

Physiologie animale 2
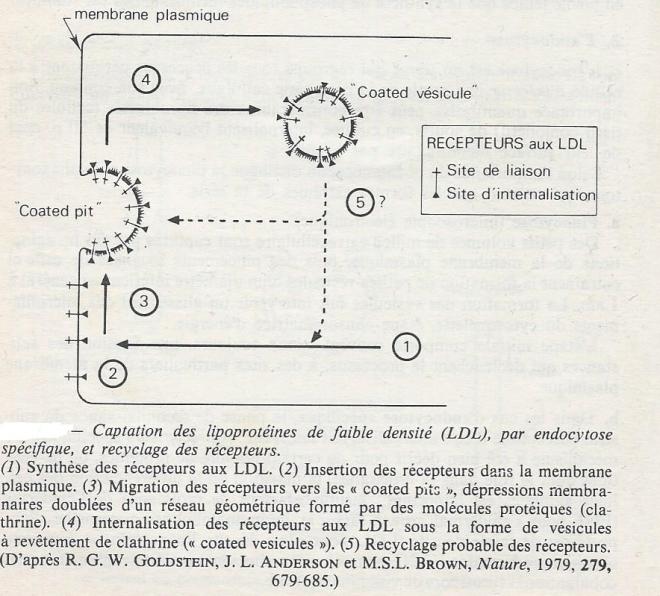
La captation des LDL ( particules d'environ 20nm de diamètre , renfermant des protéines, des phospholipides, du cholestérol) par des fibroblastes humains en culture a été particulièrement étudiée. On admet que son rôle essentiel serait l'incorporation du cholestérol , composant lipidique important des membranes plasmiques , mais dont seul quelques types cellulaires font la synthèse. Le mécanisme met en jeu des récepteurs spécifiques qui sont drainés vers des zones particulières de la membrane ( coated pits , ce qui pourrait se traduire par " dépression à revêtement" ) dès qu'ils ont fixés les LDL , puis internalisés dans des vésicules de quelques centaines de nm ( " coated vesicules "). La totalité du processus dure moins de 10mn. à 37 °C. Les protéines des LDL sont alors hydrolysées par des enzymes lysosomales et redonnent naissance a des acides aminés . Les récepteurs à LDL sont recyclés après leur internalisation . Synthétisés au niveau des polyribosomes fixés au réticulum, puis glycosylés au niveau de l'appareil de Golgy, ils sont insérés dans la membrane plasmique où ils gardent une grande mobilité , ce qui leur permet de se rassembler dans des " coated pits". Le recyclage peut être démeontré dans divers cas : les fibroblastes, par exemple , absorbent des taux de LDL en concentration saturante pendant plus de 6 heures, de façon constante , même quand la synthèse de tout protéine ( et par conséquent de tout nouveau récepteur ) est bloquée par le cycloheximide. En absence de recyclage, quelques minutes suffiraient pour épuiser le stock des récepteurs. Des travaux portant sur une quinzaine d'exemples de ce type de transport ont été effectués et les mécanismes sont très comparables.
 c). Phagocytose
c). Phagocytose
Comparable à la phagocytose , par son mécanisme, la phagocytose concerne l'ingestion de matériel particulaire beaucoup plus volumineux. Les amibes peuvent par exemple phagocyter des micro-organismes tels te les Rotifères ou les Ciliés.
La cellule phagocytaire ( amibe, granulocyte, macrophage...) émet des pseudopodes, prolongements cytoplasmiques qui vont encercler la particule cible avant de l'incorporer totalemen,t. Le mécanisme fait intervenir l'appareil motile de la cellule et il est calcium dépendant : on a isolé à partir de macrophages de poumon de Lapin , une protéine ( le " gelsoline"), qui se fixe à l'actine des microfilaments , à condition que la concentration de Ca2+soit supérieure à 10-7 moles /l dans le cytosol .
Après la phagocytose, les vacuoles ( phagosomes ) fusionnent avec les lysosomes primaires dont les enzymes hydrolytiques attaquent le contenu, pour le transporter en molécules susceptibles d'êtres transportées à travers la membrane des vacuoles ( appelés lysosomes secondaires après la fusion lysosomes primaires-phagosomes.
Les phénomènes d'endocytose interviennent à divers niveaux de l'activité des cellules : ils sont aussi bien impliqués dans les activités d'absorption à signification nutritionnelle ou défensive ( phagocytose) , que dans les stockages de diverses substances de réserve ( vitellogénèses des ovocytes d'Insectes ), dans les transports transcellulaires ( cellules endothéliales de la paroi des capillaires ) ou le recyclage de divers composants des cellules ( endocytose faisant suite à l'exocytose de divers produits de l'activité cellulaire).
1.1.7.3 L'exocytose
Divers exemples d'exocytose seront analysés plus loin en détail : extrusion des granules de zymogène par les cellules acineuses du pancréas, de l'insuline par les cellules B des îlots de Langerhans du pancréas endocrine , libération de l'acétylcholineau niveau des terminaisons cholinergiques.
