

Physiologie animale 3
Transmission synaptique
I.3 TRANSMISSION SYNAPTIQUE DE L'INFLUX NERVEUX
Au niveau de la synapse, la transmiussion de l'influx nerveux d'un neurone sera réalisée par voie chimique.
Un médiateur appelé " neurotransmetteur, libéré par la zone présynaptique ira se lier chimiquement avec des récepteurs se trouvant sur la membrane postsynaptique.
Le résultat sera une dépolarisation plus ou moins prolongée , locale dite potentiel postsynaptique d'excitation , qui engendre l'influx postsynaptique.
![]()
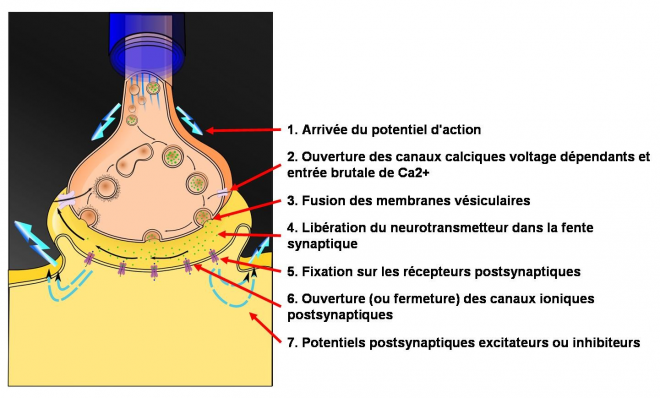
La libération des neurotransmetteurs est induite par l'arrivée du PA au niveau de la terminaison présynaptique: la dépolarisation des membranes des terminaisons nerveuses ouvre des canaux calciques voltage-dépendants situés sur la membrane présynaptique, au niveau des zones actives. L'entrée de Ca2+ dans les terminaisons pré-synaptiques est liée à la forte différence de concentration entre les milieux extracellulaire (à plus forte concentration de Ca2+) et intracellulaire (à très faible concentration de Ca2+), source d'une importante force électromotrice .
L'augmentation brutale de la concentration intracellulaire présynaptique de Ca2+ entraîne la fusion des membranes des vésicules présynaptiques avec la membrane plasmique de la terminaison axonique, au niveau des zones actives. Les vésicules présynaptiques déversent leur contenu dans la fente synaptique. Ce processus de libération des neurotransmetteurs est nommé exocytose. Il s'agit d'un processus extrêmement rapide, survenant dans les 0.2 ms suivant l'influx d'ions Ca2+ dans la terminaison. Il est probable que les vésicules concernées sont celles qui sont déjà "arrimées" aux zones actives de la synapse, via des protéines de membrane des synapses et des zones actives . En présence d'une forte concentration de Ca2+, ces protéines changent de conformation, entrainant la fusion membranaire et la libération du contenu vésiculaire. Les processus de fusion des membranes dépendent des protéines membranaires de la superfamille SNARE situées d'une part sur la membrane vésiculaire et d'autre part sur la membrane présynaptique. Ces protéines sont complémentaires les unes des autres ce qui permet aux vésicules de s'associer aux membranes présynaptiques (cette association est dénommée le docking). Leur changement de conformation sous l'effet du Ca2+ entraine la fusion des membranes.
Les granules de sécrétion déversent les neuropeptides dans la fente synaptique également par exocytose déclenchée par l'augmentation de la concentration intracellulaire de calcium mais ceci a lieu à distance des zones actives. Etant donné que ces sites d'exocytose sont plus éloignés des zones d'entrée du Ca2+, il est nécessaire que la concentration en Ca2+ s'élève suffisamment pour que les neuropeptides soient libérés. Il n'ya donc pas de libération de neuropeptides à chaque PA: celle-ci nécessite un train de PA à haute fréquence . La Libération de neuropeptides est par ailleurs un processus plus lent, qui demande 50 ms ou plus .
