
ORGANOGENESE VERTEBRES10
Organognèse vertébrés 10
3.4.4 La somatopleure
Celle-ci est essentiellement à l’origine de l’endosquelette des membres de Vertébrés.
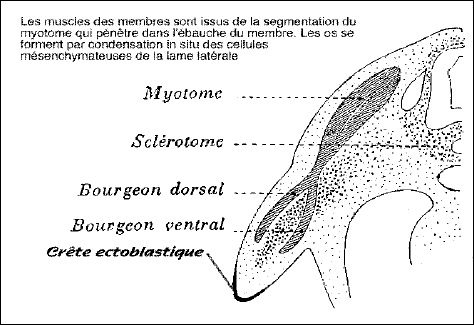
Le squelette et la musculature (striée squelettique) des membres proviennent des sclérotomes et myotomes de certains somites, mais entre aussi dans la composition des membres une portion du feuillet pariétal du mésoderme latéral et, bien entendu, le membre est recouvert de peau (composantes mésodermique et ectodermique).
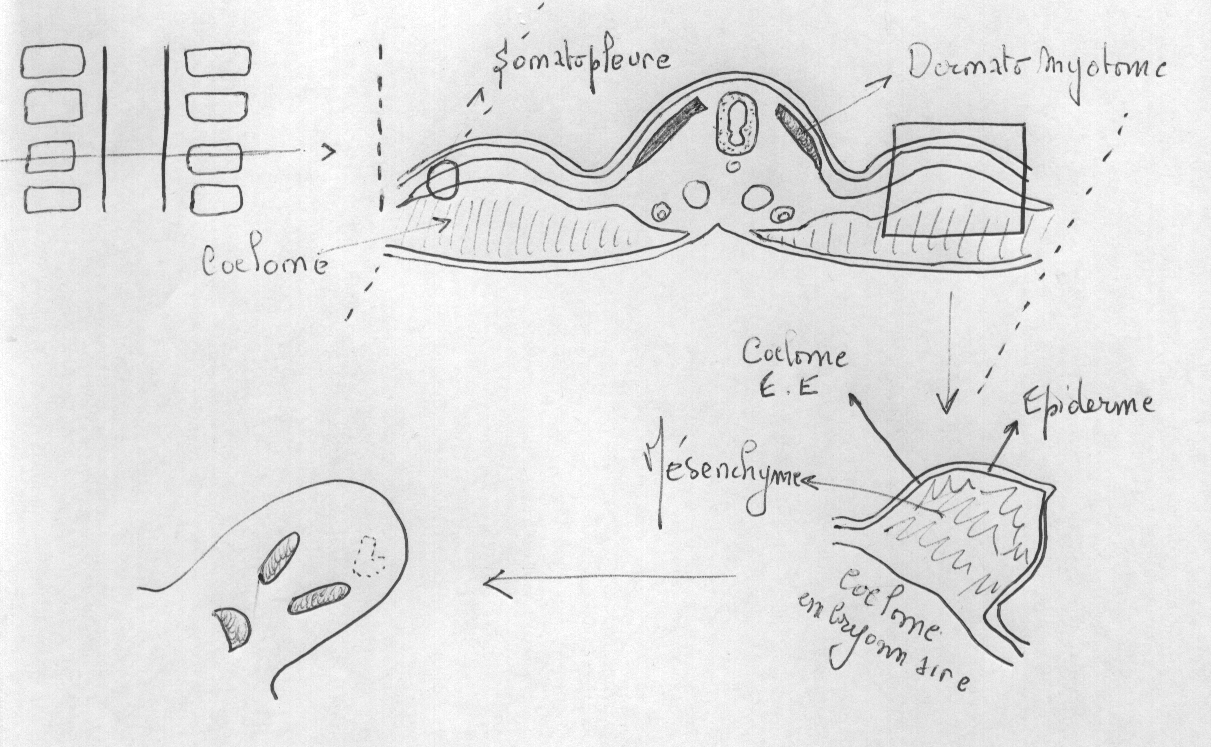
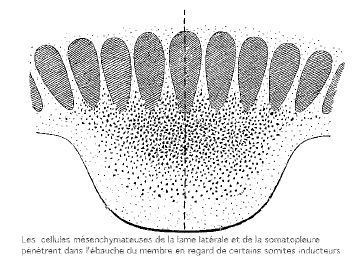
Le premier indice de la formation d'un membre est un épaississement de la partie supérieure du feuillet pariétal du mésoderme latéral. Les cellules de cet épaississement se détachent du feuillet et forment une masse mésenchymale qui migre latéralement sous la peau en formation. Chez les amphibiens, ces épaississements se retrouvent en positions des futurs membres: aux niveaux brachial et pelvien, bilatéralement. Chez les autres tétrapodes, ils peuvent se produire sur presque toute la longueur du tronc mais ne continuent de se développer qu'aux endroits des membres, régressant ailleurs. Les cellules des sclérotomes et des myotomes migrent vers les épaississements qui font maintenant saillie à la surface: ce sont les bourgeons des membres. Les
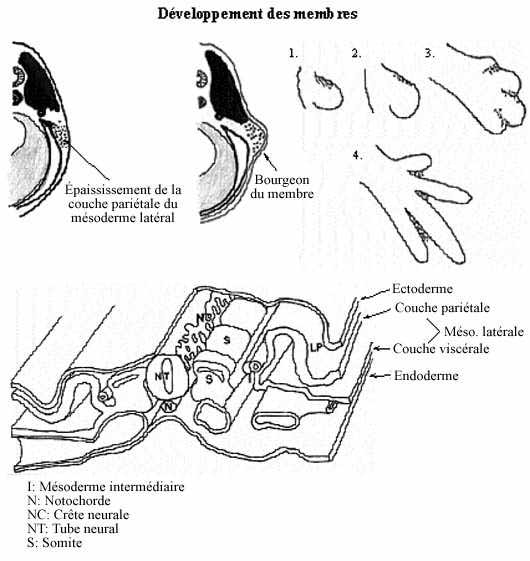
cellules du bourgeon se divisent plus rapidement que celles adjacentes du tronc de l'embryon. À l'intérieur du bourgeon, les cellules se divisent plus rapidement dans la portion distale que proximale, le bourgeon croissant ainsi plus rapidement par son extrémité.
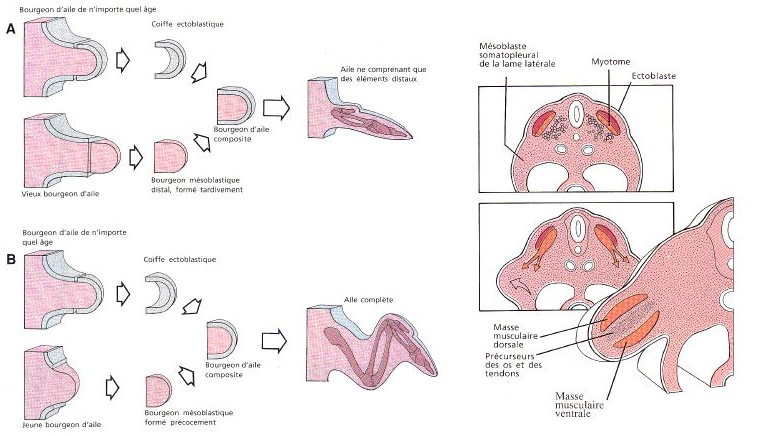
Quand la longueur du bourgeon excède sa largeur, la différenciation interne du membre commence. Le mésenchyme central, originaire des sclérotomes, se différencie en squelette, d'abord cartilagineux puis osseux, selon un gradient proximo-distal. Le mésenchyme qui l'entoure, provenant des myotomes, s'allonge et se différencie en muscles striés, selon le même gradient.
Les cellules du mésoderme latéral fournissent probablement la composante conjonctive. Les vaisseaux sanguins se forment .
L'innervation motrice des muscles provient des neurones moteurs somatiques de la corne ventrale (plaque basale) de la moelle épinière; elle est très précoce.
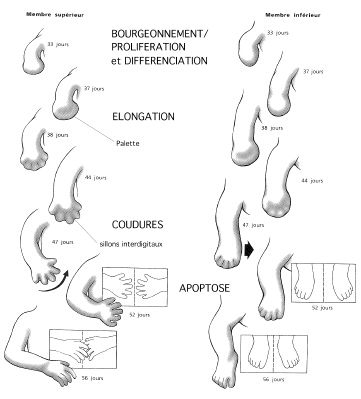
Chez les Tétrapodes ( membre chiridien) ,le bourgeon du membre s'aplatit, surtout distalement. La région distale aplatie s'élargit et, de circulaire, devient pentagonale, les pointes représentant les bourgeons des doigts. Les bourgeons des doigts continuent de croître et le tissu entre eux se nécrose, menant à la formation de doigts bien individualisés, sauf chez les animaux à pattes palmées chez qui la nécrose est moins accentuée.
Une flexure se crée à l'origine du membre l'articulant avec le tronc: l'épaule pour le membre antérieur, la hanche pour le membre postérieur . Vers le milieu du membre, une autre flexure se dessine et détermine la position du joint du coude ou du genou qui sépare le bras ou la cuisse de l'avant-bras ou la jambe . Une flexure plus distale marque le joint du poignet ou de la cheville qui sépare l'avant-bras de la main ou la jambe du pied (. De plus, les mains et les doigts de même que les pieds et les orteils s'articulent en plusieurs segments par des jointures additionnelles.
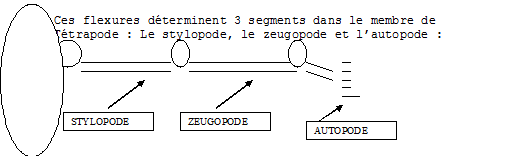
a Le stylopode.
Il est formé d’un seul os : l’humérus pour le membre antérieur et le fémur pour le postérieur. Cet os s’articule grâce au coude ou au genou.
b Le zeugopode.
Il est formé de deux os : le radius et le cubitus pour le membre antérieur, le tibia et le péroné pour le membre postérieur. Ces os s’articulent avec l’autopode grâce au poignet ou à la cheville.
c L’autopode.
C’est la main ou le pied.
Sa structure est pluriarticulée et très complexe.
- Le basipode (poignet ou cheville) : il est formé de trois rangées d’os nommés en fonction des os du zeugopode. Pour le membre antérieur, on a : le radial, l’intermédiaire et le cubital. Pour le membre postérieur, on a : le péronéal (calcanéum), l’intermédiaire et le tibial (os tragal).
On a ensuite les os centraux (de 0 à 5 os).
Enfin, on a 5 carpiens (pour la main) ou 5 tarsiens (pour le pied).
- Le métapode : Il est représenté par la paume de la main ou par la plante du pied. Ce métapode est composé de 5 métacarpiens ou métatarsiens.
