

Physiologie animale 13
2. Notion de bioénergétique des organismes
2.1 Quotient respiratoire-coefficients thermiques de l'O2
Comme nous l'avons vu, l'oxydation complète du glucose en aérobie produit du CO2 et de l'ATP et consomme de l'oxygène. La réaction nette peut s'écrire comme suit:
glucose + 36 (ADP + Pi) + 6 O2 ![]() 6 CO2 + 6 H2O + 36 ATP
6 CO2 + 6 H2O + 36 ATP
On remarque que l'utilisation d'une mole de glucose requière 6 moles d'oxygène et produit 6 moles de CO2 Dans ce cas le rapport du CO2 produit sur l'oxygène consommé (quotient respiratoire: QR) est de 1. Le résultat sera le même si le glucose est brûlé totalement dans un calorimètre. Dans un tel système, l'énergie résultant de l'oxydation d'un produit détermine une production de chaleur mesurable. Ce produit a donc une certaine valeur calorique. Les lipides et les hydrates de carbones sont oxydés en CO2 et H2O dans les cellules comme dans un calorimètre, la valeur calorique physiologique sera donc égale à la valeur calorique physique. Ce n'est pas le cas pour les protéines dont le catabolisme aboutit, chez bon nombre d'espèces à des produits d'excrétion ayant encore une certaine valeur calorique (urée, acide urique ou autres, cfr. plus loin: excrétion azotée). Pour les protéines, la valeur calorique physiologique est donc inférieure à la valeur physique (4,3 kcal/g contre 5,3). Le tableau 3-1 donne pour les trois catégories de produits utilisés par les animaux, la valeur calorique physiologique, la quantité d'oxygène consommée (QO2) et le quotient respiratoire. Il ressort de l'analyse de ces résultats que: 1) les graisses sont nettement plus énergétiques que les protéines ou les hydrates de carbones, 2) le quotient respiratoire est différent pour chacun des types de produit oxydé. La mesure du QR va donc fournir une indication sur le type de composé utilisé essentiellement à un moment donné dans le métabolisme énergétique, 3) les quantités d'énergie produites par litre d'oxygène consommé (coefficients thermiques de l'oxygène ou équivalents caloriques) restent voisines quel que soit le type de composé oxydé. Ce constat est d'importance puisqu'il est à la base du développement de l'essentiel de la bioénergétique. La mesure de la consommation d'oxygène (QO2) +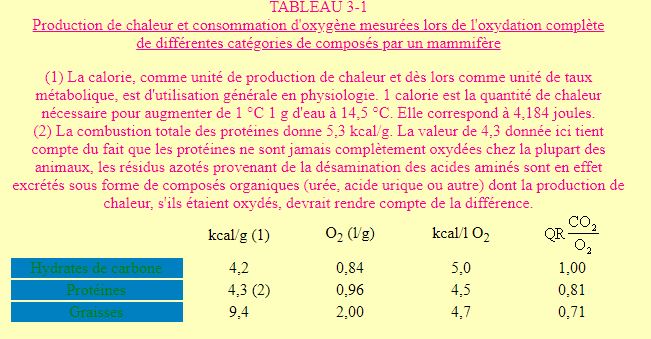 peut en effet être mise directement en relation avec la quantité d'énergie requise par un animal à un moment donné. On pourrait, en principe, faire appel aussi à la mesure du CO2 produit. Les erreurs possibles avec le CO2 sont cependant beaucoup plus importantes qu'avec l'O2 . Une des raisons majeures est que les organismes possèdent un pool de CO2 très important impliqué dans un système tampon dont les variations, en plus ou en moins, peuvent dépendre de nombreux facteurs indépendants du métabolisme (voir plus loin: balance acide - base). L'approche bioénergétique des activités biologiques par mesure de production de CO2a donc été très rapidement abandonnée, sauf dans des cas bien particuliers.
peut en effet être mise directement en relation avec la quantité d'énergie requise par un animal à un moment donné. On pourrait, en principe, faire appel aussi à la mesure du CO2 produit. Les erreurs possibles avec le CO2 sont cependant beaucoup plus importantes qu'avec l'O2 . Une des raisons majeures est que les organismes possèdent un pool de CO2 très important impliqué dans un système tampon dont les variations, en plus ou en moins, peuvent dépendre de nombreux facteurs indépendants du métabolisme (voir plus loin: balance acide - base). L'approche bioénergétique des activités biologiques par mesure de production de CO2a donc été très rapidement abandonnée, sauf dans des cas bien particuliers.
