

ORGANOGENESE VERTEBRES 11
LE PANCREAS
Anatomie ( cas de l’homme )
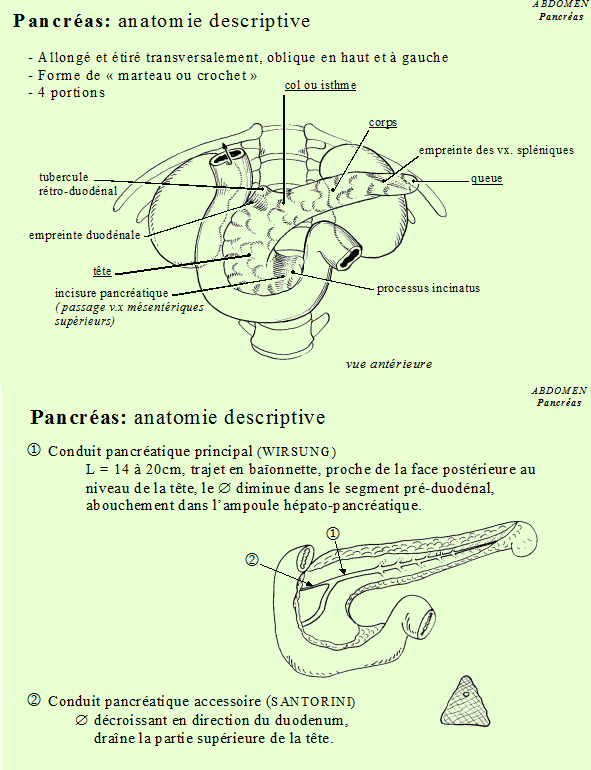
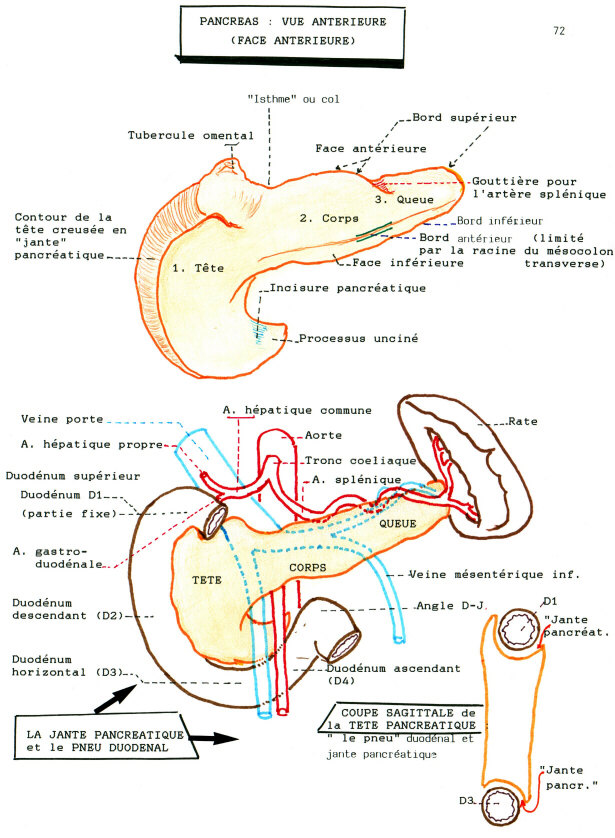
Chez l'homme le pancréas est un organe situé profondément dans l'abdomen, derrière l'estomac, devant et au dessus des reins. Il est formé d'une tête enchâssée dans le duodénum , d'un corps et d'une queue.
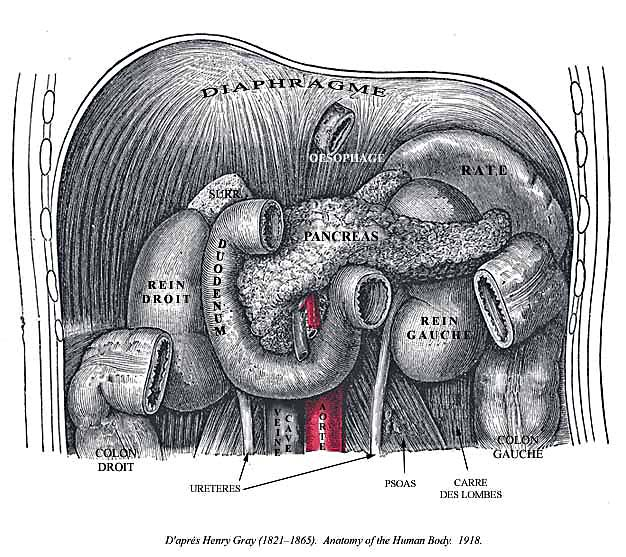
Figure : situation du pancréas
Fonctions
Le pancréas est un organe à sécrétion endocrine et exocrine c'est à dire qu'il fabrique des hormones déversées dans le sang et des enzymes digestives déversées dans le duodénum.