- L’acropode : Il est représenté par les doigts ou par les orteils. Chaque partie est composée d’une succession d’os (les phalanges) articulés entre eux. Les phalanges étaient nombreuses au départ mais ont maintenant une tendance à la réduction. Chez les mammifères,le doigt 1 (le pouce) a deux phalanges, les doigts 2 à 5 ont tous trois phalanges. De l’intérieur vers l’extérieur, on trouve la phalange, la phalangine et la phalangette. Dans le cas du pouce, c’est la phalangine qui disparaît.
Remarque : Le nombre de cinq doigts est un phénomène secondaire. Au début, chez certains amphibiens on trouvait neuf doigts.
La main humaine est assez primitive, ce qui lui laisse une grande possibilité évolutive.
Originellement, les muscles fléchisseurs des membres se retrouvent ventralement et les muscles extenseurs dorsalement. Au fur et à mesure de la croissance, il se produit, chez plusieurs tétrapodes, une rotation de l'épaule et de la hanche qui résulte en des positionnements postérieur des fléchisseurs et antérieur des extenseurs.
Notons que les nageoires pectorales des poissons sont les précurseurs évolutifs des membres des tétrapodes et que leur formation implique un apport du feuillet somatique du mésoderme latéral. Nous n'entrerons pas dans ces détails qui seront exposés en anatomie et morphologie des Vertébrés où nous étudierons également le cas des Vertébrés non Tétrapodes.
4. L’entoblaste
4.1 Destinée du feuillet entoblastique
L’entoblaste intervient dans l’édification :
- Du tube digestif et des glandes annexes ( épithélium )
- De l’appareil respiratoire, branchial et pulmonaire ( épithélium )
- Des dérivés pharyngiens : thyroïde, parathyroïde, thymus,tympan, trompes d’eustache…………
4.2 Tube digestif et glandes annexes
4.2.1 Origine
Le tube digestif se forme de façon différente selon que l'embryon dérive d'un oeuf pauvre ou riche en vitellus, donc à segmentation holoblastique ou méroblastique, et selon le type de gastrulation.
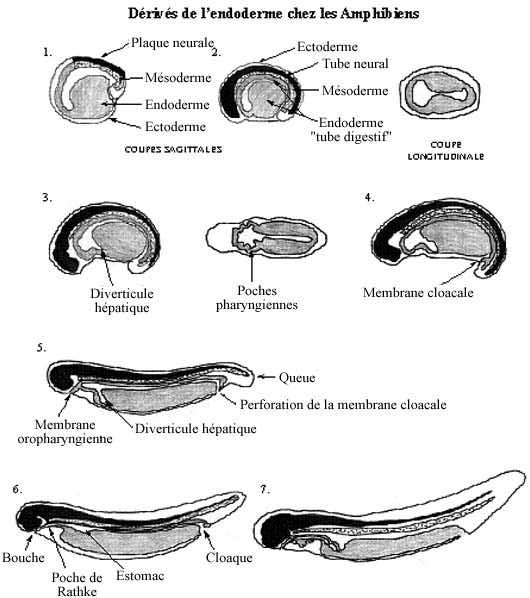
Reportons-nous à la fin de la gastrulation. Chez plusieurs poissons et chez les amphibiens, les trois feuillets fondamentaux: endoderme, mésoderme et ectoderme, sont emboîtés, comme des poupées russes; l'endoderme, feuillet le plus interne, renferme une cavité, l'archentéron.
Les trois feuillets concentriques ne sont pas sphériques, à cause de l'élongation antéro-postérieure de l'embryon. Ainsi, l'archentéron tapissé d'endoderme prend une forme tubulaire, présage du futur tube digestif. De plus, son diamètre n'est pas constant sur toute sa longueur. La portion la plus antérieure est relativement élargie et sa paroi endodermique est mince: c'est le tube digestif antérieur. La portion moyenne a un diamètre réduit; la partie dorsale de sa paroi (le toit) est mince tandis que l'épais plancher consiste en blastomères plus fortement vitellins: c'est le tube digestif moyen. La portion caudale est de diamètre réduit et se termine par le blastopore (anus présomptif): c'est le tube digestif postérieur.
En gros, la portion antérieure deviendra l'épithélium tapissant la bouche, le pharynx et la région branchiale, l'oesophage, l'estomac, le duodénum de l'intestin grêle, le foie et le pancréas. La portion moyenne deviendra l'épithélium tapissant le jéjunum et l'iléum de l'intestin grêle et le gros intestin (colons, appendice, rectum). La portion postérieure deviendra le cloaque ainsi que la vessie urinaire, formée à partir du cloaque.
Au site du blastopore, l'endoderme et l'ectoderme ne sont pas séparés l'un de l'autre par du mésoderme. Les deux feuillets fusionnent, formant la membrane cloacale qui se perfore pour devenir l'anus.
À mesure que s'allonge l'embryon et que se caractérisent les différentes régions du tube digestif, certaines d'entre elles s'allongent encore plus que l'embryon entier et deviennent circonvoluées. Ceci est particulièrement marqué dans le cas du tube digestif moyen.
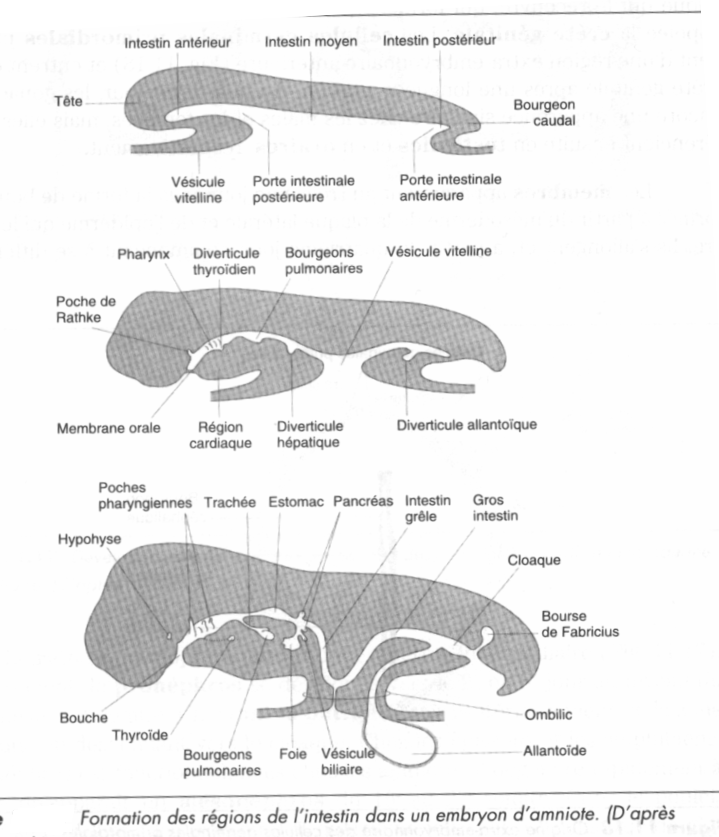
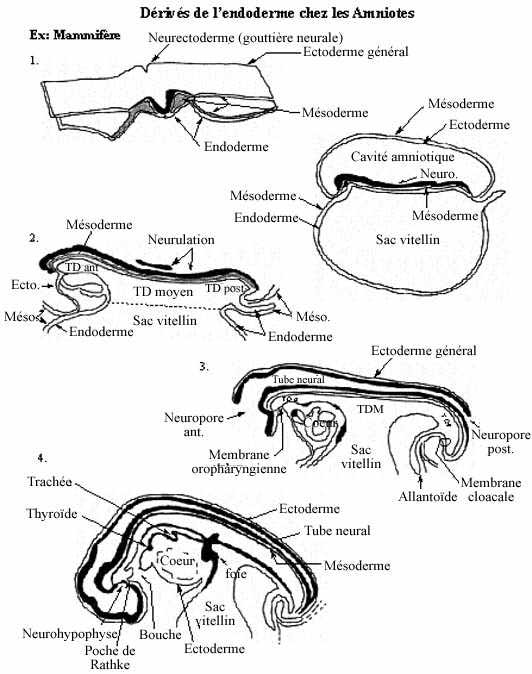
Chez les reptiles et les oiseaux, les 3 feuillets fondamentaux ne sont pas emboîtés à la fin de la gastrulation mais empilés sur le sac vitellin, nonobstant un certain espace entre ce dernier et l'endoderme, espace ne pouvant être considéré comme un archentéron véritable, mais auquel on donne néanmoins le nom, par commodité. Chez les mammifères, l'organisation des feuillets est la même mais le "sac vitellin" ne renferme pas de vitellus. Il a été nommé ainsi par analogie avec le sac vitellin des reptiles et oiseaux.
Au cours de la gastrulation, les cellules migrent à travers le blastopore ( Amphibiens ) ou la ligne médiane ( oiseaux, Mammifères ), chez les Oiseaux, par exemple, elles remplacent l’hypoblaste pour former un endoderme ( entoblaste ) définitif. Le tube de l’intestin lui-même est formé par le repliement du corps en s’éloignant du blastoderme.
La partie antérieure du corps est soulevée du blastoderme et l’endoderme devient un tube définitif de l’intestin antérieur.
La jonction de l’intestin antérieur et de l’intestin moyen ou porte intestinale antérieure est le niveau où le tube dans le feuillet plat d’endoderme.
Un peu plus tard, un processus similaire se déroule à l’extrémité arrière et l’intestin postérieur est formé, la jonction entre l’intestin moyen et l’intestin postérieur étant la porte intestinale postérieure.
La bouche se forme par fusion de l’intestin antérieur ventral avec l’épiderme qui le recouvre , un cloaque se forme à l’extrémité postérieure.
La croissance et les phénomènes morphogénétiques affectent, à ce stade,surtout les extrémités de l’embryon, de telle manière que la zone résiduelle d’endoderme, ouverte, se rétrécit rapidement. Elle se réduit au canal vitellin connectant l’intestin moyen à la masse vitelline .
A mesure que l’intestin se ferme , l’endoderme et le mésoderme se referment autour de lui pour former la paroi ventrale du corps. Celle-ci est éventuellement complète , sauf pour l’ombilic qui est la connection aux tissus extra – embryonnaires renfermant, le canal vitellin, les vaisseaux vitellins et l’allantoïde.