2.2. Q02 et dépense énergétique
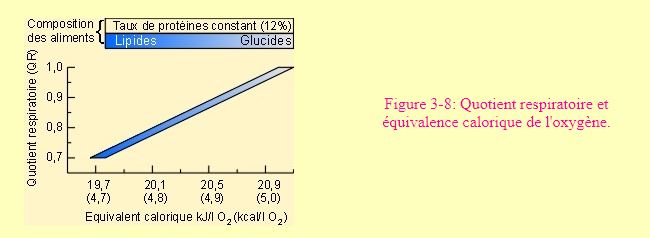
L'approche bioénergétique des activités physiologiques organismiques s'est largement développée de la fin du siècle dernier jusqu'au milieu des années 60. L'activité de recherche dans ce domaine s'est quelque peu ralentie par la suite. Ceci est du en partie, aux limitations des techniques utilisées par rapport aux exigences des problèmes qui restent à éclaircir. Comme nous l'avons déjà signalé, l'essentiel de la bioénergétique au niveau organismique repose sur le fait que les coefficients thermiques de l'oxygène pour les trois grandes catégories de produits oxydés par les animaux sont voisins, variant en gros de 4,5 à 5 kcal/l O2 (figure 3-8). En pratique, on considère d'ailleurs souvent qu'un litre d'O2 consommé correspond à une production d'énergie de 4,8 kcal ou de 20,1 kJ.
Il devient donc possible d'estimer la dépense énergétique par une simple mesure de consommation d'oxygène. Chez l'animal, une telle mesure peut être relativement facile dans certaines conditions. Elle peut devenir extrêmement difficile et peu fiable dans d'autres. Il devient dès lors parfois délicat de rapporter une consommation d'oxygène mesurée ou une variation de celle-ci à la dépense énergétique engendrée par une activité physiologique donnée. Des difficultés majeures viennent notamment du stress de manipulation ou de confinement qui entoure la prise de mesure.
D'autres paramètres bioénergétiques ont été extrapolés à partir de mesures de consommation d'oxygène et de mesures de quantités d'aliments ingérés, éliminés et digérés. Ces mesures sont souvent peu fiables étant donné les difficultés à effectuer des bilans nutritionnels corrects chez certaines espèces, notamment aquatiques. Ces paramètres ne seront pas envisagés ici.
L'approche bioénergétique des activités physiologiques a, malgré ces limitations, permis la mise en évidence d'une série de relations et de concepts de base dont certains sont brièvement résumés ci-après. Cette approche a jusqu'à présent montré, pour l'essentiel, que la demande énergétique des organismes pouvait être fonction de leur masse et de leur niveau d'activité. D'autres facteurs comme la température, la teneur en oxygène du milieu ou certaines hormones interviennent également de façon notable dans la demande énergétique globale. Ces facteurs seront discutés dans les chapitres qui leurs sont spécifiquement consacrés (cfr. plus loin: hormones et livre III écophysiologie: température/disponibilité de l'oxygène).
2.3. Métabolisme énergétique et masse des organismes
Comme le montre la figure 3-9, établie à partir de données concernant des mammifères, il existe une relation de proportionnalité entre la consommation d'oxygène (et donc le métabolisme énergétique) et la masse d'un organisme. Ainsi, la consommation d'oxygène spécifique (par unité de poids) de la musaraigne est quelque 100 fois supérieure à celle de l'éléphant (7,4 contre 0,07 l O2/kg/h). Si les données de la figure 3-9sont exprimées dans un système de coordonnées logarithmiques au lieu de semi-logarithmiques, on s'aperçoit qu'elles tombent sur une ligne droite donnée par l'équation VO2 = 0,676Mc0,75 (VO2 étant la consommation d'oxygène et Mc la masse corporelle).