Sécrétion endocrine
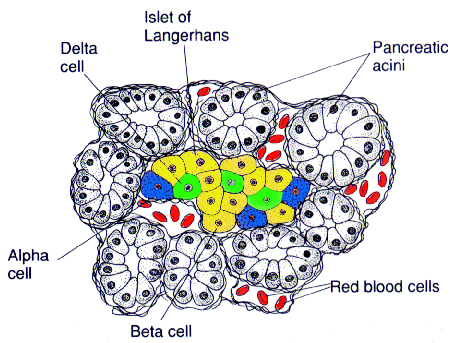
Les îlots de Langerhans, amas de cellules dispersés dans tout le pancréas, sécrétent des hormones : l’insuline surtout, qui est synthétisée par les cellules bêta, mais aussi le glucagon, la somatostatine et d'autres hormones produites par les cellules dites non bêta.
Sécrétion exocrine
Environ 80 % de la masse glandulaire du pancréas est responsable de la sécrétion exocrine c'est à dire des enzymes (20 environ , amylases, lipases, protéases …) responsables de la digestion des protéines, des triglycérides et des glucides alimentaires.
Structure histologique
Le pancréas exocrine est une glande acineuse composée à l’intérieur de laquelle sont dispersés les îlots de Langerhans
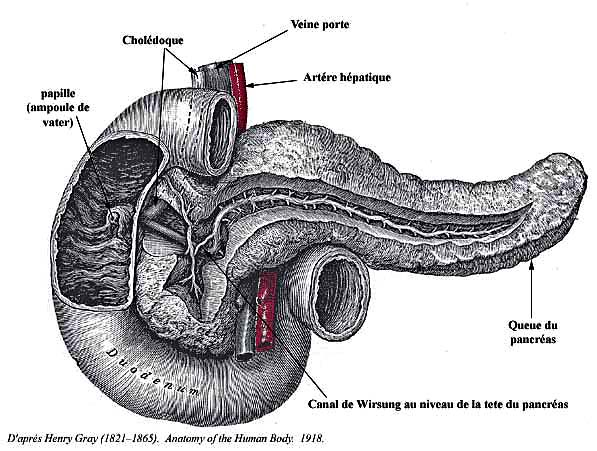
Canaux excréteurs : le canal de Wirsung et le canal de Santorini
-Le parenchyme glandulaire est divisé en lobules par de fines travées conjonctives issues de la capsule de l'organe ; ils contiennent des vaisseaux sanguins et lymphatiques ainsi que des nerfs.