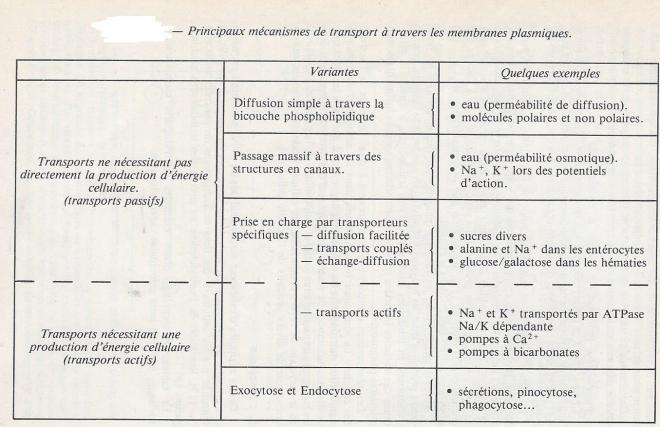
Le tableau suivant récapitule divers exemples de transports membranaires envisagés içi, avec leur principales caractéristiques.
I.2 NEURONES ET CONDUCTION RAPIDE DE L'INFORMATION
Le système nerveux constitue un appareil de communaication et d'intégration pour l'organisme, caractéristé par la rapidité de ses réactions et par la localisation précise des sites de son action. Son fonctionnement repose sur une infrastructure cellulaire très élaborée, aux connections multiples, ce qui lui confère à la fois une grande présision et une grande souplesse d'action.
I.2.1 Structure, environnement et physiologie du neurone
I.2.1.1 Le système nerveux
I.2.1.1.1 Définition
Le système nerveux d'un organisme est formé par l'ensembles des cellules nerveuses ( neurones ) et des cellules non nerveuses qui leur sont associées ( cellules gliales ). Les cellules nerveuses sont des cellules qui sont spécialisée dans l'émission , la réception, l'intégration et la conduction d'"états d'excitation ". Cette définition exclut les cellules effectrices mais s'applique aux cellules sensorielles. La fonction essentielle des neurones et donc de traiter et de créer de l'information à propagation rapide dans l'organisme.
Le système nerveux central renferme la plupart des corps cellulaires moteurs et des neurones intermédiaires du système nerveux.
Le système nerveux périphérique comprend tous les corps cellulaires des neurones sensitifs, des plexus nerveux locaux ( amas diffus de neurones et de fibres nerveuses), des ganglions nerveux ( amas nodulaires de neurones ) et les nerfs ( faisceaux d'axones ).
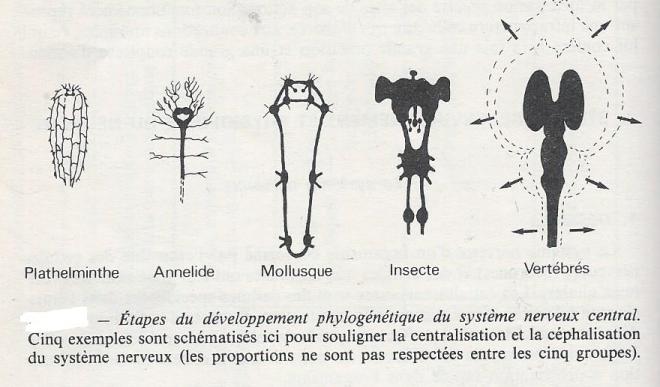
I.2.1.1.2 Phylogenèse
Le système nerveux n'existe chez les métazoaires qu'à partir du moment où les éléments fonctionnels, neurones, synapses, influx nerveux, peuvent être mis en évidence sans ambigüité. Ceci ne semble pas devoir être le cas chez les Spongiaires où l'on observe bien des cellules d'apparence nerveuse, mais ces cellules n'établissent pas ( ou peru) de connexions entre elles, et les réactions aux stimuli restent toujours très localisés chez ces animaux . Les Cnidaires possèdent un système nerveux très simple , avec parfois quelques ganglions et un réseau nerveux qui reste peu dense , il n'y a pas de système nerveux central dans ce groupe .
Le système nerveux central n'apparaît que chez les Plathelminth : un cerveau distinct est alors présent, avec des prolongement vers les autres parties du corps ; un plexus superficiel et plusieurs types d'organes sensoriels peuvent être mis en évidence.
Le système nerveux devient ensuite de plus en plus complexe chez les animaux plus évolués, avec une centralisation ( distinction de plus en plus marquée entre systèmes nerveux central et périphérique ) et une céphalisation ( concentration de la plus grande quantité de tissus nerveux vers la tête ). Cette évolution s'accompagne d'une importante augmentation du nombre des neurones , du nombre de synapses par neurone, et d'un enrichissement dans la capacité d'intégrer lers divers signaux .