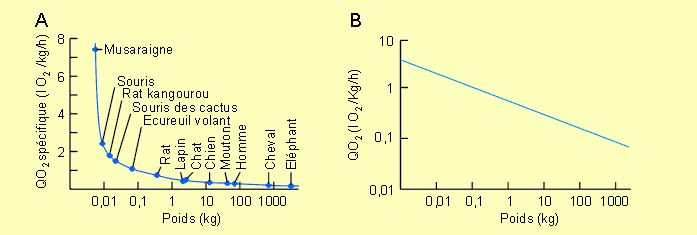
Figure 3-9: Relation entre la consommation d'oxygène (QO2) et le poids chez différents mammifères. A: QO2 spécifique, coordonnées semi-logarithmiques. B: QO2, coordonnées logarithmiques.
Des mesures de consommation d'oxygène en fonction de la masse ont été effectuées sur de nombreuses espèces, vertébrés comme invertébrés. En fonction des groupes étudiés, les données tendent toujours à s'aligner sur des droites dont la pente est toujours proche de 0,75 (figure 3-10).
Ces faits, mis en évidence par Hemmingsen en 1960, montrent que la relation qui existe entre la masse d'un organisme et sa demande énergétique s'est maintenue pratiquement constante depuis les unicellulaires jusqu'aux vertébrés les plus évolués. Ce type de relation n'est pas encore éclairci à l'heure actuelle. Il est clair que de nombreux processus, comme la dissipation de chaleur, sont fonction de la surface d'échange. Il est par ailleurs clair qu'un grand rapport surface/volume facilitera les phénomènes dissipatifs. Dans ce cadre on peut comprendre qu'un petit animal ayant une surface dissipative proportionnellement plus grande qu'un gros ait une demande énergétique plus élevée. Certains ont dès lors voulu mettre ces relations en rapport, chez les homéothermes, avec la dissipation de chaleur, le travail cardiaque pour faire circuler le sang ou les possibilités d'échanges gazeux au niveau des surfaces respiratoires. Chez ces espèces, il semble bien que la dissipation calorique engendrée par le maintien d'une température corporelle supérieure à la température extérieure intervienne dans les paramètres à prendre en considération. En effet, l'équation VO2 = 0,676Mc 0,75 établie pour des mammifères euthériens (température ±37 °C) devient VO2 = 0,409Mc0,75 pour des mammifères marsupiaux (température ±35 °C) et VO2 = 1,11Mc0,724 pour des passereaux (température ±41 °C). La pente reste donc toujours la même mais la demande énergétique sera systématiquement plus élevée chez les mammifères euthériens que chez les marsupiaux et chez les oiseaux que chez les mammifères euthériens, en fonction de la température corporelle.
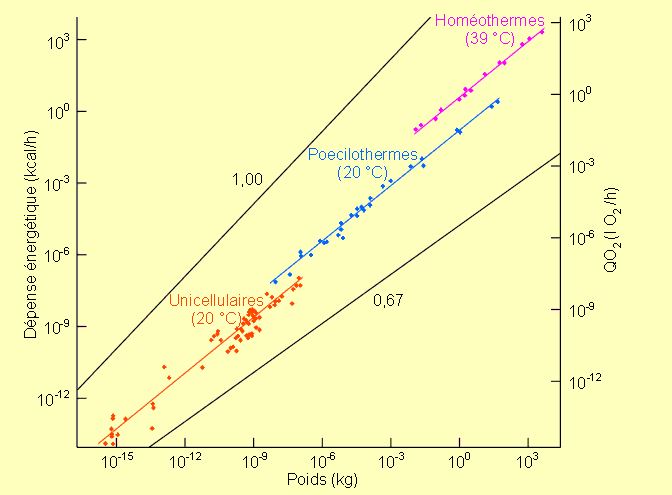
Figure 3-10: Consommation d'oxygène (QO2 ) et dépense énergétique en fonction du poids chez différents organismes, unicellulaires, poecilothermes ou homéothermes (coordonnées logarithmiques).
Le fait que la pente de la droite qui définit les relations masse - demande énergétique soit pratiquement la même chez les unicellulaires et chez les pluricellulaires les plus complexes indique cependant qu'il faut chercher l'explication de ce type de relation au niveau cellulaire et non au niveau de systèmes physiologiques organismiques complexes comme certains ont voulu le faire.
2.4. Métabolisme énergétique et niveau d'activité
Chez tous les organismes, la demande énergétique est directement fonction du niveau d'activité. Partant d'une valeur de repos, en rapport avec un métabolisme dit de base ou standard, elle est d'autant plus élevée que l'activité développée est grande.