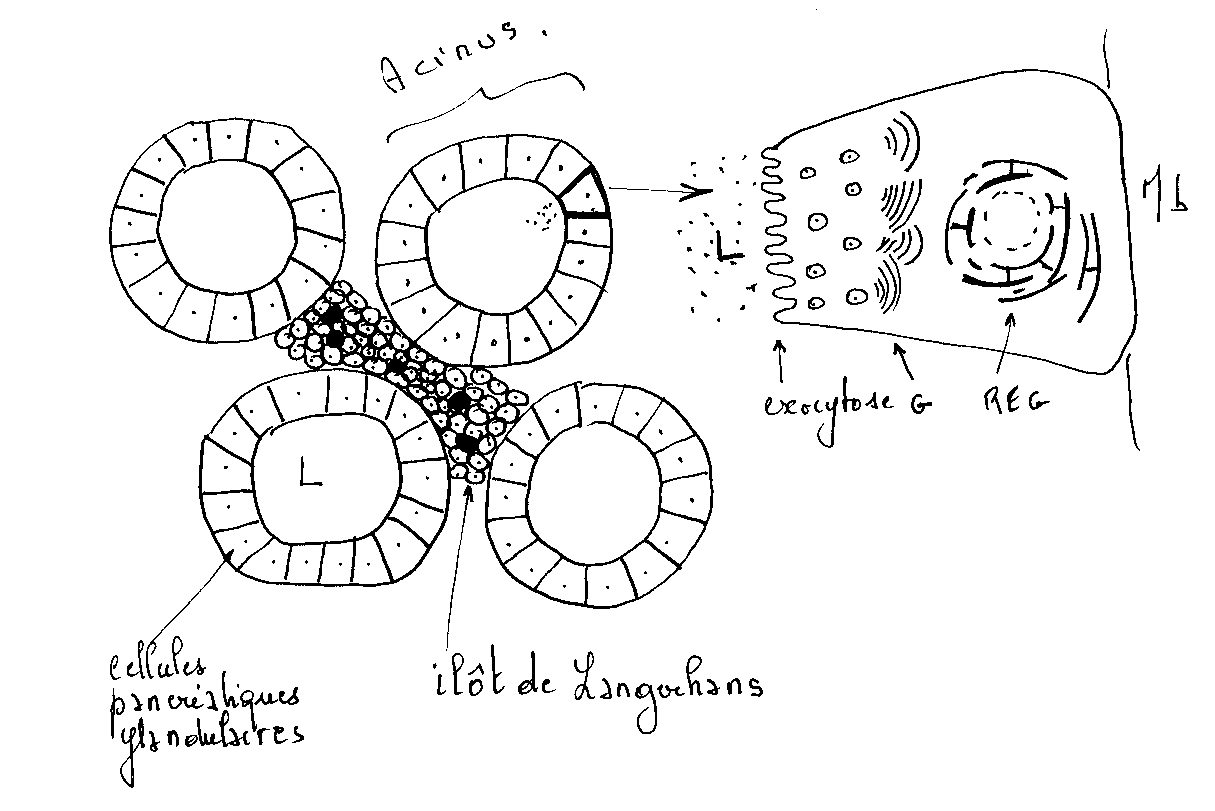
Fig : schéma simplifié, tissus et cellule pancréatique.
Il sera peut être utile de revoir la notion de cellule sécrétrice en biologie cellulaire.
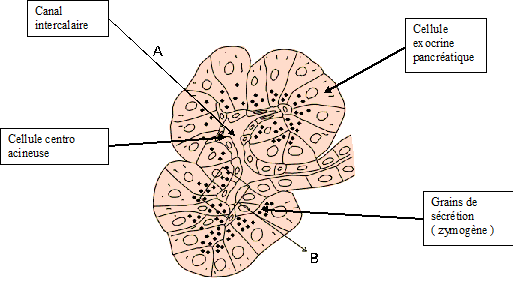
Fig : acinus pancréatique
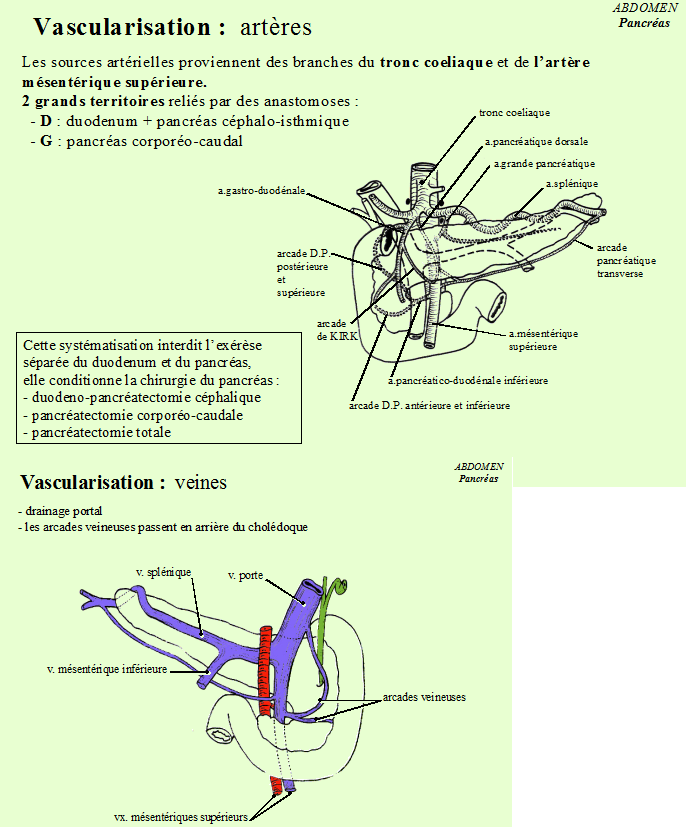
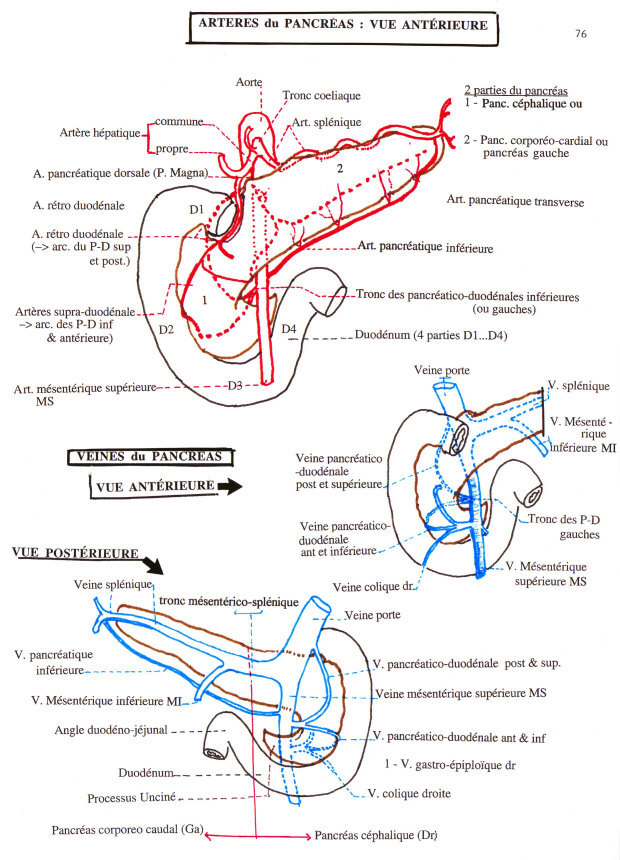
Vascularisation
Elle est assurée par les branches des artères hépatiques, spléniques et mésentériques supérieures.
Aspects comparatifs
Les cyclostomes et la plupart des actinoptérygiens ont un pancréas diffus, ne constituant pas un organe anatomiquement bien défini. Chez ces espèces, les cellules sécrétrices sont disséminées dans l'épithélium intestinal et le long des vaisseaux du mésentère, entre le foie et l'intestin. Elles s'infiltrent même dans le foie, le long des canaux biliaires. Chez les autres espèces, le pancréas se présente comme une volumineuse glande en grappe, composée de glandes acineuses. Ces glandes ont une structure voisine de celle des glandes salivaires. Elles sont constituées de cellules