plus présente , avec 60 à 80 % des cellules des îlots chez l'homme ou chez le Rat. Leur position est surtout centrale dans les îlots , ce sont les cellules à insuline.
- Les cellules D sont moins nombreuses et plus petites . Ces cellules se colorent peu avec les colorants basiques . Elles élaborent la somatostasine , polypeptide de 14 acides aminés , isolé initialement à partir d'extraits d'hypothalamus, sur base de son activité inhibitrice sur la libération de l'hormone somatotrope par l'hypophyse antérieure. La somatostasine a été retrouvée dans divers tissus de l'organisme depuis lors.
Cest trois types cellulaires appartiennent à la série des cellules "APUD" Le système APUD (ou amine precursor uptake and decarboxylation) est un système endocrinien diffus de l'organisme dont les cellules sont capables de capter et de stocker les amines et leurs précurseurs dans le but de sécréter des hormones polypeptidiques.
Les îlots de Langerhans renferment également des cellules qui sécrétent le polypeptide pancréatique , différent de l'insuline ou du glucagon, et dont le rôle physiologique est encore mal connu.
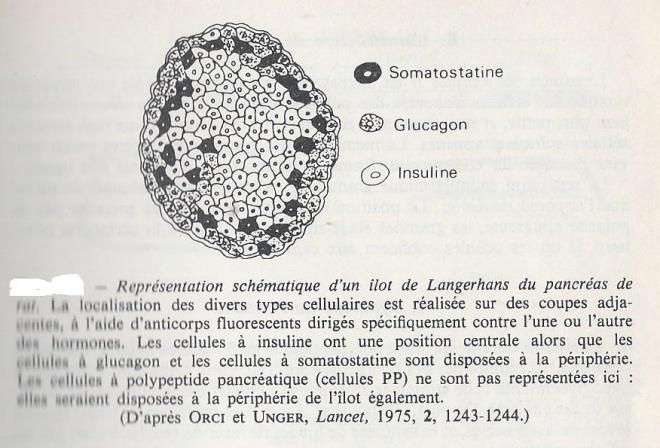
fig 5
Les diverses cellules de chaque îlots établissent des jonctions lacunaires nombreuses, au niveau desquelles peuvent transiter divers ions ainsi que des nucléotides cycliques , l'AMPc par exemple . Bien que chaque cellule ait conservé son unité structurale , on considère que l'ensemble des cellules d'un îlot fonctionne à la manière d'un syncytium, micro-organe très élaboré dans lequel chacun des types cellulaires présente une distribution numérique et topographique précise, vraisemblablement essentielle au fonctionnement intégré de l'ensemble.
4.2 Ultrastructure de la cellule B
L'examen de la cellule B en microscopie électronique révèle une structure typique des cellules élaborant des polypeptides. Les granules sécrétoires sont bien plus petits , et peu abondants par rapport à ceux qui s'observent dans les cellules acineuses voisines . La membrane des granules sécrétoires paraît souvent détachée du contenu, qui forme une sorte de cristal peu dense .
Le réticulum endoplasmique granulaire est assez peu développé , de même que l'appareil de Golgi. La position des différents organites ne présente pas de polarité apparente , les granules étant simplement situés à la périphérie cellulaire , là où les cellules confinent aux capillaires
4.3 Synthèse de l'insuline
Les phases de la synthèse et la libération de la molécule d'insuline peuvent être suivies par expérience comme l'autoradiographie après marquage des précurseurs de l'insuline. Après injection de Leucine tritiée à un animal, l'incorporation de la radioactivité est beaucoup moins intense dans les cellules des îlots de Langerhans que dans les cellules des acini voisins , mais le processus général de synthèse et de ségrégation des protéines marquées est tout à fait comparable. Il se forme d'abord un polypeptide de masse moléculaire élevée, la préproinsuline ( prépeptide de 11500 u.m.a), dont l'extrémité N-terminale est fortement hydrophobe et permet l'orientation de la synthèse vers l'intérieur du réticulum endoplasmique. La proinsuline, chaîne de 81 à 86 résidus d'acides aminés selon les espèces est rapidement détachée du prépeptide ; elle transite depuis le reticulum endoplasmique granulaire jusqu'à l'appareil de Golgi, puis la radio activité se concentre au niveau des granules de sécrétion. Le clivage de la proinsuline en insuline et peptide C au niveau de l'appareil de Golgi et les deux peptides sont intégrés dans les granules de sécrétion puisqu'ils sont ensuite sécrété de façon équimoléculaire . On pense que la synthèse du précurseur permet un assemblage des deux chaînes de la molécule d'insuline d'une grande fiabilité , alors que l'appariemment correct des ponts disulfures se fait avec un rendement très faible lorsque les chaînes A et B de la molécule d'insuline sont simplement mis en présence.