Chez l'homme, on distinguera ainsi un métabolisme de base (ou standard) d'un métabolisme d'activité, correspondant à différents niveaux d'exercice ou de travail. Chez l'animal on fera plus souvent intervenir la notion de métabolisme de transport, celui-ci correspondant à l'activité métabolique à mettre en oeuvre pour se déplacer. Le métabolisme de base (standard, de repos) devrait être dérivé de la consommation d'oxygène d'un sujet lorsque tous ces organes sont au repos complet. Une telle mesure n'est pas toujours aisée à obtenir, surtout chez l'animal.
Chez l'homme, le métabolisme de base est obtenu par mesure de la consommation d'oxygène chez un sujet couché, détendu mais éveillé, en équilibre avec une température ambiante de 19 °C et après 12 à 18 heures de jeûne. Cette période de jeûne est nécessaire pour éviter l'augmentation d'activité métabolique due aux processus digestifs. On a en effet remarqué que le métabolisme peut augmenter de 10 à 35 % entre 3 et 6 heures après un repas. Cet effet, en rapport avec les processus digestifs, est plus marqué pour les protéines que pour les lipides et les glucides. Il est nommé "action dynamique spécifique des aliments".
Le métabolisme de base chez l'homme est variable avec l'âge, passant de quelque 55 kcal/m2/h à 5 ans à 35 kcal/m2/h à 50 ans. Il paraît également légèrement inférieur après adaptation à des températures extérieures élevées.
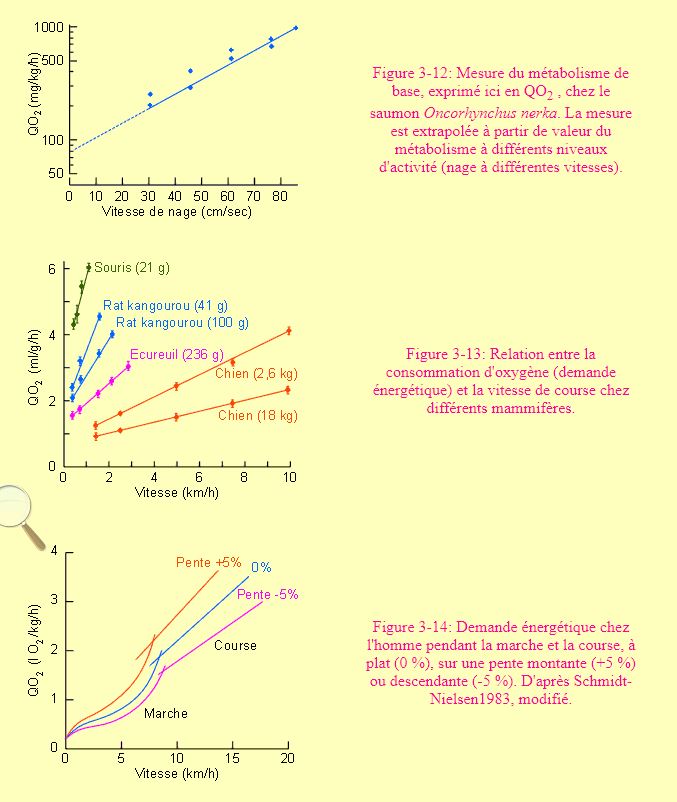
Chez l'animal, une estimation du métabolisme de base ne peut être obtenue comme chez l'homme. Certains auteurs ont approché le problème en utilisant des animaux anesthésiés. La méthode la plus courante à l'heure actuelle consiste à mesurer la demande énergétique à différents niveaux d'activité et à extrapoler une valeur au niveau 0 (figure 3-12).