acineuses qui déversent leurs produits de sécrétion dans un canal intercalaire. Les cellules du canal se prolongent dans la lumière des acini en cellules dites centroacineuses, une caractéristique distinguant les acini pancréatiques des acini salivaires. Comme dans le cas des glandes salivaires, la sécrétion des cellules acineuses peut être modifiée par les cellules canalaires.
4.3 L’appareil respiratoire
L’appareil respiratoire a pour rôle d’assurer l’échange d’oxygène et de dioxyde de Carbone entre l’organisme et le milieu extérieur.
Il existe chez les Vertébrés, deux types d’organes respiratoires :
- Branchies.
- Poumons.
4.3.1 Les branchies
4.3.1.1 Embryologie
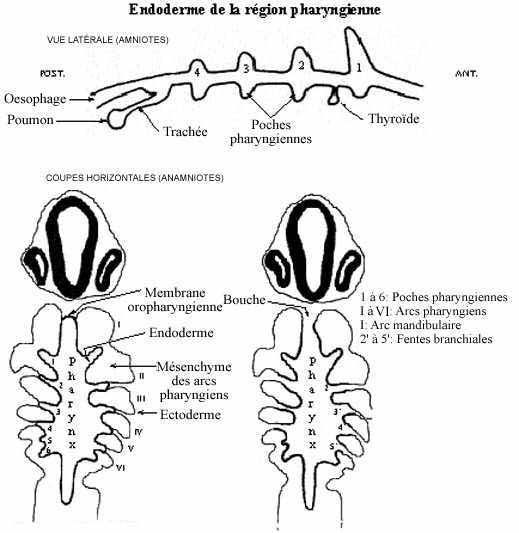
Latéralement, la paroi du pharynx décrit des évaginations digitées, les poches pharyngiennes (ou branchiales). Elles sont au nombre de 6 paires chez les anamniotes et de 4 paires chez les amniotes, et se développent selon un gradient antéro-postérieur. Elles se trouvent à délimiter les arcs pharyngiens (ou branchiaux ou viscéraux).