4.2.2 Le tube digestif et les glandes annexes.
4.2.2.1 Anatomie chez les Vertébrés.
4.2.2.1.1 Le tube digestif se divise en 5 segments différenciés sur le plan morphologique , histologique et fonctionnellement spécialisés :
- Bouche
- Pharynx
- Œsophage
- Estomac
- Intestin
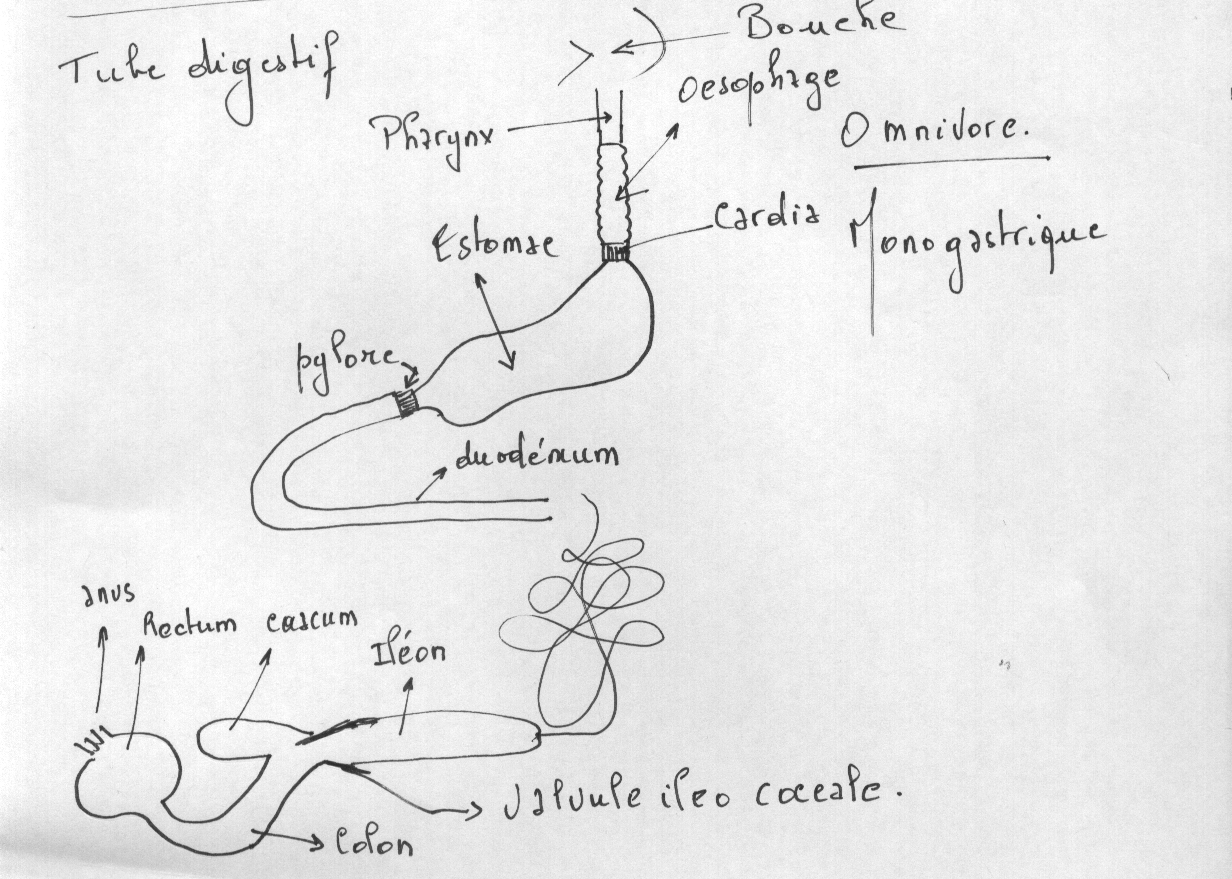
4.2.2.1.1.1 L'œsophage
L'œsophage est la portion du tube digestif qui poursuit caudalement le pharynx. Elle se développe au sein du médiastin embryonnaire et s'allonge au cours de la croissance de l'embryon. A l'origine, la lumière œsophagienne est obstruée par un bouchon épithélial qui disparaît par la suite.
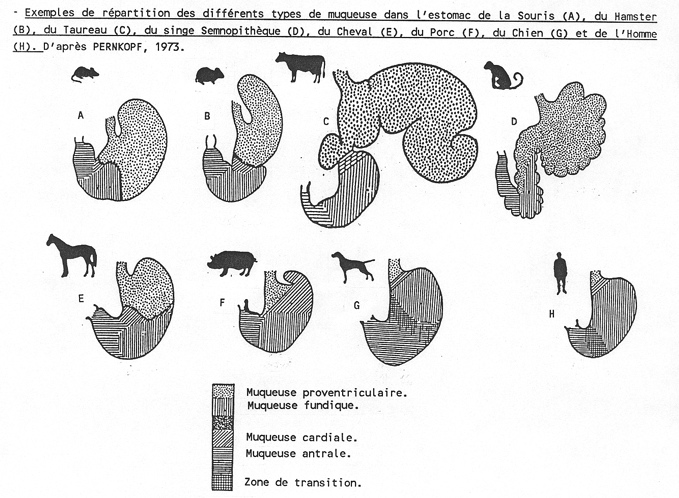
4.2.2.1.1.2 L'estomac simple
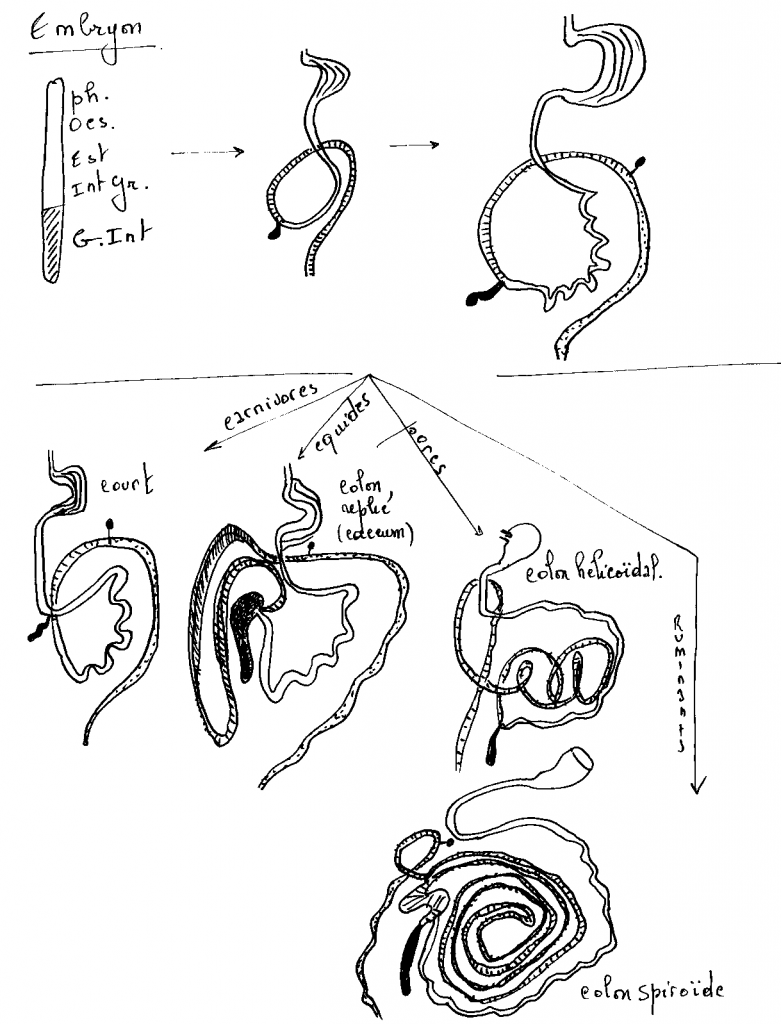
Les premières phases du développement de l'estomac sont similaires chez les polygastriques et chez les monogastriques. Le primordium de l'estomac se présente comme une dilatation de l'archentéron crânial, à convexité dorsale, en avant du septum transversum. Cette dilatation est le fait d'une croissance différentielle des bords dorsal et ventral. Deux courbures apparaissent, la grande courbure dorsale (futur fundus) et la petite courbure ventrale. Chez les monogastriques ou animaux à estomac simple (carnivores, équidés, suidés, primates, rongeurs,...), deux rotations vont amener cet organe à sa position définitive. La première rotation est précoce et se produit autour de l'axe crânio-caudal (ou longitudinal). Elle bascule la grande courbure à gauche, en sens inverse des aiguilles d'une montre pour l'observateur placé derrière l'embryon, d'un angle de 90 degrés. La grande courbure se retrouve à peu près horizontale.
La seconde rotation se fait autour de l'axe dorso-ventral, dans le sens inverse des aiguilles d'une montre pour l'observateur placé au-dessus de l'embryon, d'un angle de 90 degrés. L'extrémité caudale de l'estomac (futur pylore) pivote crânialement et à droite de la ligne médiale et l'extrémité crâniale de l'estomac (futur cardia) pivote caudalement et à gauche de la ligne médiale. Cette seconde rotation se réalise durant la vie fœtale et néonatale. Elle est en partie due à la croissance du foie qui pousse l'extrémité crâniale de l'estomac vers la gauche ( voir le chapitre relatif au coelome).
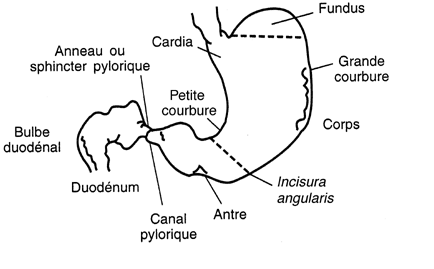
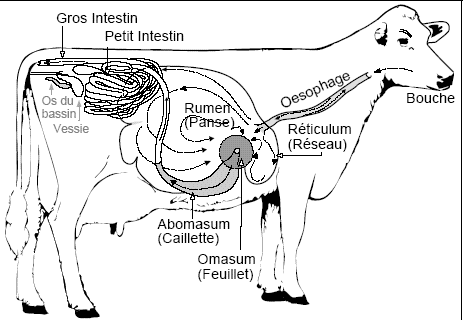
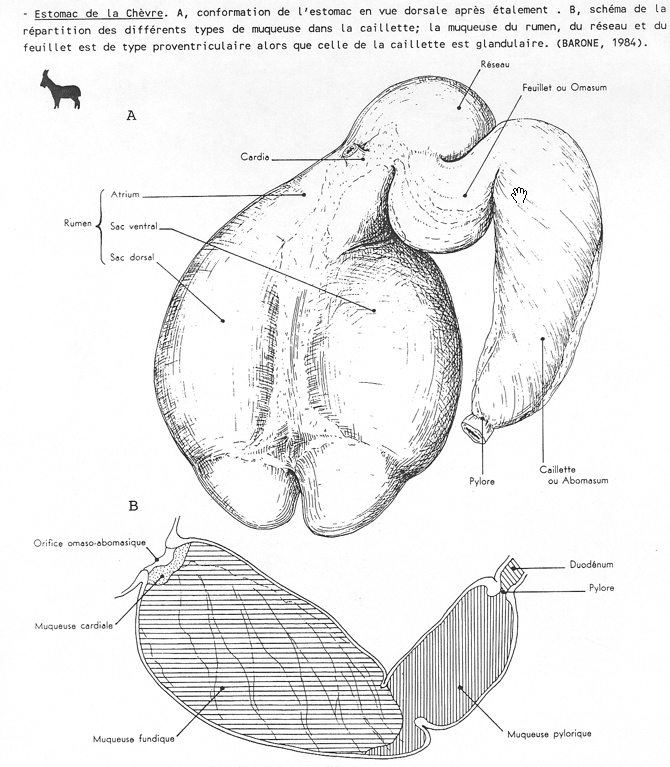
4.2.2.1.1.3 L'estomac des polygastriques
Le développement prénatal
Il débute comme chez les monogastriques par la dilatation dorsale et le basculement à gauche de la grande courbure. Au niveau de la grande courbure apparaissent trois autres zones de croissance différentielle. La plus crâniale donnera le rumen (la panse), la moyenne se transformera en réticulum (le réseau) et la plus caudale fournira l'abomasum (la caillette). Au niveau de la petite courbure, une zone de croissance différentielle apparaît à hauteur du futur réticulum. C'est l'ébauche de l'omasum (le feuillet).