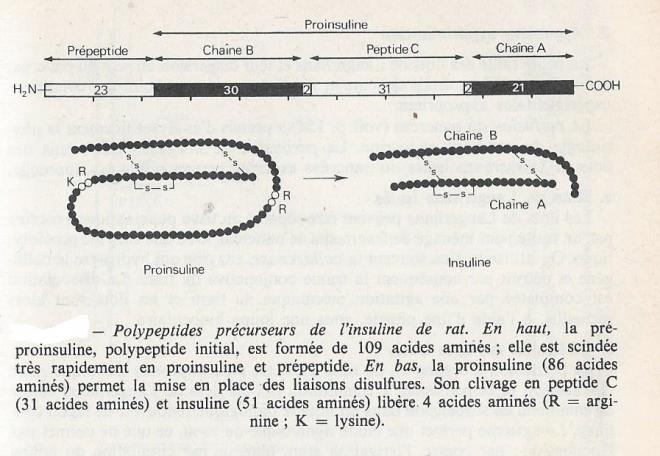
fig 6
4.3 Libération de l'insuline
La libération du contenu des granules de la cellule B du pancréas endocrine est réalisée par exocytose. La séquence des évènements qui assurent la continuité entre le stimulus physiologique et la libération d'insuline est assez bien connue.
4.3.1 Les stimulants de la cellule B
De très nombreux facteurs interviennent dans la régulation de la sécrétion d'insuline au sein de l'organisme.
Certains facteurs ont une action rapide , qu'il s'agisse de facteurs :
- nutritionnels : glucose, fructose
- hormonaux : cathécholamines ( adrénaline, noradrénaline) ou hormones gastro-intestinales (sécrétine, CCP-PZ ouy le peptide inhibiteur gastrique "GIP").
- neuro-hormonaux : acétylcholine...
D'autres facteurs ont une action beaucoup plus lente sur la sécrétion d'insuline : il peut s'agir de l'environnement hormonal qui s'installe durant la gestation ou des modifications complexes rencontrées lors du jeune.
Le glucose est le facteur insulinosécréteur le plus important sur le plan physiologique . C'est le stimulus qui a fait l'objet des études les plus complètes et c'est le seul qui sera envisagé ici .
Physiologie de la cellule Bêta
4.3.2 Reconnaissance du stimulus glucose par la cellule B
Deux théories ont été longtemps proposées pour expliquer le mécanisme de reconnaissance du glucose par les cellules B :
- Présence d'un récepteur stéréospécifique du glucose, situé probablement au niveau de la membrane plasmique
- signal intracellulaire apparaissant lors du métabolisme du glucose , à l'intérieur de la cellule B. Les arguments en faveur de la deuxième théorie sont plus nombreux ; pour n'en citer que deux :
- L'aptitude de divers glucides ( plus de vingt ont été essayés ) à faire sécréter de l'insuline est directement proportionnelle à la façon dont ils sont métabolisés dans la cellule B,
- Il est possible de faire sécréter de l'insuline en l'absence de sucre métabolisable dans le milieu extracellulaire , en provoquant simplement l'utilisation des réserves de glycogène de la cellule b ; le glycogène fournit alors du glucose qui est métabolisé sur place.
C'est donc bien la variation de la concentration intracellulaire de glucose ou de certains de ses métabolites qui déclenche le processus menant à l'exocytose des granules d'insuline.
L'hypothèse qui prévaut à l'heure actuelle ( "fuel hypothesis" des auteurs anglo-saxons ) est de considérer comme facteurs essentiels du couplage stimulus-sécrétion, ,les ions H+ et les coenzymes pyridiniques réduits ( NADH et NADPH ) qui sont formé lors du métabolisme du glucose ou des substrats tels que le pyruvate ou le lactate . Dans le même temps , le métabolisme produit l'énergie nécessaire à l'exocytose, sous la forme de molécules d'ATP .