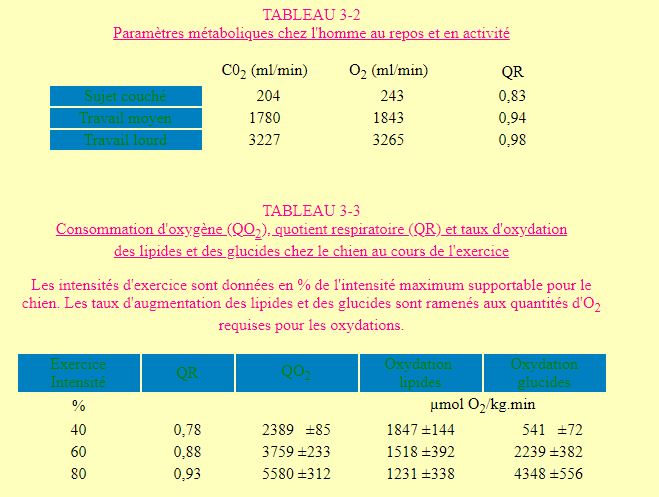
Le travail musculaire est de loin le facteur le plus important dans l'augmentation du métabolisme observée lors du passage du repos à l'activité. Chez l'homme, la demande énergétique peut ainsi passer de ± 2000 kcal/j en condition de repos à 6 - 7000 kcal/j lors de périodes de travail intense. Ce passage est toujours accompagné d'une augmentation de la consommation d'oxygène. Celle-ci va de pair avec une augmentation de production de CO2 ainsi qu'avec une augmentation du quotient respiratoire dont la valeur se rapproche d'autant plus de 1 que le travail est important (tableau 3-2). L'évolution de ce dernier paramètre indique que les glucides sont oxydés préférablement aux lipides lors de l'effort. Il en va de même chez le chien où on a pu mettre en évidence lors de l'augmentation de l'intensité d'un exercice (course), une augmentation de la consommation d'oxygène et du quotient respiratoire ainsi qu'une diminution du taux d'oxydation des lipides au profit de celle des glucides ( tableau 3-3).
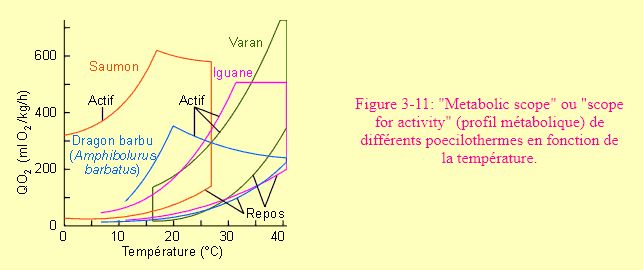
 Chez l'animal, le métabolisme d'activité correspond souvent à un déplacement plutôt qu'à un travail. On parlera d'ailleurs souvent de métabolisme de transport. L'augmentation de consommation d'oxygène observée lors du passage du repos à un déplacement à allure "normale" peut être éminemment variable. Elle est en général de 8 à 10 fois supérieure. Les différents niveaux de demande énergétique que peut manifester un animal déterminent ce que les anglo-saxons ont appelé son "metabolic scope" ou "scope for activity". Peut-être pourrait-on traduire ces expressions par "profil métabolique". Celui-ci est évidemment variable d'animal à animal et est largement fonction de différents paramètres (tels que la température chez les poecilothermes par exemple, figure 3-11).
Chez l'animal, le métabolisme d'activité correspond souvent à un déplacement plutôt qu'à un travail. On parlera d'ailleurs souvent de métabolisme de transport. L'augmentation de consommation d'oxygène observée lors du passage du repos à un déplacement à allure "normale" peut être éminemment variable. Elle est en général de 8 à 10 fois supérieure. Les différents niveaux de demande énergétique que peut manifester un animal déterminent ce que les anglo-saxons ont appelé son "metabolic scope" ou "scope for activity". Peut-être pourrait-on traduire ces expressions par "profil métabolique". Celui-ci est évidemment variable d'animal à animal et est largement fonction de différents paramètres (tels que la température chez les poecilothermes par exemple, figure 3-11).