Le rumen se développe essentiellement à gauche de la ligne médiale et se compartimentalise en deux sacs superposés, les sacs dorsal et ventral. Il repousse le réticulum crânialement et ventralement, l'omasum et l'abomasum ventralement et à droite.
La croissance du rumen se ralentit ensuite et, à la naissance, sa taille représente environ la moitié de celle de l'abomasum.
Le développement postnatal
Après la naissance, le rumen reprend sa croissance pour atteindre la même taille que l'abomasum vers 8 semaines chez le veau. A 12 semaines, le rumen est deux fois plus grand que l'abomasum et vers 18 mois, les différents compartiments stomacaux atteignent leur taille adulte. Le volume du rumen représente alors 80% de l'ensemble. Sa croissance est favorisée par une alimentation grossière qui stimule son activité de cuve de fermentation
Ces différentes parties du tube digestif seront étudiées plus scrupuleusement en anatomie des Vertébrés.
Remarque : tout en étant des polygastriques, l’anatomie des estomac de ruminants ne sont pas semblables en tous points à ceux des Camélidés.
4.2.2.1.1.4 Le développement de l'intestin
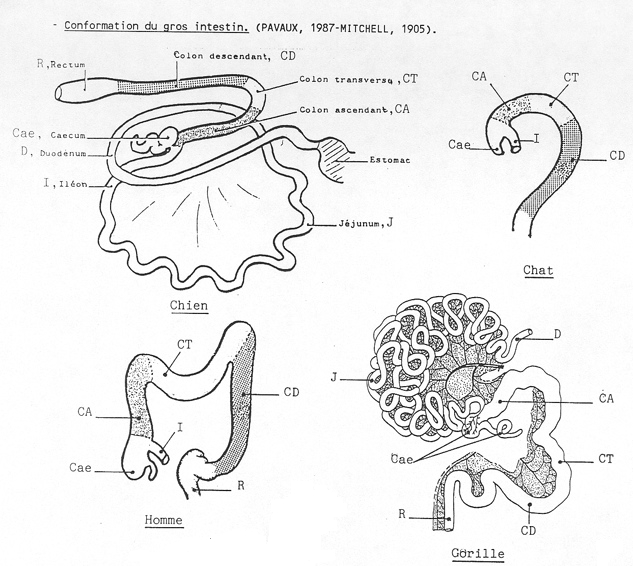
Les différents segments du tractus intestinal se forment à partir de l'anse intestinale primitive. Celle-ci est située derrière l'estomac et présente une convexité ventrale. On distingue tout d'abords une courte partie rectiligne ou futur duodénum, une partie descendante ou futur jéjunum, un coude au niveau duquel vient s'aboucher le canal vitellin, une première partie ascendante ou futur iléon, un renflement cæcal ou futur cæcum, une seconde partie ascendante ou futurs côlons ascendant et transverse et enfin une partie terminale ou futur côlon descendant. Le cloaque qui ferme caudalement le tractus digestif donnera le rectum.
L'anse intestinale primitive est initialement placée dans le plan médian. Elle est suspendue par le mésentère dorsal. Au sein de ce mésentère, l'artère mésentérique crâniale se forme entre la partie descendante et la partie ascendante de l'anse. Cette artère agit comme un pivot de rotation. L'anse opère une rotation de 180 degrés autour de l'axe dorso-ventral dans le sens des aiguilles d'une montre pour l'observateur placé au-dessus de l'embryon. La partie ascendante passe d'une position caudale vers une position crâniale par la gauche. La partie descendante crâniale passe en position caudale par la droite.
Après cette première rotation, une seconde intervient dans le même sens. Elle est d'environ 150 degrés et porte la partie descendante vers l'avant et par la gauche et la partie ascendante vers l'arrière et par la droite. Le duodénum et le côlon transverse se croisent autour de l'axe vasculaire.
Suivant l'espèce, les différents segments du tractus intestinal se développeront plus ou moins. Disons dès à présent que ce type de base présente des variations adaptées au régime alimentaire.
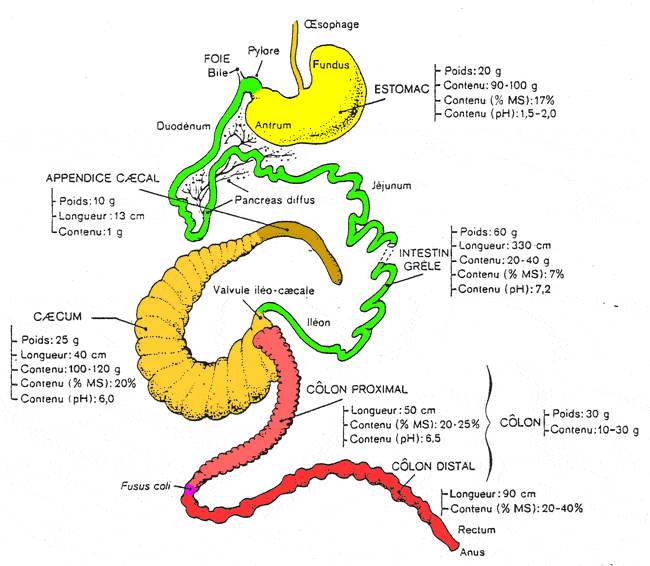
Le tractus intestinal des carnivores
Chez les carnivores (chien et chat), le duodénum (0,3 m), le jéjuno-iléon (3,5 m) sont relativement courts. Le cæcum est rudimentaire et le côlon (0,7 m) est divisé en trois brèves portions : le côlon ascendant, le côlon transverse et le côlon descendant. Cette disposition simple est également observée chez les primates.
Le tractus intestinal des équidés
Chez les équidés, les deux particularités de l'intestin sont le développement considérable du cæcum dans le flanc droit et celui du côlon ascendant (8 m) qui se replie en 4 portions successives logées ventralement et à gauche du caecum. Le jéjuno-iléon est proportionnellement plus développé que chez les carnivores (22 m).
Le tractus intestinal des ruminants
Les ruminants se caractérisent par un développement considérable du jéjuno-iléon (40 m), un développement moyen du caecum et un côlon ascendant long (11 m) et disposé en spirale.
Le tractus intestinal des suidés
Il ressemble à celui des ruminants. Le jéjuno-iléon est assez développé (15 m), le caecum l'est moyennement et le côlon ascendant (3-5 m) est disposé en spirale.
Le côlon transverse, le côlon descendant, le rectum et l'anus
Le côlon transverse fait suite au côlon ascendant chez toutes les espèces. Il est court et passe devant le duodénum, de droite à gauche pour repartir caudalement où il se poursuit par le côlon descendant.
Le côlon descendant conserve sa structure tubulaire. Il est suspendu par un méso dorsal et est déplacé à gauche de la ligne médiane par les autres viscères. Il débouche dans un élargissement final, le rectum.
mésodermique cardiaque, effet inhibiteur de la corde...)- voir embryologie expérimentale et moléculaire.
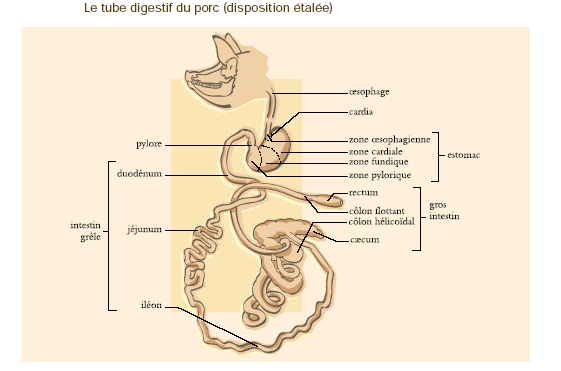
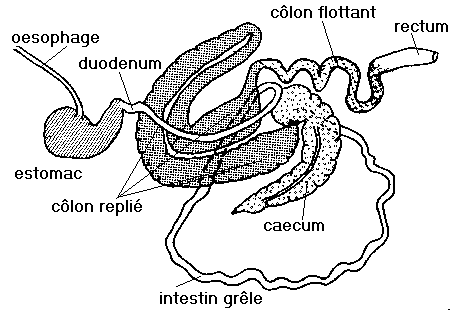
Tube digestif du cheval
Tube digestif de Lapin

4.2.2.1.2 Les glandes annexes
Les glandes annexes sont :
- Les glandes salivaires.
- Le foie.
- Le pancréas.
4.2.2.2 Origine embryologique
Les deux phénomènes essentiels de la digestion, à savoir la sécrétion d’enzymes et l’absorption se réalisent au niveau de l’épithélium d’origine entoblastique ( voir point 4.2.2 )
Cet épithélium est cependant absent :
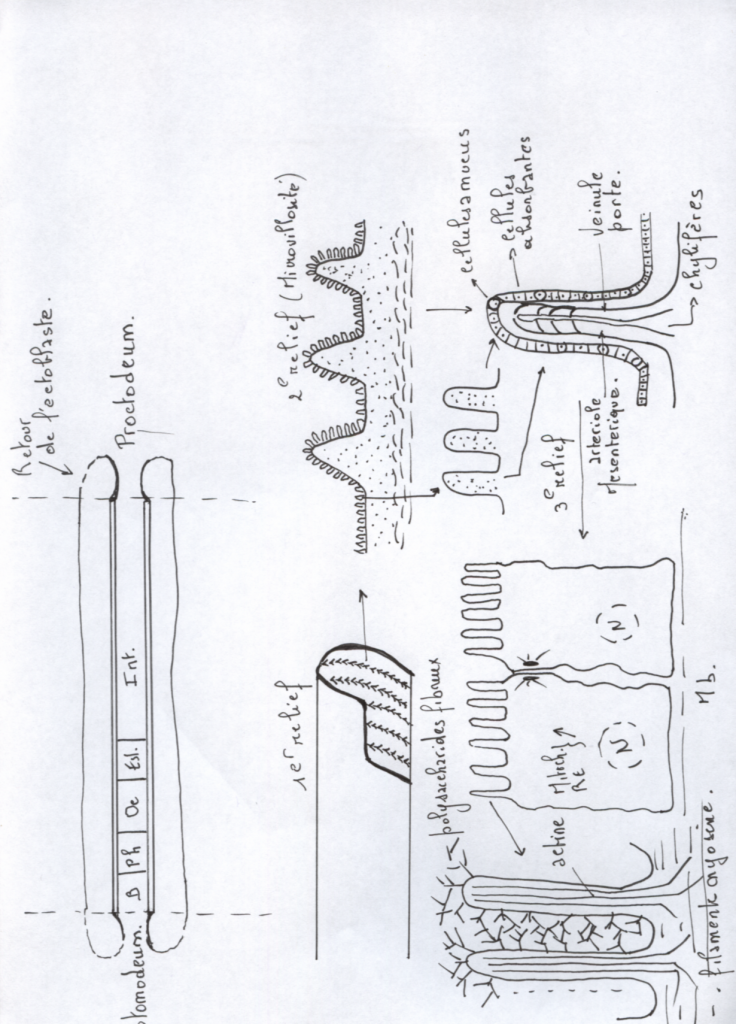
- De la bouche, la cavité buccale ou stomodéum est recouverte par l’épiblaste.