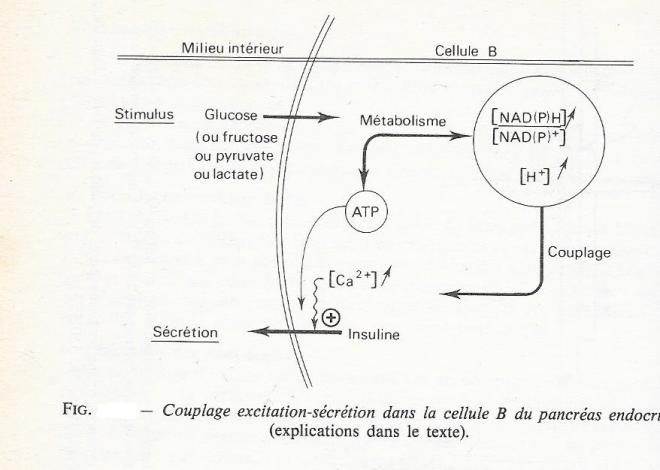
fig 7
4.3.3 Activité électrique de la cellule B et mouvements ioniques
a. Activité électrique de la cellule B
C'est par commodité que l'on parle d'activité électrique de la cellule : ce sont en réalité les mouvements ioniques qui s'accompagnent entre autres, de variation de l'équilibre électrique global de la cellule. L'activité électrique n'est pas une finalité dans le cas présent.
Les cellules B des îlots de Langerhans sont le type des cellules glandulaires excitables par un stimulus électrique, comme toutes les cellules chromaffines des glandes medullo-surrénales, diverses cellules adénohypophysaires ou encore certaines cellules glandulaires rencontrées chez des invertébrés.
Lorsqu'une dépolarisation membranaire est appliquée à ces cellules , elles réagissent ( de la même façon que lorsque leurs stimulants chimiques physiologiques les atteignent ) par une perturbation importante de leur activité électrique et on peut observer un potentielm de pointe , comparable par son aspect à ceux que l'on connaît pour les cellules nerveuses. La stimulation électrique d'une cellule B du pancréas endocrine est suivie de l'extrusion des granules sécrétoires.
Lorsque le potentiel de membrane d'une cellule B est enregistré en faisant varier la concentration extra-cellulaire de glucose, il reste stable tant que la concentration du glucose reste en deçà du seuil insulinosécréteur . Au dessus de ce seuil , la cellule présente une activité électrique , en même temps qu'elle commence à sécréter l'insuline . Ces observations attestent la signification physiologique de l'activité électrique de la cellule B.
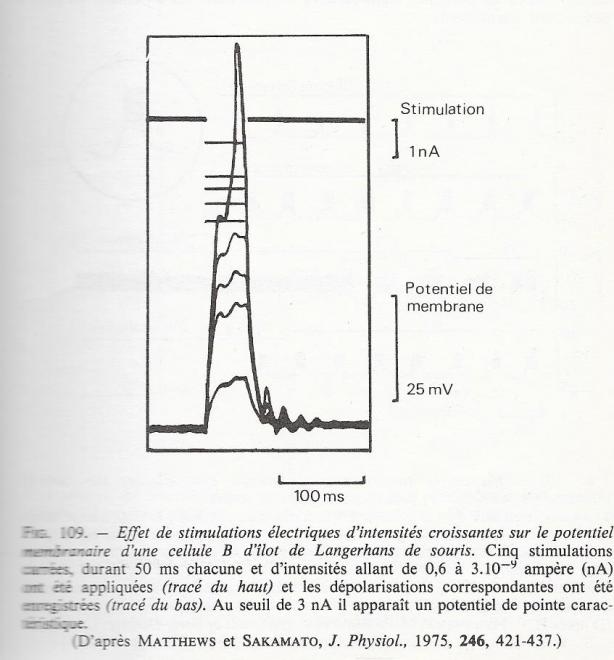
fig 8
La figure qui suit montre les variations du potentiel de membrane de la cellule B d'îlot de Langerhans, placé en périfusion, quand on modifie la concentrtation de glucose dans le milieu. Pour les concentrations de glucose efficaces sur la sécrétion d'insuline ( au dessus de 4 ou 5 mmol/l soit 0,7 à 0,9 gr/l ) il se produit une lente dépolarisation de la membrane , depuis la valeur de polarisation maximale ( vm) jusqu'à un seuil ded potentiel ( vs) à partir duquel la dépolarisation s'installe brutalement à une valeur plateau ( vp) pour une durée variable , avec une bouffée de potentiels de pointe surimposés. . Les potentiels de pointe ont une amplitude , une durée et une forme régulières : il s'agit de potentiels calciques.