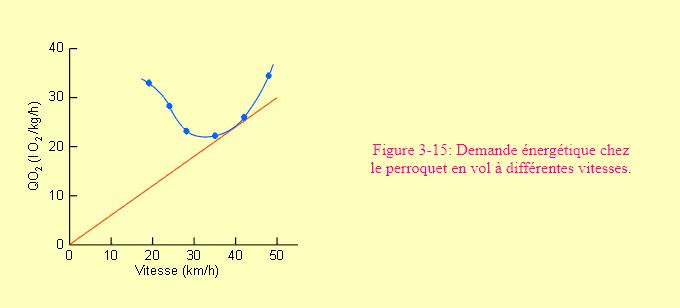
 En général, dans le métabolisme de transport, la demande énergétique est directement proportionnelle à la vitesse de déplacement. C'est en tout cas le cas pour la nage (figure 3-12) et la course (figure 3-13). La marche et le vol donnent des résultats un peu différents, avec des courbes d'allures complexes (figures 3-14 et 3-15). Dans le cas du vol, on voit nettement que certaines vitesses coûtent plus cher que d'autres. L'animal sélectionnera souvent une vitesse de déplacement dans un "preferendum" de vitesses auxquelles il doit faire face à une moindre demande énergétique. La figure 3-13 montre par ailleurs que pour une même vitesse de déplacement, la demande énergétique sera d'autant plus grande que l'animal est petit. Un phénomène qui s'explique aisément compte tenu du nombre de mouvements plus grand que devra faire un petit animal pour parcourir une distance donnée dans le même temps qu'un grand.
En général, dans le métabolisme de transport, la demande énergétique est directement proportionnelle à la vitesse de déplacement. C'est en tout cas le cas pour la nage (figure 3-12) et la course (figure 3-13). La marche et le vol donnent des résultats un peu différents, avec des courbes d'allures complexes (figures 3-14 et 3-15). Dans le cas du vol, on voit nettement que certaines vitesses coûtent plus cher que d'autres. L'animal sélectionnera souvent une vitesse de déplacement dans un "preferendum" de vitesses auxquelles il doit faire face à une moindre demande énergétique. La figure 3-13 montre par ailleurs que pour une même vitesse de déplacement, la demande énergétique sera d'autant plus grande que l'animal est petit. Un phénomène qui s'explique aisément compte tenu du nombre de mouvements plus grand que devra faire un petit animal pour parcourir une distance donnée dans le même temps qu'un grand.
 On peut par ailleurs évaluer des coûts de locomotion en O2 consommé par unité de poids, soit pour une distance parcourue unitaire, soit pour une vitesse unitaire. De tels calculs sont repris figure 3-16 pour des animaux dont le mode de déplacement habituel est la nage ou le vol ainsi que pour la course. Comme on peut le remarquer la nage est moins coûteuse que le vol et c'est la course qui est le système le plus coûteux.
On peut par ailleurs évaluer des coûts de locomotion en O2 consommé par unité de poids, soit pour une distance parcourue unitaire, soit pour une vitesse unitaire. De tels calculs sont repris figure 3-16 pour des animaux dont le mode de déplacement habituel est la nage ou le vol ainsi que pour la course. Comme on peut le remarquer la nage est moins coûteuse que le vol et c'est la course qui est le système le plus coûteux.
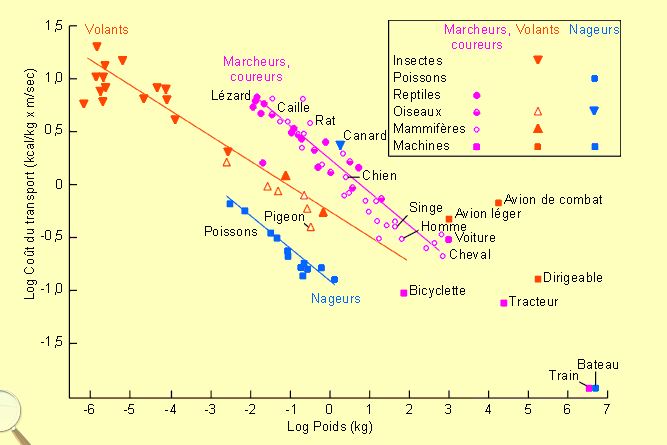
Il n'y a rien d'étonnant à ce que pour une même vitesse, la nage demande moins d'énergie que le vol. Le poisson n'a pas à supporter son propre poids; ce que l'oiseau doit faire. La viscosité et la densité de l'eau sont cependant largement supérieures à celles de l'air. Le poisson doit dès lors vaincre une résistance à la pénétration dans le milieu plus importante que celle de l'oiseau. Dans ce cadre, on remarquera que les animaux aquatiques sont les seuls à avoir évolué vers des morphologies tenant largement compte de ces problèmes. Ainsi, ils ont en général des profils donnant des coefficients de pénétration nettement inférieurs à ceux des animaux terrestres pour lesquels la faible résistance de l'air rend le problème moins essentiel.