I.2.1.2 Structure des neurones
Les neurones sont des cellules qui possèdent un ou plusieurs prolonbgements cytoplasmiques cytoplasmiques susceptibles de former une arborisation terminale plus ou moins développée et d'établir des contacts avec d'autres neurones ou des cellules effectrices ( synapses ).
I.2.1.2.1 Techniques d'étude structurale
Les cellules nerveuses étant le plus souvent des cellules ramifiées qui entretiennent des relations avec des nombreuses autres cellules, leur observation est rendue difficile par les dimensions du volume dans lequel elles sont déployées.
Histologie conventionnelle
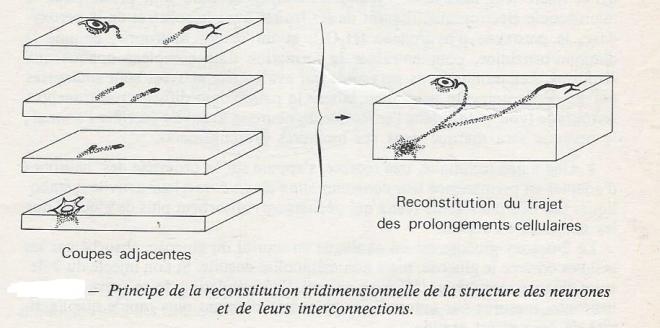
Celle-ci a permis une approche très minutieuse de la structure des neurones dès le début du XXième Siècle . Les coupes successives d'un même tissus ( coupes sériées) étaient répertoriées, colorées, puis observées une à une pour reconstituer la structure tridimensionnelle des neurones . Parmi les colorations utilisées, l'imprégnation argentique, de Golgi, est particulièrement intéressante , car, pour une raison inconnue , elle ne colore que 2 % environ des neurones dans un même tissus , ce qui permet de les isoler optiquement dans l'inextricable feutrage de fibres et de neurones qui les entourent.
Lésions des neurones
Les abblations, lésions et déafférentations de diverses régions du système nerveux permettent d'affiner l'étude topographique des connextions entre les diverses parties du système nerveux, en considérant les divers prolongements qui dégénèrent ( les fibres nerveuses sélectionnées régénèrent seulement si elles restent reliées au corps cellulaire.
Microscopie électronique
la microscopie électronique couplée à des techniques histochimiques, a permis de nouveaux progrès dans l'analyse des connections entre neurones, des relations de ceux-ci avec la neuroglie, des transmissions synaptiques. Il est possible de suivre très précisément , sur des coupes sériées, l'architecture complète d'un neurone.
- l'incorporation d'un précurseur radioactif permet d'effectuer des autoradiographies.
- l'injection de chlorure de cobalt dans le neurone peut être révélée, après que le produit ait diffusé, en traitant les coupes avec du sulfure d'ammonium , ce qui donne lieu à une précipitation métallique dense aux électrons.
- Des données fonctionnelles intéressantes peuvent être obtenues également en combinant l'expérimentation sur l'animal et l'étude ultrastructurale après traitement approprié des échantillons. Ainsi, il est maintenant possible de déterminer quels neurones ont été recrutés ( c'est à dire quels neurones ont été actifs ) dans une expérimentation et de tracer alors avec précision les voies nerveuses correspondantes. Plusieurs procédés ont été utilisés, les deux principaux étant les suivants :
l'endocytose qui fait suite à la libération des neurotransmetteurs peut être mise à profit en faisant incorporer des marqueurs à la cellule qui vient de libérer son médiateur . La péroxydase du Raifort est un de ces marqueurs : cette enzyme injectée à l'animal juste avant une stimulation des voies nerveusessera absorbée par endocytose au niveau des terminaisons des seuls neurones ayant libéré leur médiateur . Quand les coupes de tissus sont prêtes pour la microscopie électronique , il suffit de les traiter avec le substrat de la péroxydase, le peroxyde d'hydrogène ( H2O2 ) , et un réactif approprié, comme la diamino-benzidine, pour entraîner la formation d'un complexe opaque aux électrons . Les terminaisons nerveuses qui avaient été activées sont soulignées par un dépôt noir. On peut aussi laisser diffuser la péroxydase par transport rétrograde dans l'ensemble du neurone avant de sacrifier l'animal; le neurone sera marqué dans ses moindres prolongements.
- une autre technique s'appuie sur la propriété des neurones d'adapter en permanence leur consommation de glucose à leur activité métabolique . Les neurones actifs ( ceux qui déchargent ) absorbent plus de glucose que les neurones au repos . Le 2 desoxy-glucose est un analogue structural du glucose, absorbé par les cellules comme le glucose, mais non métabolisé ensuite . Si l'on injecte du 2 desoxy-glucose marqué au 14C à un animal, l'accumulation de ce sucre dans les neurones, mesurée par autoradiographie , sera d'autant plus rapide que l'activité de ceux-ci est grande.