L'évolution des variations lentes de la polarisation membranaire ( entre vm et vp ) est commandée précisément par la concentration en glucose :
- quand la concentration en glucose augmente, la périodicité des variations lentes augmente et les périodes avc potentiels de pointe sont prolongées :
- aux concentrations de glucose très élevées ( extraphysiologiques ) les oscillations lentes du potentiel membranaire cessent mais les potentiels de pointe deviennent permanents ;
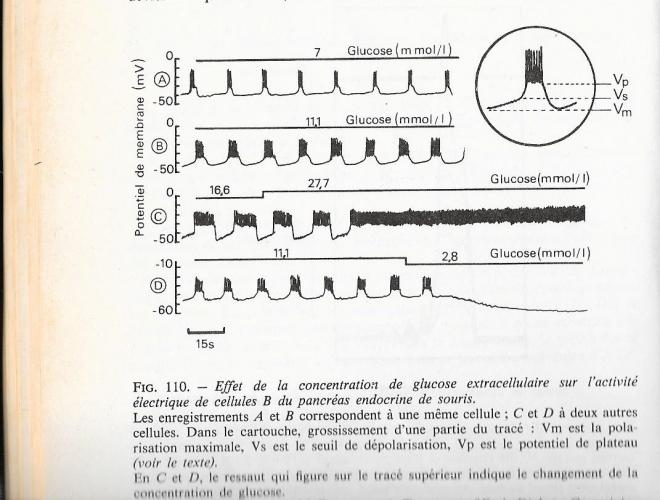
fig 9
- Lorsque la concentrtation de glucose est très faible ( 2,8 mmol/l )la polarisation membranaire est maximale et ne varie plus .
b. Les mouvements ioniques
Les mouvements ioniques qui gouvernent les variations du potentiel de membrane seraient les suivants :
- Le passage de Vm à Vs est lié à une diminution de la conductance au potassium ( gK+), ,provoquée par la production accrue des ions H+ et/ou par l'augmentation du rapport NADPH/NADP+ qui succèdent à l'augmentation du métabolisme du glucose. Le K+ s'accumulerant dans la cellule , la dépolarisation membranaire s'installerait par une accumulation de charges positives
- Lorsque la dépolarisation atteint le seuil Vs, l'ouverture d'un canal à calcium voltage dépendant se produit, et les ions Ca2+ s'engouffrent dans la cellule par suite du gradient de concentration qui existait entre le milieu extracellulaire et l'intérieur de la cellule . Le flux de Ca2+ vers le cytosol serait encore renforcé par une mobilisation de Ca2+ séquestré dans les mitochondries et les caccules du réticulum ( en effet, le phosphoénolpyruvate, l'un des métabolites produits par la glycolyse, provoque un efflux de calcium, des mitochondries vers le cytosol . Les mouvements de Ca2+ permettent ainsi de comprendre les variations du potentiel membranaire entre les valeurs Vs et Vp.
Les mouvements de l'ion sodium sont également modifiés par la concentration de glucose extracellulaire , mais la variation nette reste peu importante et ne peut rendre compte des différences de potentiel membranaire enregistrées.
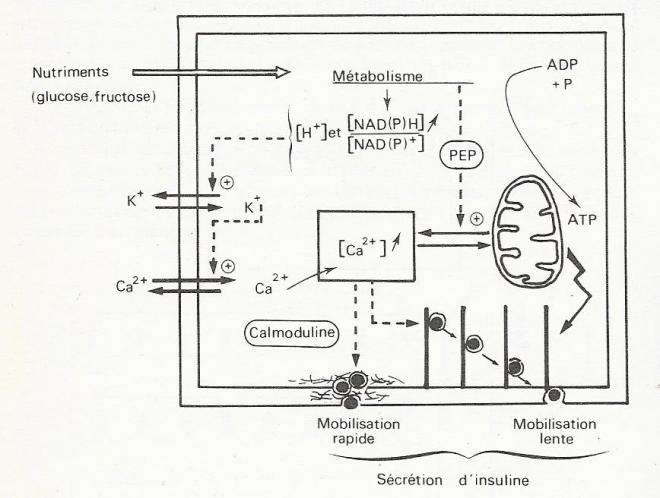
fig 10
4.3.4 Translocation des granules et exocytose
Divers auteurs ont évoqué les similitudes qui existent entre le mécanisme d'exocytose des granules d'insuline lors de la stimulation d'une cellule B et le mécanisme de la contraction des fibres musculaires striées. En particulier, on note dans les deux cas :
- le besoin d'énergie sous forme d'ATP,
- une dépolarisation membranaire précédant le mouvement
- la nécessité des ions Ca2+
L'observation directe des celkules ben culture a montré que le mouvement des granules vers la périphérie cellulaire se fait de façon " saltatoire " , chaque granule présentant des déplacements saccadés, dans une seule direction , et à des vitesses de 1,5 micromètres /seconde. Lorsqu'un granule se déplace , ses voisins peuvent rester totalement immobiles . Ces observations font penser à une intervention du cytosquelette dans la mobilité des granules. Des arguments expérimentaux, d'ordre pharmacologique , permettent d'imaginer un schéma cohérent ( fig 10 ) qui fait intervenir les microfilaments et les microtubules présents dans les cellules B .