En s'évaginant, la poche pharyngienne repousse le tissu qui la sépare de l'ectoderme; ce dernier s'invagine et entre en contact avec l'endoderme de la poche pharyngienne; les deux fusionnent et forment la membrane branchiale. Sauf pour la 1ère et la 6ème paires, les membranes branchiales des anamniotes se perforent, résultant en la formation des fentes branchiales. Chez les Vertébrés aquatiques, des filaments branchiaux se développent à la surface des fentes branchiales; ce sont les branchies, surface respiratoire de ces animaux. Chez les amniotes, les poches peuvent ne pas du tout s'ouvrir vers l'extérieur; celles qui se perforent se referment secondairement. Comme des branchies ne se développent pas chez les amniotes, certains auteurs préfèrent éviter le terme branchial pour qualifier tant les poches que les arcs, et utiliser le terme pharyngien qui peut s'appliquer à tous les Vertébrés.
4.3.1.2 Note sur les branchies
Chez les Vertébrés, les branchies et les poumons semblent des organes bien différents. Pourtant leur évolution et leur morphogenèse prouvent qu'ils ne sont qu'une adaptation de l'épithélium pharyngien à deux environnements respiratoires différents. En quelque sorte, les poumons sont deux branchies invaginées. Il ne fait plus aucun doute que les branchies ont précédé les poumons dans l'histoire évolutive des Vertébrés, et qu'elles furent héritées sans grand bouleversement d'un ancêtre cordé primitif.
Les branchies des Vertébrés sont donc essentiellement des extensions latérales du pharynx richement capillarisées et tapissées d'une fine paroi cellulaire.
Les larves de nombreux Poissons et des Amphibiens laissent traîner simplement des branchies plumeuses évaginées dans leur milieu aquatique. Mais cette disposition est désavantageuse : les branchies sont exposées à toutes sortes de dommages, il n'est pas possible de faire mouvoir un courant d'eau adéquat à leur surface quand l'animal est au repos, et si les besoins en oxygène sont importants, ces branchies devraient être des touffes démesurées.
Chez les Poissons adultes, les branchies sont enfermées dans des cavités protégées du milieu extérieur par des clapets de peau et de muscles (Elasmobranches), finalement renforcés d'os (Téléostéens).
Très tôt au cours de l'embryogenèse chez les Vertébrés, 4 à 7 évaginations se forment à droite et à gauche du pharynx : ce sont les poches branchiales pharyngiennes. L'épithélium de ces poches - qui est de nature endodermique - arrive au contact de l'épithélium tégumentaire ectodermique et fusionne avec lui. Le mésenchyme qui jusque là avait occupé l'espace entre l'ectoderme et l'endoderme, se trouve réduit en colonnes verticales entre les poches successives : ce sont les arcs branchiaux. Les poches branchiales déchirent une communication avec l'extérieur : les fentes branchiales, qui apparaissent le long d'une ligne de fusion entre l'endoderme et l'ectoderme. Ainsi, l'eau peut être avalée par la bouche et expulsée par les fentes branchiales.