I.2.1.2.2 Le corps cellulaire
Le corps cellulaire des neurones est limité par une membrane plasmique typique et renferme tous les organites essentiels d'une cellule animale nucléée : noyau, réticulum endoplasmique, appareil de Golgi, mitochondries, lysosomes, ainsi qu'une grande quantité de filaments microscopiques. Les neurofilaments sont les filaments intermédiaires du cytosquelette ( 6 à 10 nm ); des microtubules de 20 à 30 nm de diamètresont également présents. Ces structures ne sont pas particulières aux neurones , mais sont ici bien développées. Les " neurofibrilles " de ma microscopie optique correspondent à l'agrégation des filaments et tubules du cytoplasme , provoquée par les techniques de préparation. De même les corps de Nissl sont en fait les éléments du réticulum endoplasmique granulaire, très abondants dans les neurones ( en relation avec des synthèses protéiques abondantes ). En plus de ces organites , le neurone peut renfermer diverses inclusions, comme des globules lipidiques, des granules pigmentaires, du glycogène...
Le corps cellulaire ( ou péricaryon ) peut avoir , pour plus grande dimension, de 2µm ( certains neurones d'Insectes) à plus de 800 µm ( certains neurones d'aplysie ). L'importance relative du volume du corps cellulaire par rapport au volume de ses prolongements est aussi très variable : le rapport des deux volumes varie de 1 pour certains neurones globulaires à axones court , à plus de 10000 dans les neurones géants de certains Annélides polychètes.
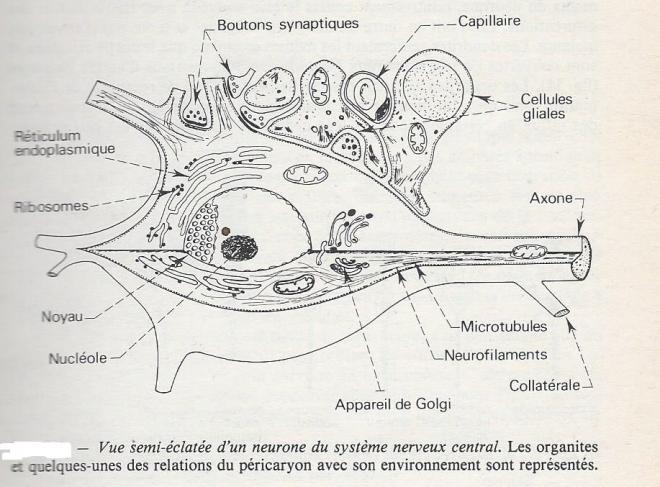
I.2.1.2.3 L'axone
L'axone est un prolongement du neurone , plus long que les autres et de diamètre relativement constant. Sa membrane est lisse , il possède quelques bifurcations et rencontre un petit nombre de terminaisons synaptiques. La plupart des neurones ont un axone unique . Le nomde cellules amacrines est donné aux neurones dont on ne peut pas dire s'ils ont un axone individualisé ( de telles cellules sont par exemple rencontrées dans la rétine de Mammifères ). La reconnaissance d'un axone est généralement plus aisée chez un Vertébré que chez un Invertébré, ce qui suggère que chez ces derniers , la différenciation entre dendrites et axones est moins marquée.
L'axone est pratiquement dépourvu de réticulum endoplasmique granulaire , formation qui assure les synthèses protéiques, alors que les neurofilamentset les microtubules sont très abondants et orientés parallèlement à l'axe du prolongement axonique. Cette disposition revêt une grande importance fonctionnelle au niveau des transports axoniques de substances.
I.2.1.2.4 Les dendrites
sur des bases cytologiques, on définit les dendrites comme des prolongements du neurone, relativements courts le plus souvent, avec fréquement des bifurcations . Elles ont en outre un diamètre irrégulier et n'ont pas d'enveloppe isolante . Les dendrites renferment les mêmes organites que le corps cellulaire et sont couvertes de très nombreux boutons synaptiques issus d'autres neurones . Les dendrites assurent essentiellement un rôle de réception de l'information mais on a pu montrer qu'elles pouvaient aussi transmettre des influx en certaines zones et leur définition formelle est peu précise.