Les granules d'insuline situés à proximité de la membrane plasmique sont retenus par le feutrage de microfilaments qui tapissent la face cytoplasmique de la membrane. Lors d'une augmentation de la concentration de glucose , les mouvements ioniques qui s'ensuivent entraînent l'augmentation du Ca2+ cytoplasmique . L'intervention d'un intermédiaire , qui serait la calmoduline , provoque un mouvement des microfilaments et l'extrusion des granules . La cytochalasins B, qui dépolymérise les microfilaments , provoque en effet une libération immédiate d'insuline , comme si les granules avaient été simplemet retenus jusqu'alors.
Si la stimulation par le glucose est maintenue , le recrutement des granules situés plus loin de la surface cellulaire ( granules de réserve ) fait appel à un mécanisme différent, avec intervention des microtubules. Les substances qui interfèrent avec la polymérisation de la tubuline ( colchicine, vincristine ) inhibent en effet la deuxième phase de la sécrétion d'insuline alors qu'elle sont sans effret sur la première phase.
VII. D'AUTRES ASPECTS DE LA PHYSIOLOGIE DES CELLULES SECRETRICES
A.Les types sécrétoires
1.Modalités de la sécrétion
a) Cellules endocrines et exocrines
- Les cellules exocrines exportent leur produit de sécrétion soit vers une cavité naturelle de l'organisme (tube digestif, voies respiratoires ou génito-urinaires ), soit vers l'extérieur de l'organisme . La cellule acineuse du pancréas en est le type, ainsi que les cellules des glandes salivaires , celles des glandes sudoripares ou de la glande mammaire...
- Les cellules endocrines déversent leurs sécrétions ( hormones ) dans les espaces intercellulaires. De là, la sécrétion gagne la circulation sanguine . Les hormones agiront à une distance plus ou moins importante sur certaines cellules cibles.
- Les cellules neuroendocrines ( ou neurosécrétrices )sont des neurones dont les fonctions de synthèse sont hypertrophiées. On les rencontre dans le système nerveux : ce sont par exemple les cellules à hormones anti-diurétique ( ADH ) situées dans l'hypothalamus des mammifères, ou les cellules neurosécrétrices de la pars intercerebralis du cerveau des insectes (fig 11)
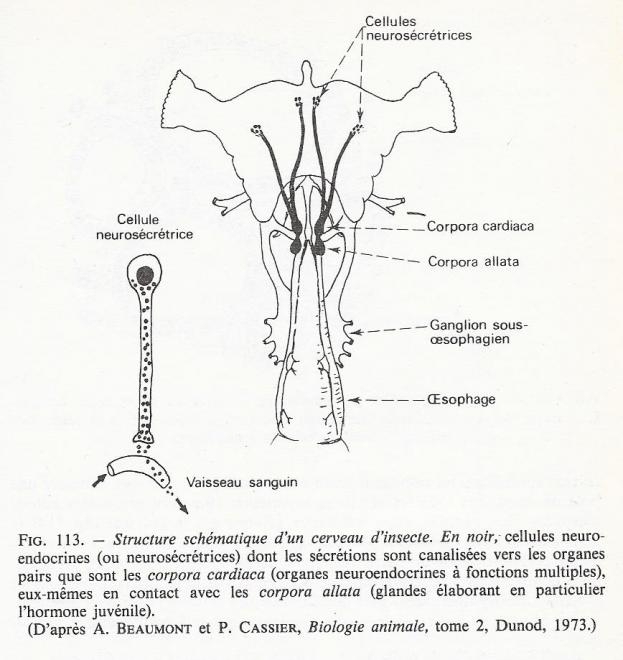 b) Cellules non glandulaires
b) Cellules non glandulaires
Lorsque des cellules sécrétent des composés qui s'accumulent autour d'elles, on ne peut les rattacher aux cellules glandulaires, bien que leur physiologie soit très proches, c'est le cas des chondrocytes, cellules du cartilage , qui élaborentdes protéines complexes associées à divers sucres, comme le xylose et le galactose, et à des radicaux sulfates ( protéoglycanes). Les ostéoblastes , cellules qui participent à l'élaboration du squelette ont également une forte activité synthétique : elles élaborent et déposent autour d'elles le collagène et les glycoprotéines qui forment la matrice organique des os.