- Le cloaque ou proctodeum est également recouvert par l’épiblaste.
Notons que la différenciation antéro-postérieure du tube digestif ne peut se réaliser que si des signaux d’inductions sont envoyés par le mésoderme ( influence de la zone cardiaque et de la corde )
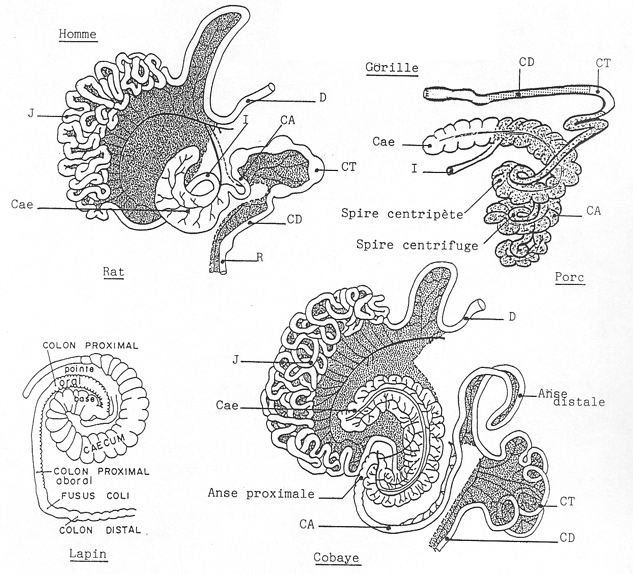
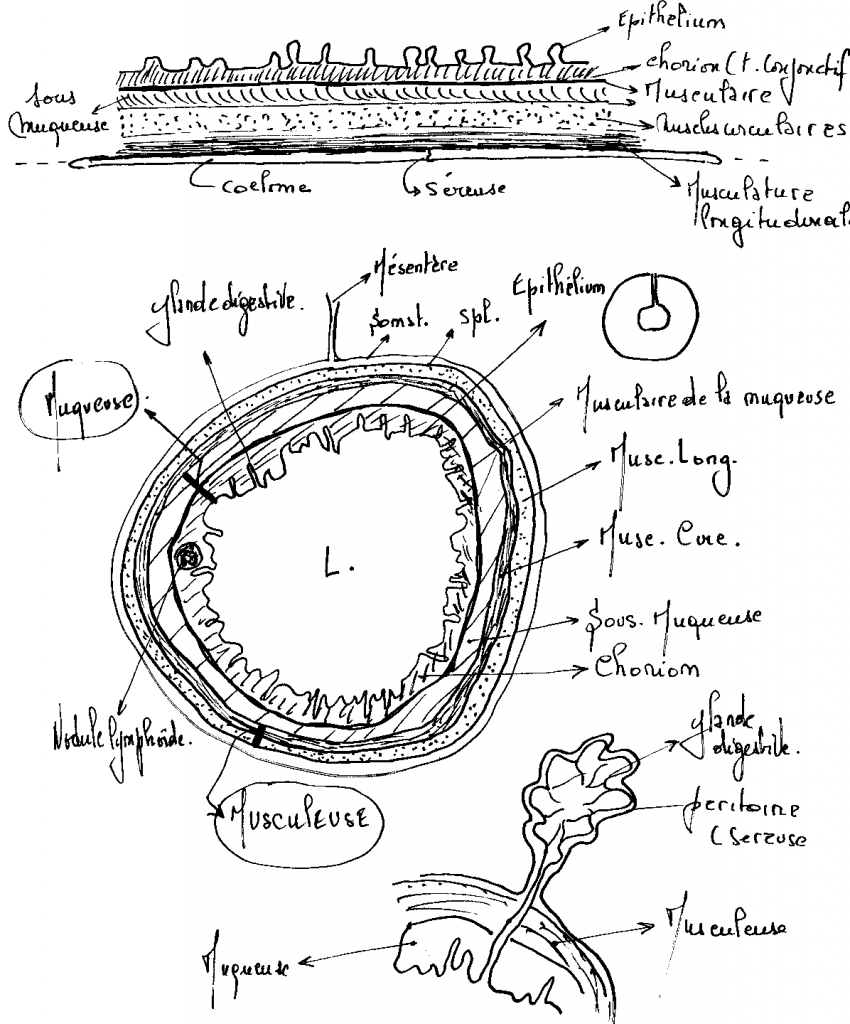
4.2.2.3 Histologie
4.2.2.3.1 L’épithélium de l’intestin est caractérisé par la différenciation de deux grandes catégories de cellules
- Les cellules muqueuses et séreuses :
Les muqueuses disposées sur toute la longueur du tube sécrètent un mucus ayant pour fonctions le transport des aliments et la protection de la muqueuse.
Les séreuses sécrètent des enzymes responsables de la digestion , elles sont localisées dans l’épithélium gastrique et une partie de l’intestin. Ces cellules glandulaires sont situées :
- Soit au niveau d’invaginations épithéliales
- Soit différenciées à l’extérieur de la paroi du tube en volumineuses formations autonomes reliées à la cavité par des canaux excréteurs ( glandes salivaires, foie, pancréas.
Epithélium digestif
4.2.2.1.2 Les grandes glandes
4.2.2.1.2.1 Les glandes salivaires
La salive est une sécrétion aqueuse contenant du mucus, des ions minéraux et des enzymes. Elle a un rôle digestif qui est l'humidification et la lubrification des aliments, l'hydrolyse partielle de l'amidon par l'amylase (première enzyme digestive rencontrée par les aliments); elle a aussi un rôle antiseptique grâce au lysozyme, enzyme antibactérienne, notamment vis à vis des caries dentaires.
La salive est produite par trois paires de glandes salivaires principales : les glandes parotides, les glandes sous-maxillaires, les glandes sub-linguales, et par de nombreuses petites glandes salivaires accessoires éparpillées dans la paroi de la cavité buccale : lèvres, langue, palais, joues.
Structure générale
Les petites glandes salivaires sont contenues dans le chorion de la muqueuse buccale.
Les glandes salivaires principales sont des organes cernés par une capsule conjonctive fibreuse; de cette capsule partent des cloisons conjonctives qui délimitent des lobules d'où l'aspect anatomique bosselé de ces glandes; l'intérieur des lobules forme le parenchyme salivaire; les cloisons interlobulaires contiennent des canaux excréteurs, des vaisseaux et des nerfs.
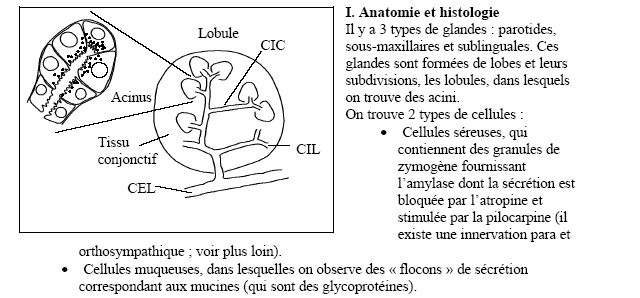
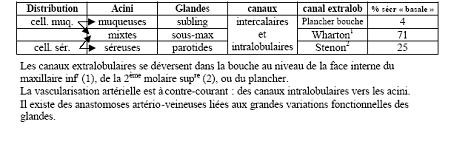
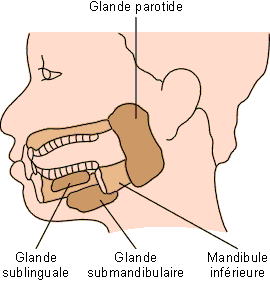
Figure : Localisation des glandes salivaires chez l'homme. La glande parotide est située devant l'oreille, entre le muscle masséter et la peau; son conduit s'ouvre au niveau des molaires supérieures. La glande submandibulaire occupe une position centrale sous le plancher buccal; son conduit débouche derrière les incisives inférieures à la base du frein. La glande sublinguale est localisée sous la langue, contre la face interne de la mandibule inférieure; elle s'ouvre par de nombreux conduits au niveau du plancher buccal.
Glandes parotides
Les deux glandes parotides droite et gauche sont les plus volumineuses des glandes salivaires. Elles épousent les parois de la loge parotidienne, ce qui leur confère une forme en prisme à large base latérale. Ce sont des glandes tubulo-acineuses. La salive est élaborée à partir de cellules séreuses groupées en acini. Elle est ensuite drainée par un système canalaire arborisé, tapissé par des cellules épithéliales, convergeant vers un canal collecteur principal : le canal parotidien ou canal de sténon ( aboutissant au niveau de la seconde molaire supérieure ).
Les acini parotidiens contiennent également des cellules myoépithéliales, éléments contractiles qui permettent un drainage actif de la salive.
La parotide contient en outre un tissu conjonctif de soutien constitué de fibres collagènes, de fibroblastes, de vaisseaux, de lymphatiques et de nerfs. On retrouve également des plasmocytes synthétisant une immunoglobuline de type A sécrétoire jouant un rôle dans les mécanismes immunologiques de défense.
Glandes sous maxillaires
Les glandes sous-maxillaires (situées sous le plancher buccal), leur conduit débouche au niveau des incisives inférieures.
Glandes sublinguales
Les glandes sublinguales (situées sous la langue), de nombreux conduits débouchent au niveau du plancher buccal.