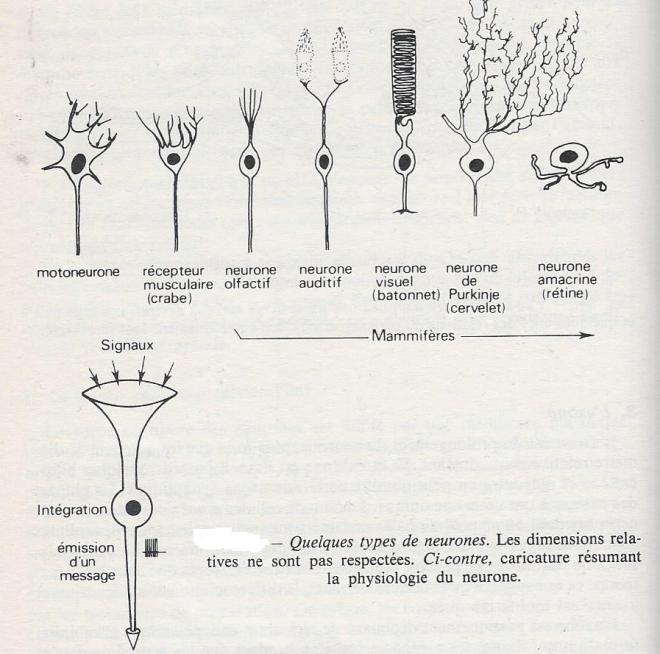
I.2.1.2.5 Diversité des types de neurones
L'anatomie microscopique des neurones est très variable selon l'organisme considéré, la position centrale ou périphérique du neurone , son rôle moteur ou sensoriel etc. La figure ci dessous donne un aperçu de cette variété. Les neurones sont unipolaires, bipolaires, multipolaires selon le nombre de prolongements issus du péricaryon. Les neurones unipolaires sont fréquents dans le système nerveux central des Invertébrés ( motoneurones et neurones intermédiaires)alors qu'ils sont rares dans celui des Vertébrés.
I.2.1.3 Physiologie des neurones
I.2.1.3.1 Besoins énergétiques des neurones
En plus d'une activité métabolique de base, nécessaire au simple renouvellement de ses constituants ( maintenance ), le neurone a des activités spécifiques telles la transmission des influx nerveux ou la synthèse et la libération de neuromédiateurs en quantités appréciables , qui sont coûteuses sur un plan énergétique . Dès lors on ne sera pas surpris par l'importance de la consommation énergétique du cerveau : chez les Mammifères , le cerveau prélève 20% du flux d'oxygène entrant dans l'organisme , alors que lui même ne représente que 2% du poids corporel ( chez l'homme ) On peut supposer qu'une part importante des besoins énergétiques du tissus nerveux est destinée à l'entretien de l'activité des pompes ioniques.
Contrairement à la plupart des tissus , le tissus nerveux ne consomme que du glucose en conditions normales ( il peut utiliser des corps cétoniques dans les cas de jeûne prolongé) et ne peut survivre en anaérobiose . C'est pourquoi les neurones du cerveau ne peuvent supporter sans séquelles pour l'individu une interruption du flux sanguin oxygéné pendant plus de quelques dizaines de secondes.
La consommation moyenne minimale ( repos ) de glucose par le cerveau de l'homme est de l'ordre de 6 gr par heure.
I.2.1.3.2 Transport axoplasmiques
La synthèse des divers constituants des neurones ne peut avoir lieu que dans le péricaryon, là ou est centrée toute l'infrastructure nécessaire aux processus synthétiques ( noyau, réticulum endoplasmique granulaire, appareil de Golgi...). L'axone, parfois très long ( c'est le cas pour les axones des motoneurones médullaires innervant les aorteils , par exemple ) , dépend ainsi du corps cellulaire pour renouveler ses constituants. On a pu vérifier qu'il existait en effet un transit de matière très important entre le corps cellulaire et la terminaison axonique des neurones, mais aussi un transport dirigé dans l'autre sens : de la terminaison axonique vers le corps cellulaire.
Technique d'étude
il suffit de ligaturer un nerf pendant quelques heures , ou quelques jours , pour observer l'accumulation des substances transportées en amont de la ligature . On mesure la concentration de la substance en question dans le tronçon de nerf situé en amont de la ligature et on compare la valeur trouvée à la concentration de la même substance mesurée dans la région homologue du nerf contralatéral. dans le nerf sciatique de Rat , on a pu mesurer de cette façon le transport d'enzymes comme la choline acétyltransférase ou l'acétylcholinestérase et de médiateurs peptidiques comme la somatostatine.
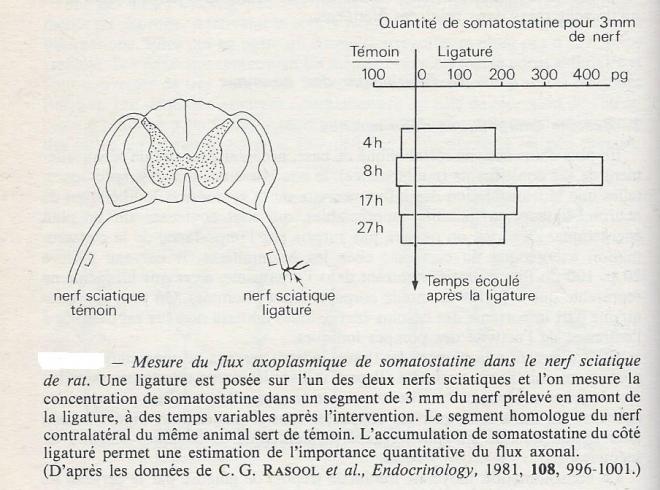
Transport orthograde
Le transport orthograde est dirigé du péricaryon vers la terminaison de l'axone. Dans les divers cas qui ont été étudiés , on a toujours trouvé deux modalités de ce transport de substances :
- Un transport lent ( 1 à 10 mm par jour ) constitué par un flux global de tout le cytoplasme axonique et assurant la régénération de tous les constituants de l'axone ( flux axoplasmique )
- Un système de transport rapide ( plusieurs dizaines ou centaines de mm par jour ), qui met en jeu les protéines fibrillaires de l'axoplasme ( transport axonal rapide ) .