c) sécrétion continue ou cyclique
Lorsque la sécrétion est continue, le produit de sécrétion est libéré au fur et à mesure de sa formation; c'est le cas pour les cellules à mucus de l'épithélium gastrique ou pour les cellules folliculaires de la thyroïde par exemple . Les diverses phases du processus sécrétoire se déroulent simultanément dans la cellules.
Dans d'autres cas , le cycle sécrétoire est discontinu : synthèse et transport intracellulaire ( ségrégation ) sont suivis de l'accumulation du produit élaboré dans des granules de stockage. les exemples vus précédemment appartiennent à cette catégorie .
2.Nature de la sécrétion
Les sécrétions fluides sont encore appelées sécrétions séreuses ( c'est le cas le plus fréquent ). Les sécrétions visqueuses sont aussi appelées sécrétions muqueuses.
Les substances sécrétées peuvent être très diverses :
- électrolytiques ( bicarbonates, acide chlorhydrique ...),
- protéiques ou polypeptidiques ( enzymes, hormones ;;;),
- glycoprotéiques ( cellules à mucus ),
- dérivées d'acide aminés ( hormones thyroïdiennes ...),
- stéroïdiennes, etc.
B. Cellules sécrétant des dérivés d'acides aminés
1. Les cellules folliculaires de la thyroïde
a. Thyroïde et hormones thyroïdiennes
La glande thyroïde dérive de la région céphalique du tube digestif. De leur origine épithéliale, les cellules thyroïdiennes ( ou thyréocites ) ont conservé une polarité marquée / Chez les vertébrés supérieurs elles sont organisées autour d'espaces clos , remplis d'une substance gélatineuse, la colloïde ( fig )et délimitent ainsi des follicules pouvant atteindre un mm de diamètre. La vascularisation de la thyroïde est particulièrement développée . L'innervation est également dense et l'on note des terminaisons catécholaminergiques ( appartenant au système orthosympathique ) près de la basale des cellules; ceci peut être considéré comme un indice de leur rôle régulateur sur le fonctionnement thyroïdien;
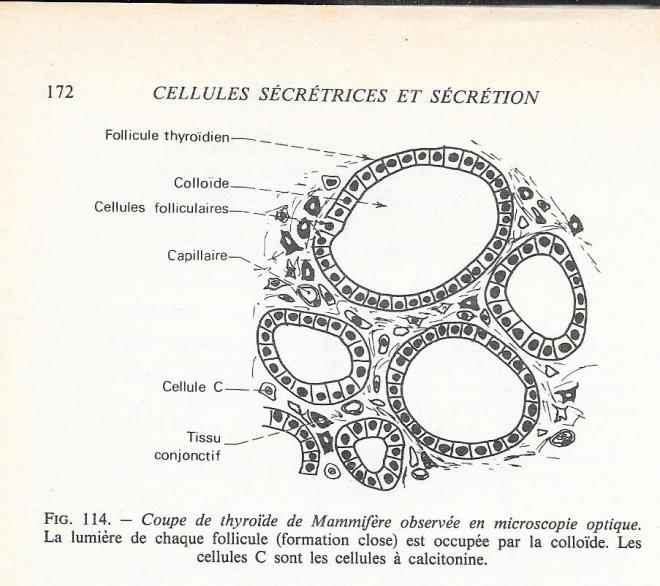
Entre les cellules se trouvent des cellules parafolliculaires beaucoup moins nombreuses, les cellules C ( "C" pour claires ). Les cellules C appartiennent à la série "APUD". Elle sécrétent la calcitonine , polypeptide hormonal hypocalciémant.
Les cellules folliculaires de la thyroïde sécrétent des dérivés iodés de l'acide aminé tyrosine que l'on regroupe sous le terme d'hormones thyroïdiennes. La thyroïde est une glande endocrine particulière à plus d'un titre , mais spécialement par ses possibilités de stockage des hormones ou de leurs précurseurs au niveau de la colloïde. Les réserves peuvent ainsi être équivalentes à plusieurs mois de sécrétion chez l'homme, alors que les autres glandes endocrines ont une réserve hormonale qui ne dépasse pas en général les besoins quotidiens..