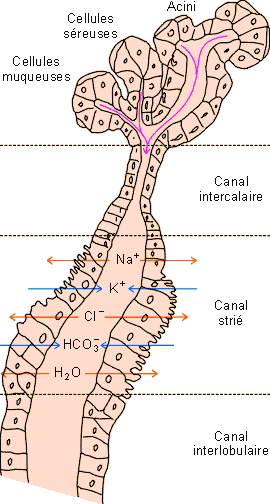
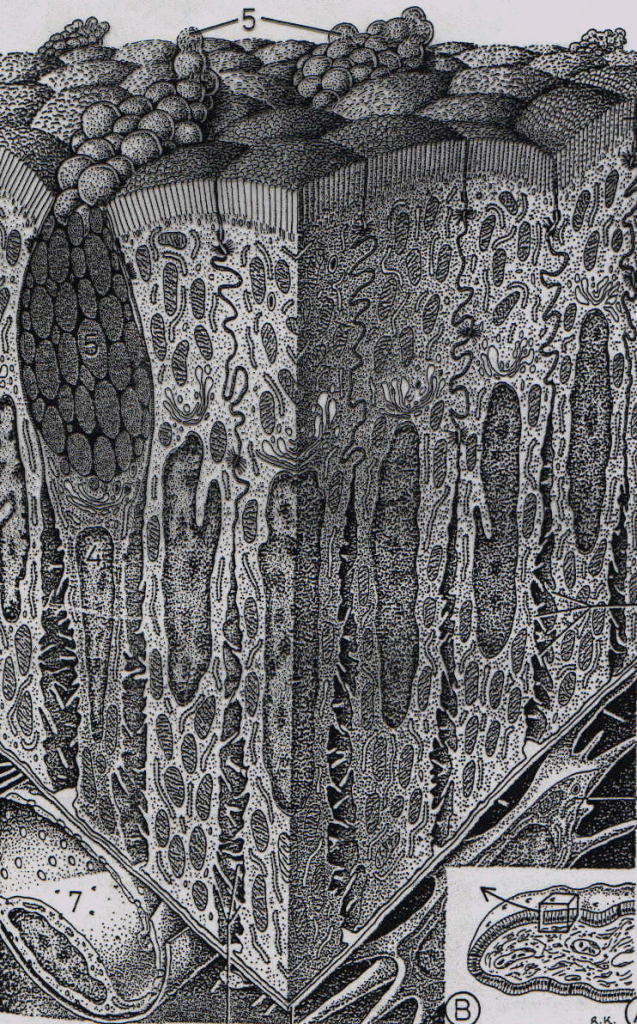
Figure : Structure d'une glande salivaire de mammifère. Les acini sont essentiellement constitués de cellules dites séreuses produisant les diverses protéines de la salive. Les mucines sont sécrétées par des cellules dites muqueuses organisées en tubules muqueux faisant suite aux acini. L'ensemble est relié par un canal intercalaire à un canal strié au niveau duquel s'opère l'essentiel des modifications de composition.
4.2.2.3.2.2 L’anneau hépato pancréatique
Développement
Le foie et le pancréas proviennent d’une série de quatre bourgeons épithéliaux de l’intestin antérieur, formant l’anneau hépato-pancréatique.
Le bourgeon ventral est hépatique
Les bourgeons latéraux ( ventral droit et gauche ) et dorsaux sont pancréatiques,chez un nombre important de Vertébrés, seule l’ébauche dorsale se maintiendra.
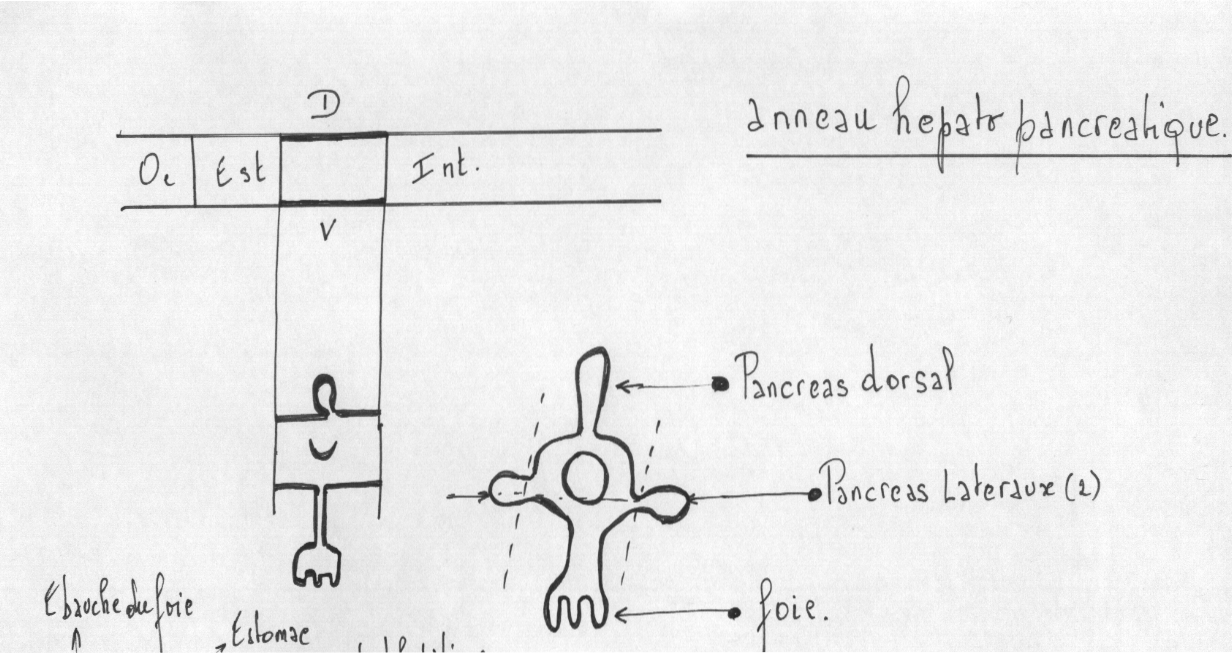
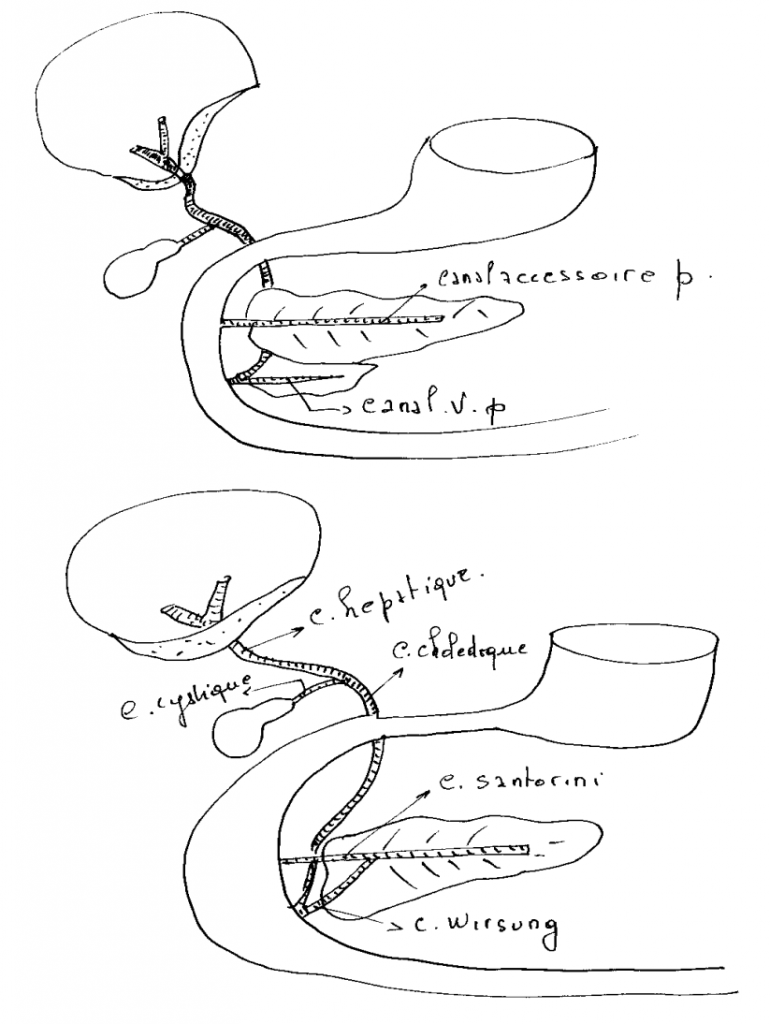
Une excroissance du plancher du duodénum, le diverticule hépatique, en s'embranchant maintes fois conduit à la formation de l'épithélium glandulaire du foie. À partir de cet épithélium, une autre excroissance produit l'épithélium (non glandulaire) tapissant la vésicule biliaire. La vésicule biliaire sert de réservoir de l'une des sécrétions hépatiques, la bile, et demeure reliée au foie par le canal biliaire. Le foie garde sa communication avec le duodénum, dans lequel il rejette ses sécrétions, par le canal pancréatique.
Une excroissance du toit du duodénum mène à la formation du pancréas dorsal tandis qu'une évagination du plancher du duodénum ou du canal pancréatique donne naissance au pancréas ventral. Sauf chez certains poissons, les deux rudiments pancréatiques se rejoignent éventuellement en un organe unique qui joint le canal pancréatique. Les sécrétions exocrines du pancréas sont donc rejetées dans la lumière duodénale.
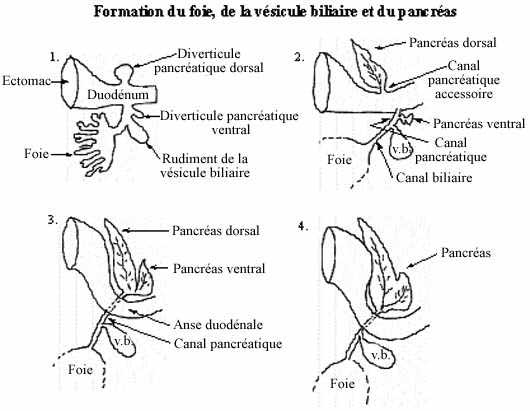
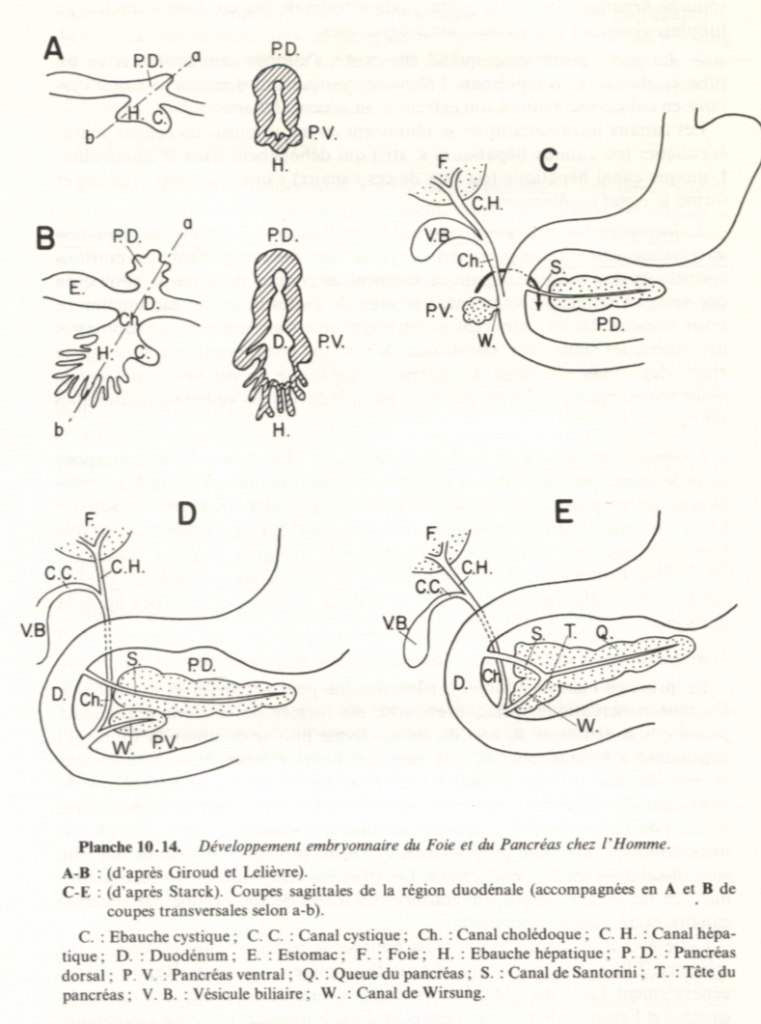
LE FOIE
Fondamentalement, c’est une glande à sécrétion externe, mais elle est également adaptée à la sécrétion interne
Ces deux types de sécrétion sont assurées par les mêmes cellules.