Dans les cellules neurosécrétrices, cellules nerveuses ayant évolué vers la sécrétion hormonale ( sécrétion d'ADH ou d'ocytocine, par exemple ), le transport orthograde est très amplifié .
Transport axonique rétrograde
Le transport rétrograde implique également les protéines motrices de l'axone. Il permet un recyclage des matériaux prélevés par endocytose à l'extrémité de la terminaison axonique ( voir plus loin , synapse neuro-musculaire). C'est également par cette voie que certaines toxines ( tétaniques par exemple ) ou des virus ( rage, herpès ), gagnent le corps cellulaire du neurone.
I.2.1.3.3 l'influx nerveux ou signal nerveux
L'information unitaire que trnsmat un neurone est un influx nerveux. L'influx nerveux peut se définir comme une perturbation brusque , brève, stérotypée ( elle est toujours semblable, quelles que soient les causes de son apparition ) de l'équilibre dynamique qui existait entre le neurone et le milieu extracellulaire . Cette perturbation est un processus complexe affectant avant tout la membrane du neurone et se propageant rapidement. Il se produit des échanges ioniques très rapides , accompagnés de phénomènes électriques, métaboliques, thermiques...
Toutefois les modifications majeures accessibles à l'expérience , lors de la propagation du signal nerveux le long d'un axone, sont les variations de la perméabilité membranaire aux ions et les variations associées de l'équilibre électrique de la membrane ( potentiel d'action ) . Ces deux manifestations de la physiologie nerveuse ont permis d'analyser les mécanismes de genèse et de propagation du signal nerveux et donné lieu au développement de l'électrophysiologie dont l'intérêt majeur est de permettre une analyse temporelle très fine , avec une sensibilité de mesure intéressante.
I.2.1.4 Les cellules gliales
Les cellules non nerveuses qui accompagnent les neurones, entretiennent des relations avec le corps cellulaire ou avec l'axone. ces cellules forment la neuroglie , ou simplement la glie , environnement immédiat des neurones . Cette terminologie ne concerne pas , cependant, les cellules des vaisseaux sanguins , des trachées aériennes, des muscles, tissus conjonctifs, glandes, qui peuvent être associées au tissus nerveux. La quantité des cellules gliales est très supérieure à celle des neurones dans le système nerveux central de l'homme : 10 cellules gliales par neurone . Dans le système nerveux périphérique, les cellules de Schwann sont des cellules gliales. Les cellules gliales compartimentent le système nerveux, forment la gaine de myéline, ont un rôle phagocytaire et représentent un tissus de soutien pour les neurones. Les cellules gliales sont souvent l'intermédiaire , dans les échanges entre les neurones et les capillaires sanguins . Leur présence, enfin, limite la diffusion des signaux nerveux d'un neurone à ses voisins.
Il existe trois types principaux de cellules gliales chez les Vertébrés :
- Les astrocytes, cellules étoilées à prolongements nombreux, elles compartimentent le tissus nerveux et envoient des prolongements ( podocytes ) vers les vaisseaux sanguins et vers la pie mère.
- Les oligodendrocytes : cellules gliales les plus nombreuses , elles enroulent des feuillets cytoplasmiques autour des axones pour former la gaine de myéline , à la façon dont le couvercle d'une boîte de conserve de sardines s'enroule autour de l'ouvre boîte. - Les microglies sont des cellules ressemblant aux fibroblastes et dont le rôle phagocytaire est important
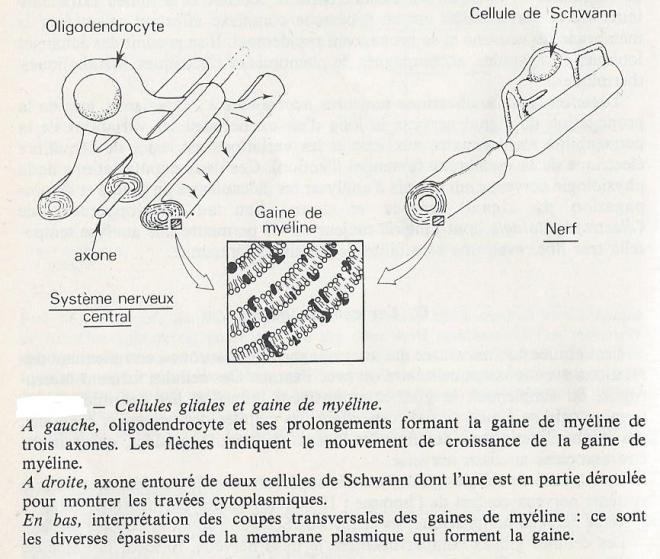
Les cellules de schwann qui entourent les axones des nerfs sont des cellules gliales très allongées ( 0,5 à 2mm), qui enveloppent plusieurs fois également l'axone pour former un manchon, la gaine de myéline ( 2 à 20 µm d'épaisseur ). Entre les cellules de Schwann successives , il existe un très petit espace au niveau duquel l'axone n'est plus isolé de l'environnement : cet espace est le neud de ranvier , dont l'importance est considérable pour la propagation rapide de l'influx nerveux ( propagation saltatoire ).