Dans le sang la T4, est la plus abondante ( environ 90.pc ), mais l'hormone physiologiquement active est la T3. Une partie de la T3 provient de la désiodation de la T4 par divers tissus . La T3 réserve est normalement très peu abondante ; elle est augmentée lors de certains états pathologiques ( jeunes, maladies graves ) et chez le nouveau né. Les actions physiologiques de l'hormone thyroïdienne sont multiples ( morphogenèse, stimulation des oxydations celluleires , avec accélération du métabolisme )
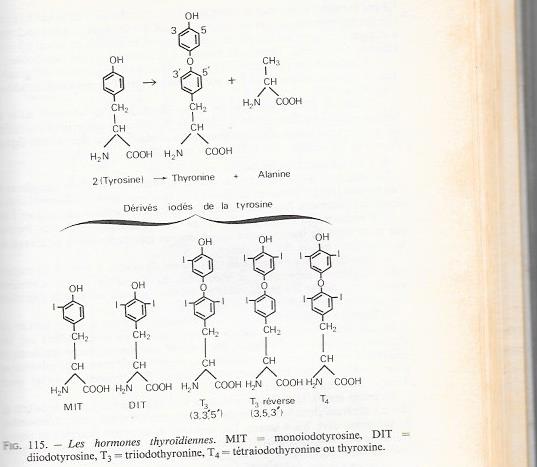
b) Synthèse des hormones thyroïdiennes
Les étapes de la synthèse des hormones thyroïdiennes sont très complexes et l'on ne peut présenter qu'un schéma qui ne peut tenir compte de la simultanéité des divers processus , les étapes essentielle sont schématisées à la figure qui suit :
- Captation des acides aminés et des sucres ( galactose, mannose, fucose ) par un système de transporteurs membranaires spécifiques situés au niveau de la membrane basale des cellules.
- Absorption des iodures par un système de pompes à iodures ( transport actif ) également situé dans la membrane basale de la cellule . Certaines substances comme les perchloractes ou les thyocyanate inhibent la captation des iodures entrant en compétitions avec eux .
- Synthèse de la thyroglobuline La synthèse protéique a lieu au niveau du réticulum granulaire . Les chaînes polysaccharidiques sont greffées sur les chînes protéiques ( la thyroglobuline est une protéine de masse moléculaire 680000 , formée de quatre sous unités ) au niveau du réticulum endoplasmique pour certains sucres ( mannose , glucosamine ) de l'appareil de Golgy pour d'autres ( fucose, galactose ).
En même temps la molécule a transité vers la partie apicale de la cellule et elle est déversée dans la lumière du follicule sans avoir été iodé . L'ensemble du processus dure de 2 à 4 heures . Les vésicules de sécrétions chargées de thyroglobuline sont mues par le système de microtubules et microfilaments du cytosquelette .
- Oxydation des iodures . La réaction d'oxydation des iodures ,
I- + H2O2 ---> I0 + H2O + O,
est catalysée par une péroxydase localisée dans les microvillosités apicales des cellules. L'iode est alors exporté vers la colloïde . Les substances comme le propylthiouracile ( PTU) ou le carbimazole qui empêchent l'organification de l'iode, c'est à dire sa fixation sur les molécules organiques telles que les hormones thyroïdiennes , agissant à la fois en inactivant la péroxydase, mais aussi en empêchant la condensation des iodotyrosines en T3 ou T4
- iodation des radicaux tyrosils de la thyroglobuline . Elle se fait à l'intérieur du follicule en quelques minutes seulement .
- Elaboration des hormones iodées et sécrétion.
La configuration spatiale de la thyroglobuline permet le rapprochement des radicaux tyrosils iodés, qui précède leur condensation en une molécule de thyronine . L'élaboration des hormones se fait à l'intérieur de la cellule qui pour cela absorbe par endocytose , la thyroglobuline néosynthétisée
Les vésicules d'endocytose se distinguent des vésicules d'exocytose dans une même cellule par leur taille supérieure . Les vésicules d'endocytose migrent cette fois vers le pôle basal de la cellule.