Après fusion avec la membrane présynaptique, les constituants de la membrane sont recyclés dans la terminaison présynaptique afin de reconstituer de nouvelles vésicules. Les vésicules sont en effet initialement synthétisées dans le corps cellulaire du neurone mais la distance entre le soma et la terminaison synaptique ne permet pas d'assurer un transport de vésicules suffisant en cas d'activité soutenue: le recyclage local des vésicules permet de fournir une quantité suffisante de neurotransmetteurs . La restitution de la vésicule dans le cytoplasme est un processus dénommé endocytose . Dans les périodes d'intense stimulation, d'autres vésicules prises dans une "réserve" liée au squelette de la terminaison axonale peuvent intervenir . L'élévation intracellulaire de Ca2+ mobilise ces vésicules et permet leur arrimage aux zones actives de la membrane .
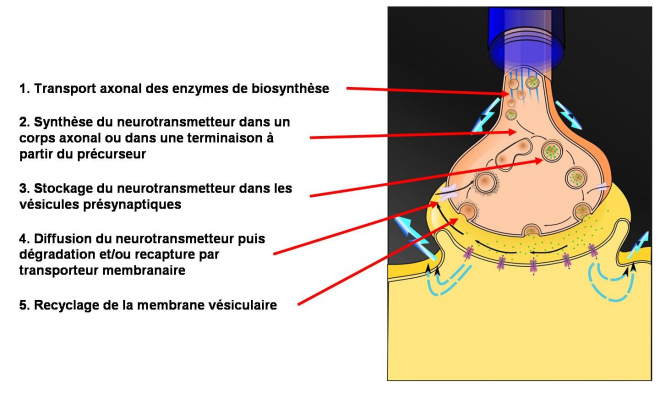
La liaison neurotransmetteur-récepteur doit ensuite être rapidement interrompue afin de permettre la transmission d'un nouveau signal chimique en rapport avec l'arrivée de nouveaux potentiels d'action. Le neurotransmetteur peut simplement diffuser hors de la fente synaptique, être dégradé dans la fente synaptique ou être recapturé soit par la cellule présynaptique soit par les cellules gliales environnantes.
La réintégration intracellulaire est un processus actif, s'effectuant à l'aide de transporteurs protéiques spécifiques des neurotransmetteurs, situés dans la membrane présynaptique . Les neurotransmetteurs sont ensuite détruits par des enzymes spécifiques du cytosol ou réincorporés dans les vésicules synaptiques. Les cellules gliales situées autour de la synapse jouent ici un rôle très important en éliminant les neurotransmetteurs de l'espace synaptique . C'est notamment le cas des acides aminés excitateurs comme le glutamate, éliminé de l'espace synaptique par les astrocytes périsynaptiques et/ou des transporteurs situés dans la partie postsynaptique de la synapse .
Une destruction enzymatique des neurotransmetteurs dans l'espace synaptique peut également permettre leur élimination. C'est le cas de l'inactivation de l'Ach au niveau de la jonction neuromusculaire . L'enzyme acétylcholinestérase (AChE) détruit la molécule d'ACh, la rendant inactive au niveau des récepteurs à l'ACh .
I.3.1 Les neurotransmetteurs
a) Les amines biogènes
- Les catécholamines ( dopamine, noradrénaline, adrénaline ) dérivent d'un acide aminé : la tyrosine.
- La sérotonine est synthétisée à partir du trypotophane
b) L'acétylcholine
Est synthétisée dans la zone présynaptique à partir de l'acétyl-CoA ( origine mitochondriale ) et de la choline que l'axone puise dans le milieu extra)cellulaire. La cholinestérase contrôle l'acétylation de la choline .

c) Le gaba, décarboxylation de l'acide glutamique.
I.3.2 Les vésiculers synaptiques
Les neurotransmetteurs sont stockés dans des véhicules.
- Les vésicules contenant des amines biogènes adrénergiques proviendaraient de la face de maturation de l'appareil de Golgi.
- Les vésicules contenant de l'acétylcholine ( cholinergique )se formaient localement par endocytose de l'axolemme présynaptiques.
I.3.3 Les récepteurs de la membrane postsynaptique
Il existe deux types de récepteurs sensibles aux médiateurs
a) les récepteurs à action rapide : le transfert de l'information est assuré en agissant sur la perméabilité d'un canal ionique.
b) les récepteurs à action durable : activés par le neurotransmetteur. Ils entraîneront la formation d'un second messager qui transmet l'effet du médiateur à l'intérieur du neurone. La plupart des médiateurs possèdent les deux types de récepteurs.