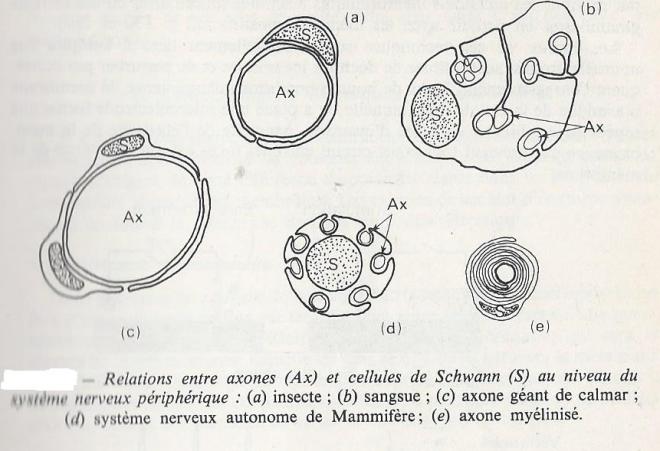
I.2.2 La conduction et transmission de l'influx nerveux
1.2.2.1 L'influx nerveux
1.2.2.1.1 Phénomènes électriques membranaires
1) Le potentiel de repos
Entre deux faces de l'axolemme , une différence de -60 à -90 mV, due à des différences de concentration ionique.
Mécanisme : dans la membrane de l'axolemme , mécanisme dénommé " pompe à sodium et à potassium" ( protéine) qui maintient une concentration élevée en Na+ à l'extérieur et une concentration élevée en K+ à l'intérieur.
Le mécanisme de pompage dépend naturellement du métabolisme cellulaire.
Dans cette même membrane existent des protéines canaux ( ex : canal à Na+ et canal à K+ ) qui à l'état de repos sont fermées.
Les protéines canaux : deux propriétés essentielles :
- Sélectivité.
- Effet de porte.
La sélectivité : la perméabilité sélective existe existe mais forte variation d'un type de canal à l'autre . Elle est rarement absolue.
Ex.
Type peu sélectif : 85 ions Na pour 100 ions K, canal activé par l'acétylcholine .
Type très sélectif : 7 ions Na pour 100 ions K, canal potassium, diamètre plus faible et moins d'eau , pourtant l'ion Na est 30% plus petit !
Les mécanismes généraux permettant cette sélection mettraient en jeu des interactions d'une part, entre les ions et d'autre part, les éléments plus ou moins hydratés de la structure du canal.
L'effet de porte : 2 formes à ce mécanisme :
- Mouvements de la porte due à l'effet de différences de potentiel membranaire , on parlera, dans ce cas, de canaux " voltage dépendant" ;
- Mouvements de la porte dus à des effets chimiques ( médiateurs). Dans ce cas , canaux " chimio-dépendants " ( sur la membrane post-synaptique).
Tous ces mouvements sont occasionnée par des changements de forme des protéines sollicitées. ( Déplacement des parties de la molécule qui sont situés en des points clés).
2) Le potentiel d'action
- Un stimulus ( excitation électrique, par exemple ) modifie la polarisation de la membrane . Elle se dépolarise temporellement et localement . C'est le potentiel local. Cette excitation a provoqué une modification de la conformation des protéines canaux. Lorsque le potentiel de membrane diminue , les canaux s'ouvrent ( le canal sodium rapidement, le canal potassium plus lentement.
- Si le stimulus atteint une certaine densité , la dépolarisation de l'axolemme se propage le long de la fibre nerveuse et il est possible d'enregistrer une courbe ou potentiel d'action ( voir schéma ).

1.2.2.1.2 La conduction de l'influx nerveux
La membrane des cellules est si fine que la différence de 70mV de part et d'autre , au repos , engendre un champs électrique intense de l'ordre de 100 KiloV par centimètre.
Les dipôles électriques des protéines du canal sodium tendent à s'aligner dans le champs électrique membranaire ( idem pour dipôle magnétique s'alignant dans les lignes de force d'un champs magnétique).
Dès lors , il est aisé de comprendre que les modifications de l'intensité du champ membranaire puissent faire passer le canal de la conformation fermée à la conformation ouverte.
Quand la face interne de la membrane est rendue plus positive par la pénétration des premiers ions Na , les canaux sodiques tendent à rester dans la configuration ouverte un temps plus long, C'est le processus connu sous le nom d'activation des canaux sodium.