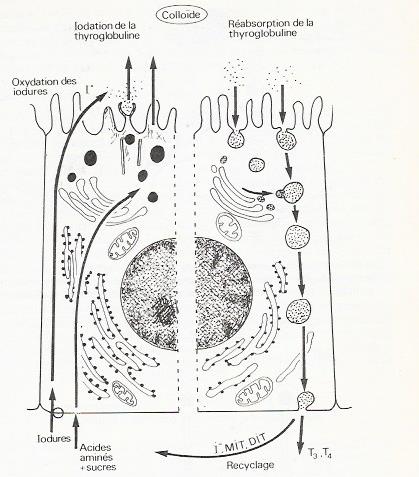
La formation des hormones T3 et T4, fait appel au processus d'hydrolyse lysosomiale . Des lysosomes primaires détachés de l'appareil de Golgy vont fusionner avec les vacuoles contenant la thyroglobuline iodée pour former des lysosomes secondaires . Les enzymes lysosomiales ( glucosidases, peptidases ) hydrolysent alors la thyroglobuline . Les produits de l'hydrolyse , acides aminés, sucres , T3, T4, MIT, DIT, se retrouvent dans le cytoplasme. De la il peuvent gagner les espaces extracellulaires et la circulation sanguine , c'est le cas pour la T3 et la T4, ou bien être recyclés dans le même cellule , ce qui est le cas pour l'iode, les sucres ou les acides aminés. La sécrétion des hormones thyroïdienne est régulée par un système d'interaction entre la thyroïde, l'hypophyse et l'hypothalamus.
2. Les cellules de la médullo-surrénale
La médullo-surrénale est située dans la partie profonde de la glande surrénale . Il s'agit d'une glande endocrine dont les cellules contiennent de fines granulations qui brunissent lorsqu'elles sont oxydées par le dichromate de potassium ; il s'agit de la réaction chromaffine , et les cellules qui réagissent positivement à cette réaction sont appelées cellules chromaffines . Les cellules chromaffines de la médullo-surrénale élaborent et sécrètent des catécholamines : adrénaline essentiellement , mais aussi un peu de noradrénaline . Les catécholamines peuvent être localisées en microscopie électronique , par leur propriété de réduire le tétroxyde d'osmium , avec formation d'un produit dense aux électrons . Leur synthèse débute près de la membrane nucléaire , où apparaissent les goutelletes issues de l'appareil de Golgi. Ces goutellettes migrent vers la membrane plasmique en même temps que leur taille et leur densité aux électrons augmentent . Elles forment alors des granules sécrétoires de 100 à 300 nm de diamètre.
La décharge des catécholamines dans l'espace extracellulaire est provoquée par une excitation de la cellule via les neurones symphatiques préganglionaires qui libèrent de la noradrénaline au contact de la cellule chromaffine L'exocytose est ici encore calcium dépendante et ferait intervenir les éléments du cytosquelette.
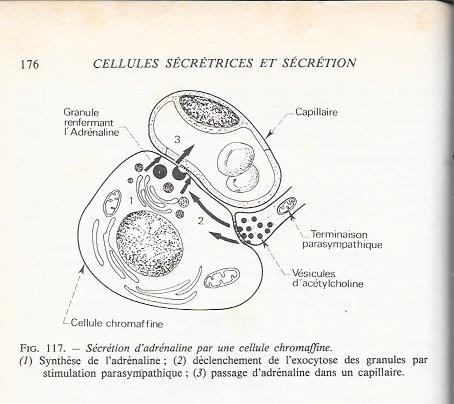
C. Cellules sécrétant un mucus glycoprotéique
Les cellules exocrines de ce type sont rencontrées en divers points de l'organisme : voies respiratoires, tube digestif, voies génitales . Le type en est la cellule caliciforme de ma muqueuse intestinale , glande cellulaire mucipare , située de place en place entre les cellules absorbantes .
La cellule se caractérise par la présence de très gros granules peu colorables qui encombrent son cytoplasme et le font ressembler à ces sacs de boules de coton à démaquiller que l'on trouve dans le commerce ; les granules sont par contre denses aux électrons . Ils sont formés de glycoprotéines . Le noyau se trouve repoussé en position basale , dans une zone où le réticulum granuleux est très développé .
Les études dynamiques de la synthèse protéique ont permis de mettre en évidence un mécanisme tout à fait comparable à celui qui existe dans les cellules acineuses du pancréas, à ceci près que dans le cas de la cellule mucipare , des motifs glucidiques sont associés aux chaînes protéiques au niveau de l'appareil de Golgi.
Date de dernière mise à jour : 30/03/2018
Commentaires
-
ces documents sont assez vieux, veuillez attendre un peu que je retrouve les sources !
Ajouter un commentaire