I.3.4 Le mécanisme de libération des neurotransmetteurs.
a) Migration des vésicules synaptiques : migration vers la membrane présynaptique.Mécanisme en jeu : complexe myosine - actine - calcium.
b) Fusion des vésicules avec la membrane présynaptique.
c) Libération du médiateur dans la fente synaptique, 2 mécanismes possibles :
- exocytose.
- passage par des synaptopores.
d) Liaison neurotransmetteur-récepteur :
- L'acétylcholine : ex : jonction musculaire. La fixation du médiateur sur la protéine réceptrice post-synaptique va provoquer une modification de la configuration spatiale de cette dernière, qui se matérialisera par une ouverture des canaux sodium : naissance du potentiel postsynaptique.
- L'adrénaline : ex. : innervation des cellules musculaires lisses par des fibres sympathiques amyéliniques. Le complexe médiateur-récepteur active l'adénylcyclase qui transformera l'ATP en AMP cyclique, facteur déclenchant l'effet biologique
- Le GABA : agit sur un récepteur dont le canalest sélectivement perméable aux ions chlorures. Lorsque ces ions pénètrent dans la cellule réceptrice par le canal ouvert, ils augmentent la différence de potentiel à travers la membrane et inhibent temporairement la cellule. Il s'agit donc, dans ce cas, d'un médiateur inhibiteur.
e) Le mécanisme d'inactivation du neurotransmetteur. L'action déclenchée par le médiateur est rapidement arrêtée soit :
- Par diffusion : en dessous d'un certain seuil de concentration , le médiateur est incapable d'engendrer un potentiel d'action.
- Par destruction enzymatique :
* les amines biogènes sont inactivées par la monoamine-oxydase localisée dans les mitochondries de l'axoplasme présynaptique et du cytoplasme postsynaptique ( MAO)
* l'acétylcholine est hydrolysée par les cholinestérases non spécifiques et par une enzyme spécifique : l'acétylcholinestérase.
- Par recapture : par la zone présynaptique . Le médiateur récupéré sera repris par des vésicules synaptiques et disponible de la sorte , pour un nouvel emploi. Les amines biogènes sont recaptées, de la sorte, en totalité. L'acétylcholine ayant été hydrolysée, seule la fraction choline sera récupérée par un mécanisme d'endocytose. L'acétate, quant à lui, pénètre dans la région post-synaptique.
Résumé :
- Le neurone présynaptique synthétise les molécules de neurotransmetteurs qui sont stockées dans les vésicules synaptiques au niveau des boutons terminaux.
- L’arrivée de l’influx nerveux (potentiel d’action) au niveau de la terminaison présynaptique provoque la dépolarisation de la terminaison axonale.
- La dépolarisation entraîne l’entrée d’ion calcium (Ca2+) dans la terminaison axonale, grâce à l’ouverture de canaux calcique voltage dépendant présent en grande concentration au niveau des boutons synaptiques.
- Le calcium permet le mécanisme de fusion de la membrane des vésicules synaptiques à la membrane présynaptique.
- Le neurotransmetteur est alors libéré par exocytose dans la fente synaptique. Cette exocytose nécessite de l’énergie fournie par les mitochondries de la terminaison présynaptique.
- Le neurotransmetteur se fixe à ses récepteurs spécifiques localisés sur la membrane post-synaptique.
- Cette fixation entraîne, selon la nature du neurotransmetteur, du récepteur et de la chaîne de transduction, l’ouverture ou la fermeture de canaux ioniques post-synaptique.
- Les variations de conductances ioniques modifient alors la polarisation électrique de cette membrane de façon plus ou moins importante. On parle de potentiel post-synaptique (PPS). La naissance de ce courant post-synaptique modifie le signal électrique du neurone post-synaptique.
- Le neurotransmetteur après action est ensuite inactivé ce qui limite l’existence du PPS. La transmission synaptique est ainsi rendue efficace grâce à l’inactivation rapide des neurotransmetteurs.
Date de dernière mise à jour : 30/03/2018
Ajouter un commentaire