Pendant ce temps, le mésenchyme des arcs branchiaux différencie des supports cartilagineux, des vaisseaux sanguins
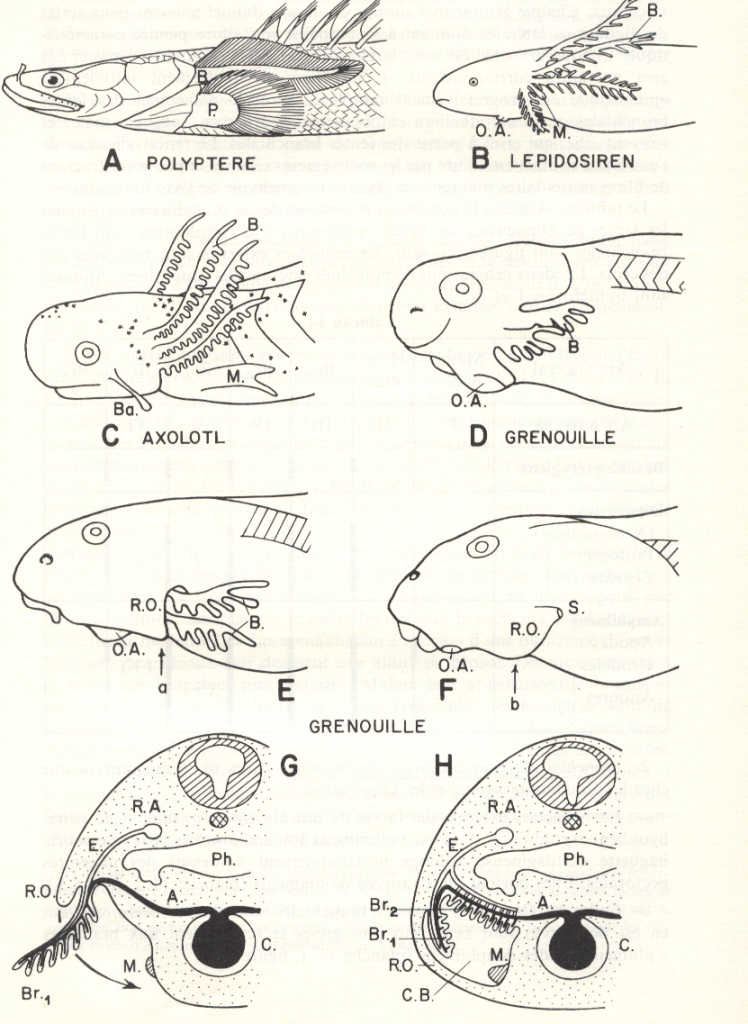

limite les fenêtres branchiales. Chez la plupart, c'est l'épithélium situé sur la face endodermique de la fente qui subira l'excroissance considérable caractérisant la différenciation des filaments branchiaux.
Les arcs branchiaux subissent une différenciation nouvelle : ils forment des denticules et des épines sur le bord exposé à l'intérieur du pharynx. Cet appareillage constitue un filtre qui empêchera le passage de nourriture, de débris, de matières solides, à travers les fentes branchiales et sur les délicats filaments branchiaux.
Enfin, les filaments branchiaux vont encore différencier une série d'excroissances : les lamelles branchiales.
La structure branchiale du Téléostéen adulte
Les Téléostéens sont les Poissons les plus abondants, et ils possèdent le système branchial le plus efficace ; ils réussissent à extraire 80% de l'oxygène contenu dans l'eau passant sur leur branchie, contre 50% chez les Elasmobranches. Ils serviront d'exemple.
On voit quatre paires d'arcs branchiaux au fond de la cavité bucco-pharyngienne; chacun des arcs supporte une double rangée de filaments. Les filaments branchiaux ne sont pas parallèles, mais s'écartent progressivement l'un de l'autre vers l'extrémité. Chaque double rangée de filaments portés par un arc est appelé holobranchie, et chaque rangée simple : hémibranchie. L'hémibranchie antérieure d'un arc vient s'interdigiter avec l'hémibranchie postérieure de l'arc qui précède. Donc, l'eau devra passer à travers un espace réduit, et obligatoirement sur les surfaces des lamelles secondaires. Ces lamelles constituent la membrane respiratoire vascularisée.
Les renforts squelettiques.