Ce type de recherches a donné des résultats curieux qui restent à l'heure actuelle à l'état de constats inexpliqués. Ainsi par exemple chez les espèces arboricoles, le déplacement par "brachiation" a un coût spécifique plus élevé que la course sur les branches. De même, le déplacement par bonds successifs des grands kangourous s'effectue à un coût spécifique plus élevé que la course jusqu'à des vitesses de 18 km/h. A vitesse plus élevée le déplacement par bonds coûte significativement moins cher que la course (Dawson et Taylor, Nature 246, 313, 1973). Il est intéressant de remarquer dans ce cadre que les grands kangourous se déplacent normalement sur longues distances à des vitesses de 25 à 30 km/h. Ce phénomène curieux ne paraît vrai que pour les grands kangourous, on ne le retrouve pas chez les petites espèces, kangourous ou rongeurs, qui pratiquent également le déplacement par bonds.
On pourrait s'étonner de ce que le moyen de locomotion le plus coûteux soit la course. Ce phénomène est du au fait que les mouvements et la façon dont les muscles doivent être utilisés pour réaliser ces mouvements sont différents de ce qu'ils sont pour le vol ou la nage et sont de toutes façons moins efficaces. La course est caractérisée par une succession de montées et de descentes du centre de gravité (bonds). Le contrôle des " montées" et des "retombées" nécessite une dépense d'énergie qui n'a rien à voir avec la propulsion. Cette façon improductive de dépenser de l'énergie musculaire dans la course pour contrecarrer les influences gravitationnelles et décélérer les retombées a été appelée "production de travail négatif". C'est ce travail négatif qui rend compte du coût élevé de la course.
La production de travail négatif dans la course est minimisée par l'utilisation d'éléments élastiques plutôt que musculaires. Ces éléments peuvent emmagasiner une partie de l'énergie de décélération à la descente et la restituer au départ du bond suivant. Dans ce cadre, les pattes des animaux coureurs sont remarquables de minceur. Elles ne contiennent que très peu de muscles et sont constituées essentiellement d'os et de forts tendons élastiques. On considère généralement que les différentes attitudes de déplacement adoptées par certains animaux en fonction de leur vitesse peuvent correspondre à une optimisation de l'utilisation de l'énergie stockée dans les structures élastiques (marche, trot, galop chez le cheval).
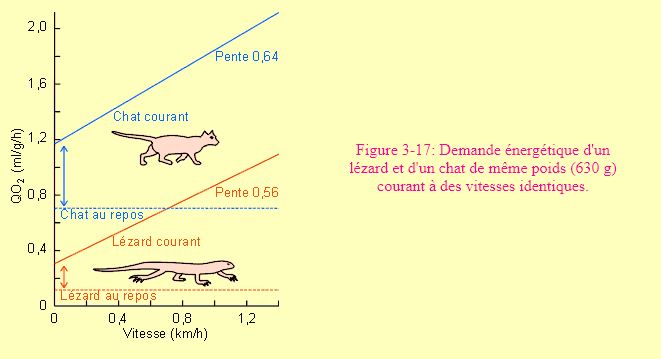
Il est par ailleurs clair que le coût de locomotion sera fonction, pour un même poids et une même vitesse, de la morphologie des animaux. Seront ainsi pris en compte notamment le plus ou moins bon hydrodynamisme chez les nageurs ou l'organisation des structures locomotrices chez les coureurs. Ainsi, la course à une même vitesse coûte moins cher à un lézard qu'à un chat de même poids (figure 3-17). Ce qui varie ici essentiellement n'est pas l'augmentation de demande énergétique avec l'augmentation de vitesse; la pente de la droite n'est en effet que peu inférieure chez le lézard. Ce qui varie grandement c'est le coût du passage du repos à l'activité de course (intersection de la droite avec l'axe Y). Cette différence serait en rapport avec ce que l'on a appelé les "coûts posturaux" de la course. C'est-à-dire les coûts nécessaires pour faire se mouvoir différentes parties vers le haut ou le bas, l'efficacité des systèmes élastiques etc... De plus amples informations à ces sujets relèvent plus de la morphologie fonctionnelle que de la physiologie.
Date de dernière mise à jour : 17/09/2018
Ajouter un commentaire