- Sécrétion interne : Pas d’hormones, mais un ensemble de substances métaboliques ( glucose, lipoprotéines, protéines plasmatiques ). Pratiquement toutes les réactions du métabolisme intermédiaire ( synthèses, dégradations ) peuvent se réaliser dans cette glande.
- Sécrétion externe : bile contenant les produits de sécrétion ( les sels biliaires ) et des produits d’excrétion ( pigments biliaires.).
Le foie occupe une position stratégique dans l’organisme.
Structure du foie.
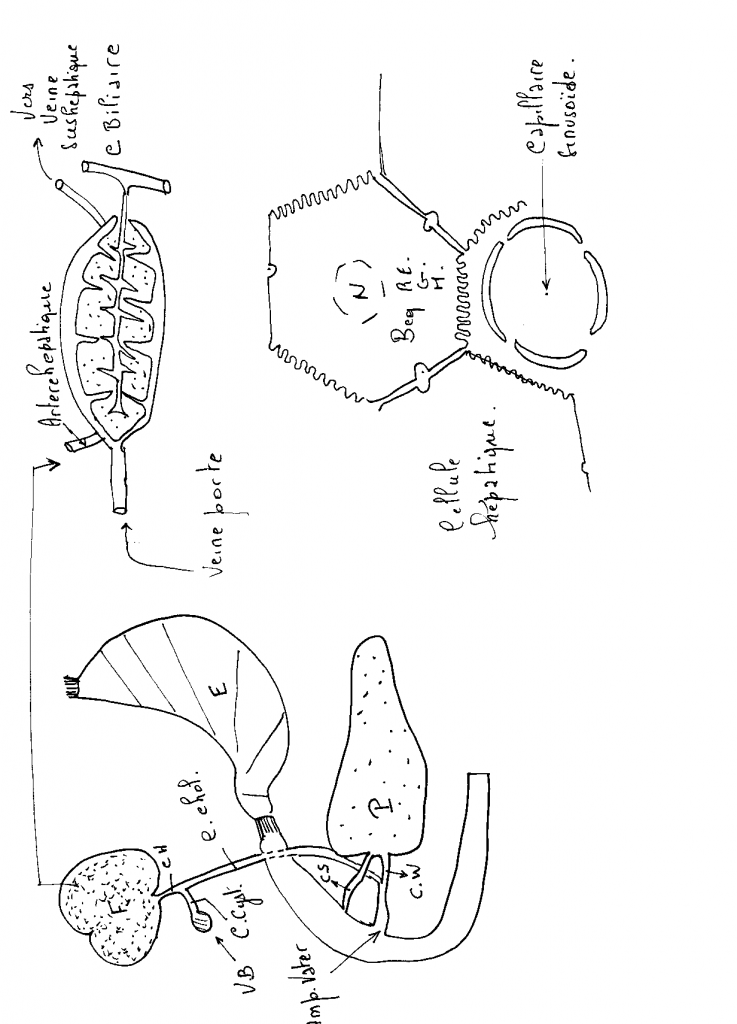
L'unité de base est la cellule hépatique ou hépatocyte, qui forme le parenchyme hépatique. Elle est riche en substance de réserve: le glycogène.
Les cellules vont se regrouper pour former un lobule.
Les lobules sont séparés par du tissu conjonctif où cheminent des vaisseaux et des canaux biliaires intra-hépatiques.
Il faut distinguer l’acinus hépatique et le lobule hépatique.
L’unité morphologique élémentaire est l’acinus de Rappaport qui est construit autour de la triade portale (artère, veine porte, canal biliaire) et forme autour de cette dernière une petite masse de forme analogue à une petite baie. On y distingue classiquement 3 zones (1,2,3) dont l’oxygénation est différente. Le sang de la branche veineuse se distribue dans l’acinus. Le sang artériel (à faible débit) arrive sous forte pression et se mêle au sang veineux au niveau des sinusoïdes hépatiques par un réseau de shunts, entraînant un effet de chasse qui favorise la circulation. Le sang est mieux oxygéné au centre de l’acinus (zone 1) qu’en périphérie zone 3), ce qui explique la répartition inégale de certaines lésions parenchymateuses. Le drainage de l’acinus se fait en périphérie par le réseau des veines collectrices qualifiées de veineuses centro-lobulaires
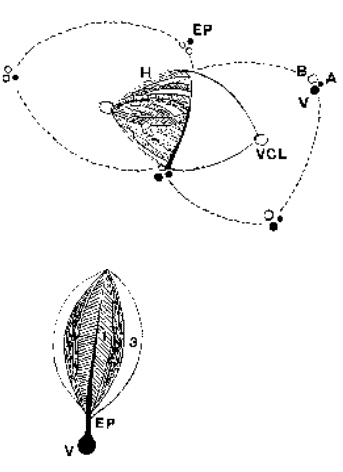
Le lobule hépatique (en pointillé) est axé sur la veine centro-lobulaire (VCL). Il est délimité en périphérie par les espaces portes (EP). Le lobule hépatique ne constitue pas une unité fonctionnelle mais doit être considéré comme une subvision anatomique du foie.
A l’intérieur de l’acinus, les hépatocytes (CH) se disposent en un réseau complexe de cloisons, perforés par les sinusoïdes et les canaux biliaires (CB) de drainage. En coupe ces cloisons se présentent sous forme de lames unistratifiées séparées les unes des autres par les sinusoïdes dont la structure est complexe. Leur endothélium (CE) est largement fenêtré, il ne repose sur aucune membrane basale. Il est séparé des hépatocytes par l’espace de Disse (ED) dans lequel on trouve des fibroblastes, des neurones et des cellules chargées de stocker de la vitamine A (cellules de ITO) (CI). Des cellules macrophagiques nombreuses (cellules de Kupffer) (CK) accolées à l’endothélium flottent dans la lumière du sinusoïde. Elles sont douées d’une importante capacité de phagocytose.
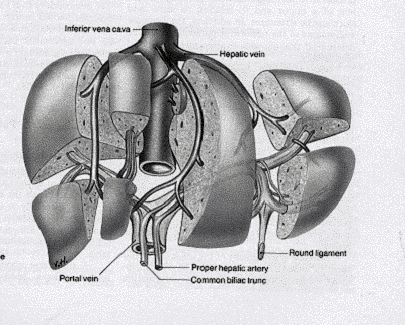
Vascularisation.
L'artère hépatique va assurer la circulation nutritionnelle de la glande hépatique, amener de l'oxygène indispensable à la vie des hépatocytes. Elle se ramifie au niveau du foie jusqu'au centre des lobules.
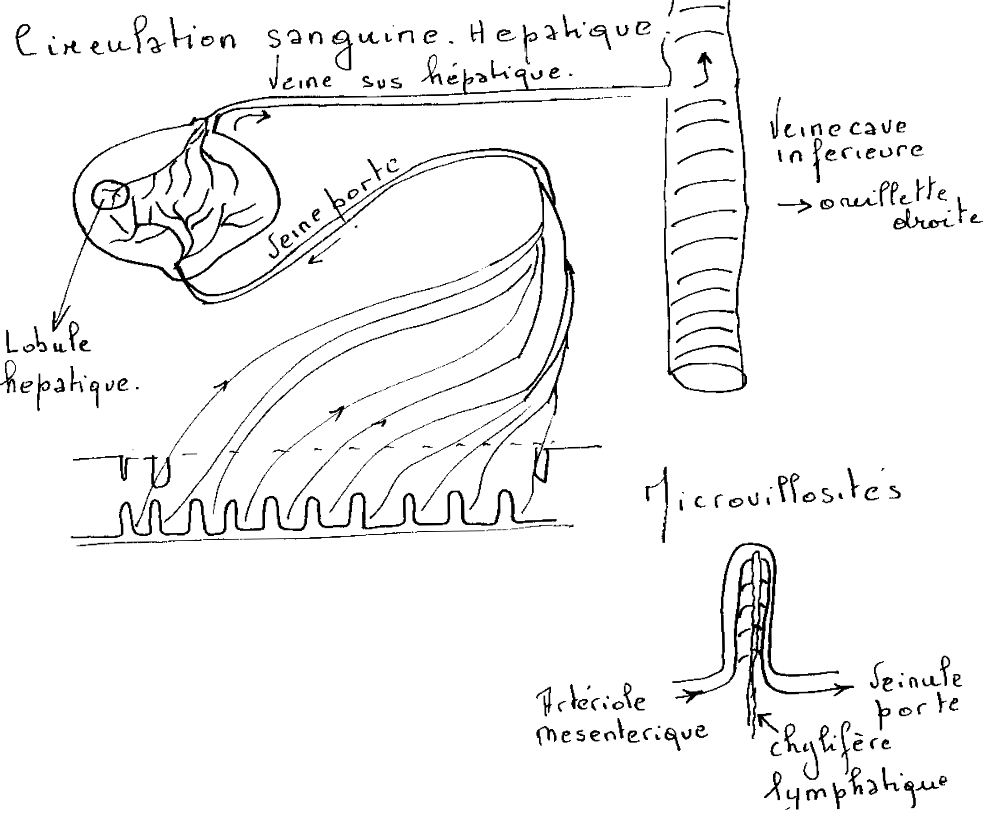
La veine porte et ses branches vont permettre d'assurer la circulation fonctionnelle du foie, car elle va apporter au foie des nutriments nécessaires aux hépatocytes qui va les stoker et les transformer. La veine porte se ramifie jusqu'au centre des lobules pour ensuite rejoindre la veine sus-hépatique qui va permettre le retour du sang.
Le système veineux sus-hépatique recueille la totalité du sang qui vient du foie et va ensuite se jeter dans la veine cave inférieure de l'oreillette droite.