Les arcs branchiaux contiennent une pièce osseuse ou cartilagineuse disposée juste au-delà du filtre branchial de denticules. Ce renfort
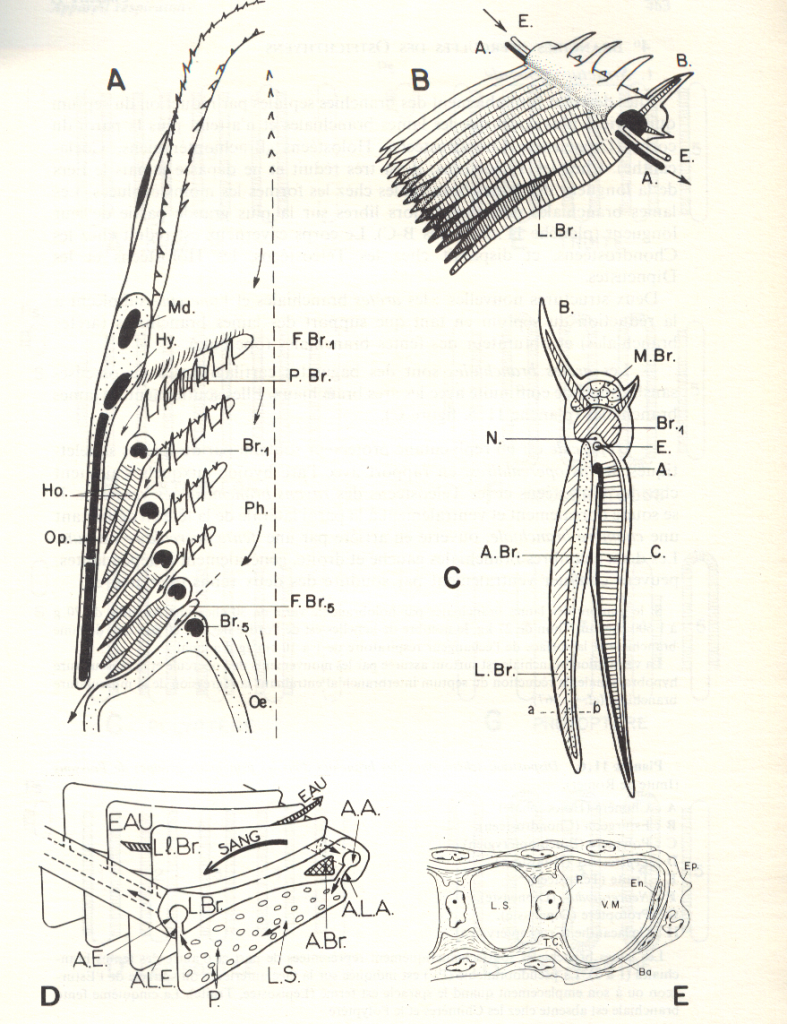

protégera ainsi les vaisseaux sanguins sous-jacents. Souvent, chaque filament branchial sera lui-même supporté par de minces renforcements squelettiques; des muscles y sont insérés pour l'adduction ou l'abduction des filaments.
L'irrigation et l'innervation.
Le sang propulsé du coeur dans l'aorte ventrale est réparti dans 4 paires d'arcs aortiques. De ces arcs aortiques surgit une succession d'artères afférentes, qui se dirigent vers l'extrémité des filaments branchiaux. Des branches pénètrent dans les lamelles, où elles se divisent finalement en réseau capillaire.
C'est un réseau capillaire très inhabituel. Ses canaux sont creusés dans une population de cellules dites "en pilastre". Ces cellules endothéliales fort particulières contiennent notamment des microfilaments contractiles qui permettent de régler le volume lacunaire et donc de modifier la microcirculation.
A la sortie du réseau capillaire, le sang est recueilli dans une branche qui le déverse dans l'artère efférente située dans le filament, puis celle de l'arc, et enfin l'aorte dorsale.
Des ramifications nerveuses provenant des nerfs crâniens IX et X parcourent les arcs branchiaux et prolongements pénètrent dans les filaments branchiaux.
L'épithélium de surface.
Le réseau capillaire creusé dans les cellules en pilastre est directement recouvert de l'épithélium tapissant les lamelles. Cet épithélium pavimenteux simple ou double présente souvent des microvillosités superficielles, dont le rôle consiste à retenir un film muqueux protecteur. On distingue parfois un espace entre les deux assises épithéliales, et la lymphe pourrait y circuler; à ce niveau, on retrouve aussi des granulocytes et des macrophages.
La lame basale de l'épithélium de surface est proche de celle des cellules en pilastre; ces lames basales se confondent en certains endroits. Localement, elles s'écartent et les fibres collagènes déposées dans ces espaces participent aux structures de soutien.
Dispersées dans l'épithélium recouvrant les lamelles, on observe des cellules à mucus et de grandes cellules acidophiles : les cellules à chlorure.
Ces dernières sont plus nombreuses dans l'épithélium stratifié logé à la base entre les lamelles, ainsi que dans l'épithélium proche de l'artère efférente branchiale. Les cellules à chlorure renferment de nombreuses mitochondries et de larges plages de réticulum lisse. Elles ont pour rôle de régler les échanges de NaCl entre le milieu intérieur et l'environnement, ce qui explique leur ultrastructure. Elles sont capables de pomper le sel contre un gradient de concentration; selon le milieu dulçaquicole ou marin, elles peuvent fonctionner dans l'un ou l'autre sens.