Processus de fermeture des canaux sodiques : inactivation sodique
La différence de potentiel qui a provoqué l'ouverture temporaire des canaux sodiques entraine finalement une configuration fermée spéciale , différente de celle qui caractérise l'état de repos du canal . Cet état d'inactivation s'installe plus lentement que l'activation , de sorte que les canaux restent ouverts un cours laps de temps avant d'être fermé par inactivation. Après cette phase d'inactivation , les canaux reviennent à l'état de repos . ensuite ce sont les canaux Potassium qui s'ouvrent, provoquant ainsi une repolarisation de la membrane et une hyperpolarisation temporaire.
L'onde de dépolarisation va se propager évidemment de manière différente dans les fibres nerveuses amyéliniques et dans les fibres myéliniques.
- - Polarisation dans les fibres amyéliniques : Un courant électrique s'établit entre le zone inactive ( polarisée ) et la zone active ( dépolarisée) . Il se forme ainsi des courants locaux ou circuits locaux qui assurent la propagation de l'influx nerveux de proche en proche , à partir de la zone active.
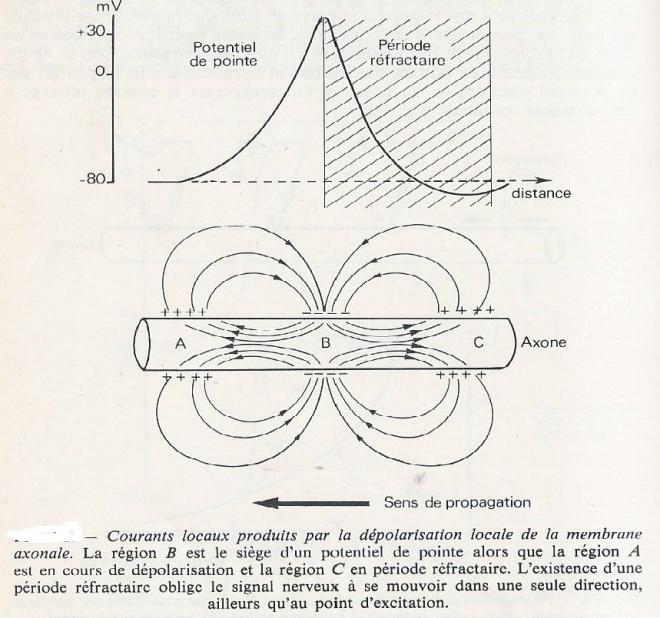
En théorie, le sens de la conduction est indifférent, il est bidirectionnel, mais il faut savoir que la zone qui vient d'être dépolarisée connaît une brève période réfractaire ( la zone n'est plus excitable), ce qui empèche la propagation de la dépolarisation dans le sens de cette zone réfractaire, la conduction sera donc pratiquement unidirectionnelle et la dépolarisation se déplacera dans le sens inverse
En effet : Après la repolarisation, la membrane demeure inerte un certain temps ; les canaux à Na+ ne peuvent pas s’ouvrir (période réfractaire).
 -
-
- - La propagation dans les fibres à myéline
La théorie saltatoire. : la gaine de myéline isole l'axone , sauf aux noeuds de Ranvier. Lorsque la membrane se dépolarise à un noeud, le courant local ne pourra exciter la membrane qu'au niveau du noeud suivant, d'où le nom de théorie saltatoire. Cette conduction saltatoire est plus rapide et plus économique.

il suffit d'un flux relativement faible d'ions sodium et potassium pour dépolariser la région membranaire correspondant à un noeud.
Dès lors, il suffit d'une faible quantité d'énergie pour rechasser les ions sodium hors de l'axone et réintroduire les ions potassium lors de la repolarisation de la membrane. Dans le cas d'une fibre amyélinique , c'est la totalité de la membrane axonique qui doit se dépolariser puis se repolariser.
Donc, plus les noeuds de Ranvier sont espacés, plus le seuil de stimulation est bas.
I.2.2.1.3 Conduction dans le nerf
Le nerf est un système qui permet de retrouver les principales caractéristiques de l'axone :
- seuil d'excitation : le potentiel d'action nerveux étant la somme des potentiels individuels des fibres qui constituent le nerf , il n'y a pas de phénomène de tout ou rien, mais une lente gradation de l'amplitude , quand le stimulus appliqué augmente , par suite du recrutement d'un nombre croissant d'axones dont la position dans le nerfet l'excitabilité sont variables.
- Périodes refractaires absolues et relative : Le profil du potentiel d'action nerveux présente souvent plusieurs sommets, par suite de la présence de fibres dont l'excitabilité et la vitesse de propagation des signaux sont variables. Les fibres de fort calibre sont à la fois plus excitables et conduisent l'influx plus rapidement : elles sont à l'origine du premier sommet enregistré, alors que les signaux transmis par les autres fibres apparaissent avec un retard sur l'enregistrement.
Date de dernière mise à jour : 30/03/2018
Ajouter un commentaire