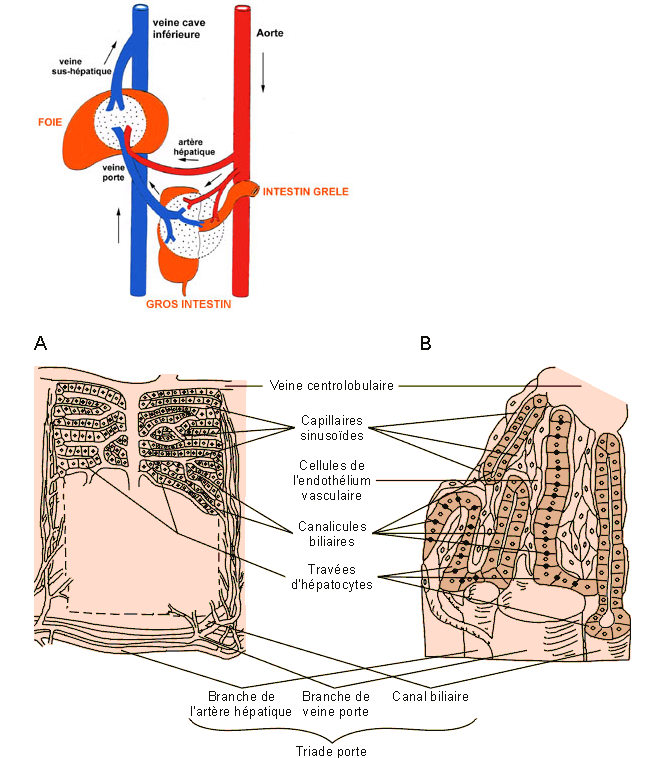
Figure : Organisation schématique du tissu hépatique d'un foie lobulé de mammifère. En coupe, le lobule hépatique à l'aspect d'un hexagone dont les angles sont entourés d'une triade porte amenant le sang oxygéné par des branches de l'artère hépatique et les produits du métabolisme et de la digestion par des branches de la veine porte. Ce système est en connexion avec les lumières des capillaires sinusoïdes qui entourent les travées d'hépatocytes (beiges en B). Les produits du métabolisme hépatique se déversent dans les sinusoïdes et sont drainés par la veine centrolobulaire. La sécrétion biliaire est déversée dans de fins canalicules entre les hépatocytes et drainée par les canalicules biliaires dans les canaux biliaires des triades porte.
Morphologie
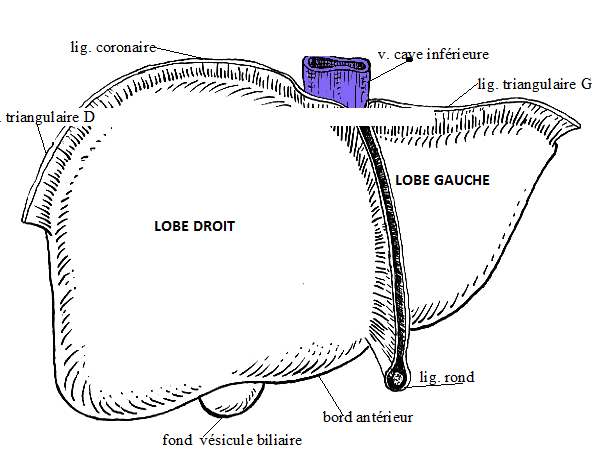
Le foie est un organe plein. Il présente deux faces (une crâniale qualifiée de diaphragmatique et une caudale de viscérale), séparées par quatre bords (dorsal, ventral et latéraux), le bord ventral étant échancré de fissures divisant l'organe en différents lobes.
La veine ombilicale (donc son vestige, le ligament rond), divise le foie en deux grands territoires, droit et gauche. La vésicule biliaire divise la partie droite en deux territoires, droit et intermédiaire, ce dernier étant lui-même séparé en deux étages par la porte du foie. La disposition primitive montre donc un lobe gauche, un lobe droit, un lobe carré (étage inférieur de la partie intermédiaire), un lobe caudé (étage supérieur) que l'échancrure de la veine cave caudale subdivise en processus caudé à droite et processus papillaire à gauche.
Les lobes droit et gauche peuvent eux-mêmes être divisés en lobes droits médial et latéral et en lobes gauches médial et latéral.
Histologie
Schématiquement, le foie est composé :
- D’une masse de cellules épithéliales formant le parenchyme hépatique
- De capillaires veineux appartenant au système porte
( voir schéma lobule hépatique )
La cellule hépatique ( hépatocyte )
-( voir schéma )
Cette cellule possède deux types de faces.
- Premier type :la face Comporte un espace péricapillaire ( espace de Disse) et est couverte de microvillosités.
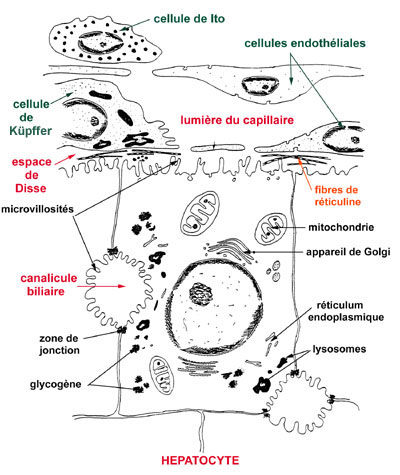
Deuxième type : en contacte avec les autres cellules hépatiques , formation de canalicules biliaires.
Remarques :
- Quantité importante de glycogène associée au REL
- Les capillaires sanguins sont du type sinusoïde, ils ont les caractéristiques suivantes :
- Calibre irrégulier, toujours grand
- Paroi endothéliale discontinue, existence de pores de 1 à 5 microns de diamètre. Passage facile du plasma dans l’espace péricapillaire et contact avec les microvillosités des entérocytes.
- Possibilité qu’on certaines cellules endothéliales, les cellules de Kupfer ( issues de la splanchnopleure ), de se transformer en cellules phagocytaires.
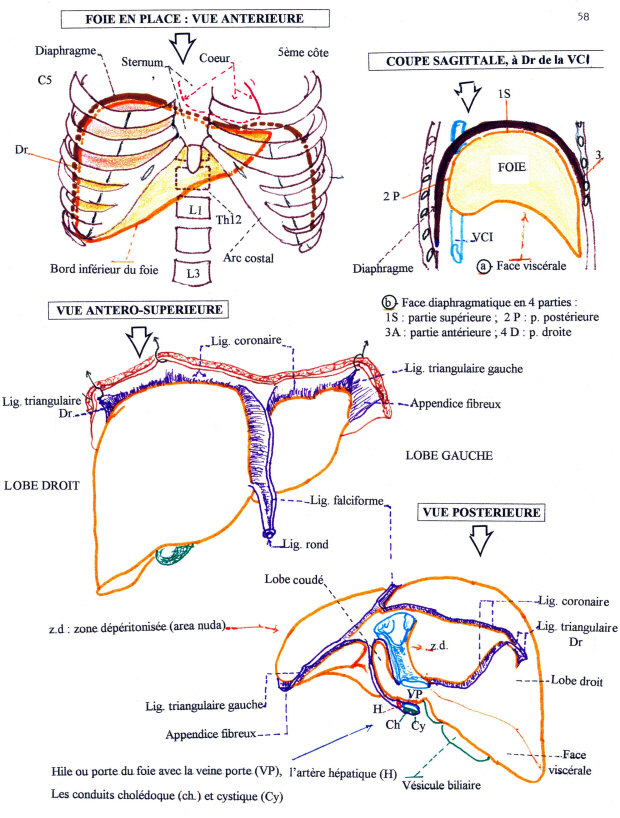
VOIES BILIAIRES ( voir schéma plus haut )
Une des fonctions du foie est la sécrétion de la bile. Elle est recueillie par les voies biliaires.
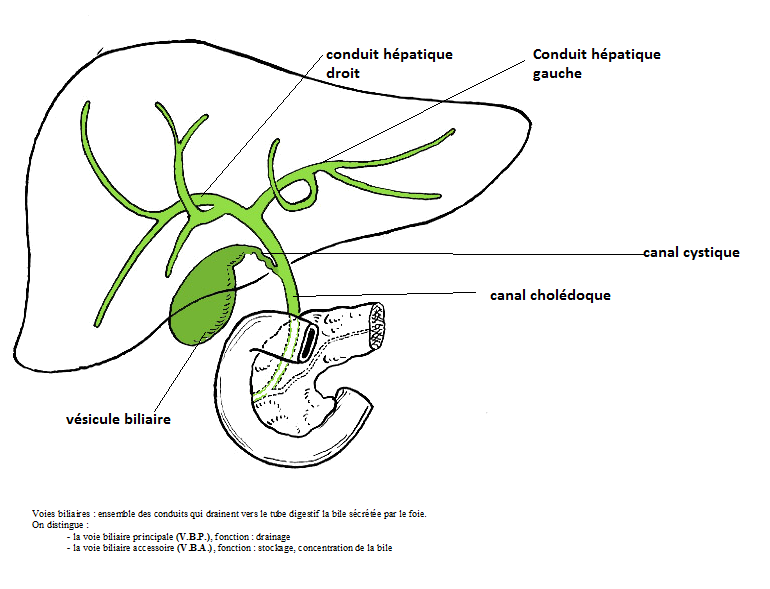
Les voies biliaires débutent dans le parenchyme hépatique par des canalicules sans parois propres; il s'agit essentiellement de l'espace intercellulaire qui forme un réseau en relation avec des canaux biliaires intra-hépatiques qui se réunissent en canaux (extra) hépatique aboutissant à un canal cholédoque qui conduit la bile jusqu'au duodénum.
Les voies biliaires intra-hépatiques.
Au niveau de chaque lobule hépatique, la bile sécrétée par les hépatocytes va être recueillie par un ensemble de petits canaux. La réunion de ces canaux va former le canal hépatique droit et gauche qui sortent du foie au niveau du hile du foie. Les canaux vont former un canal hépatique commun, qui devient extra-hépatique.
Les voies biliaires extra-hépatiques.
La voie biliaire principale.
Elle naît de la réunion des canaux hépatiques droit et gauche qui vont se réunir pour former le canal commun qui va se réunir au canal cystique pour former le canal cholédoque, qui va aboucher au duodénum par le canal de Vater.
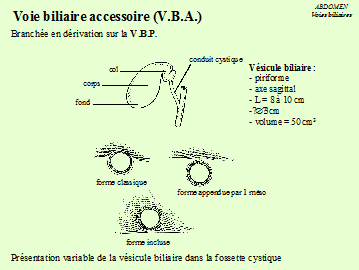 |
La voie biliaire accessoire(la vésicule biliaire).
C'est un réservoir où s'accumule la bile dans l'intervalle des périodes de digestion. L'orifice de sortie est le canal cystique.
Date de dernière mise à jour : 30/03/2018
Ajouter un commentaire