Chez les Téléostéens marins, l'apex de ces cellules contient une petite vésicule où le sel est concentré avant d'être déchargé. Par contre, cette vésicule n'existe pas chez les Téléostéens d'eau douce, qui extraient de leur milieu ambiant le peu de sels qu'ils y trouvent, pour le décharger dans leur milieu intérieur cette fois. La formation ou la mise en fonctionnement des cellules à chlorure peut être stimulée par l'administration de solutions salines dans le tube digestif. On conçoit très bien que l'on ne retrouvera pas de cellules à chlorure chez les Elasmobranches à peu près isosmotiques par rapport à l'eau de mer.
La pseudobranchie.
On pense que certaines cellules acidophiles, en fait très semblables à celles qui viennent d'être décrites, sécréteraient l'anhydrase carbonique. Elles sont nombreuses dans la pseudobranchie.
La première hémibranchie est toute seule sur son arc. Chez l'embryon, elle est irriguée par une branche de l'aorte ventrale, mais chez l'adulte, elle est nourrie par du sang oxygéné venant d'une dérivation de l'aorte dorsale; on présume donc qu'elle n'exerce aucune fonction respiratoire et c'est pourquoi on l'appelle "pseudobranchie".
Chez les Elasmobranches et certains Ganoïdes comme l'Esturgeon, la pseudobranchie est déployée et montre la même structure histologique que n'importe quelle branchie normale. Chez certains Téléostéens - comme le Cabillaud - les pseudobranchies sont recouvertes de l'épithélium buccopharyngien. Chez d'autres, des amas de cellules d'apparence glandulaire sont enrobés dans le conjonctif sous-épithélial; dans ce cas, on parlera de glandes pseudobranchiales.
Les cellules acidophiles que l'on y retrouve contiennent une dense population mitochondriale disposée en paquets ou en alignements, entourés d'un système de tubules de réticulum lisse très développé. La fonction de ces cellules n'est pas très claire : l'anhydrase carbonique qu'elles sécrètent scinde l'acide carbonique en eau et CO2, ce qui faciliterait peut-être la diffusion locale. D'autre part, on put mettre en évidence des relations vasculaires préférentielles entre la pseudobranchie et la vessie natatoire, ainsi qu'avec la glande choroïde de l'oeil.
Le cinquième arc.
La corbeille squelettique branchiale est formée de 5 arcs! Le cinquième et dernier ne porte pas de branchie.
4.3.2 Les poumons
Globalement, appareil respiratoire des Amniotes
4.3.2.1 Embryologie
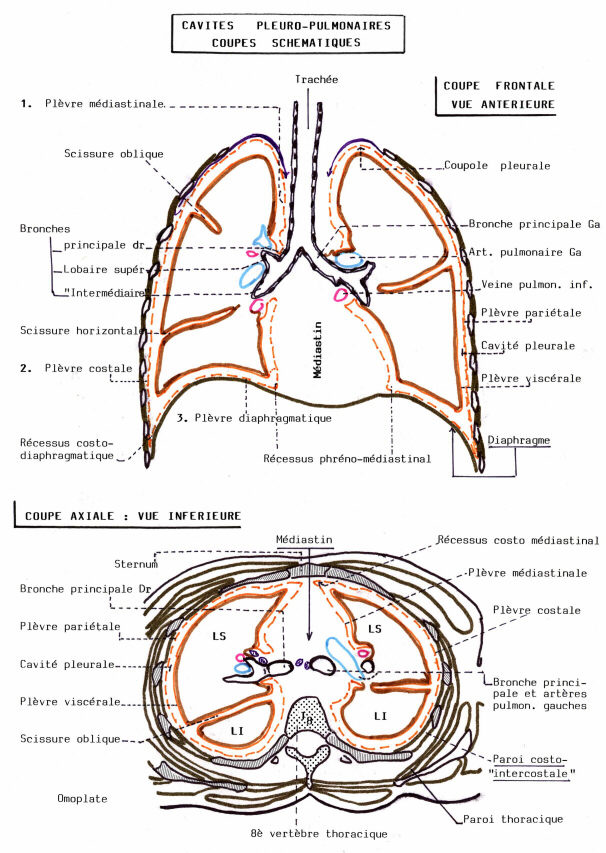
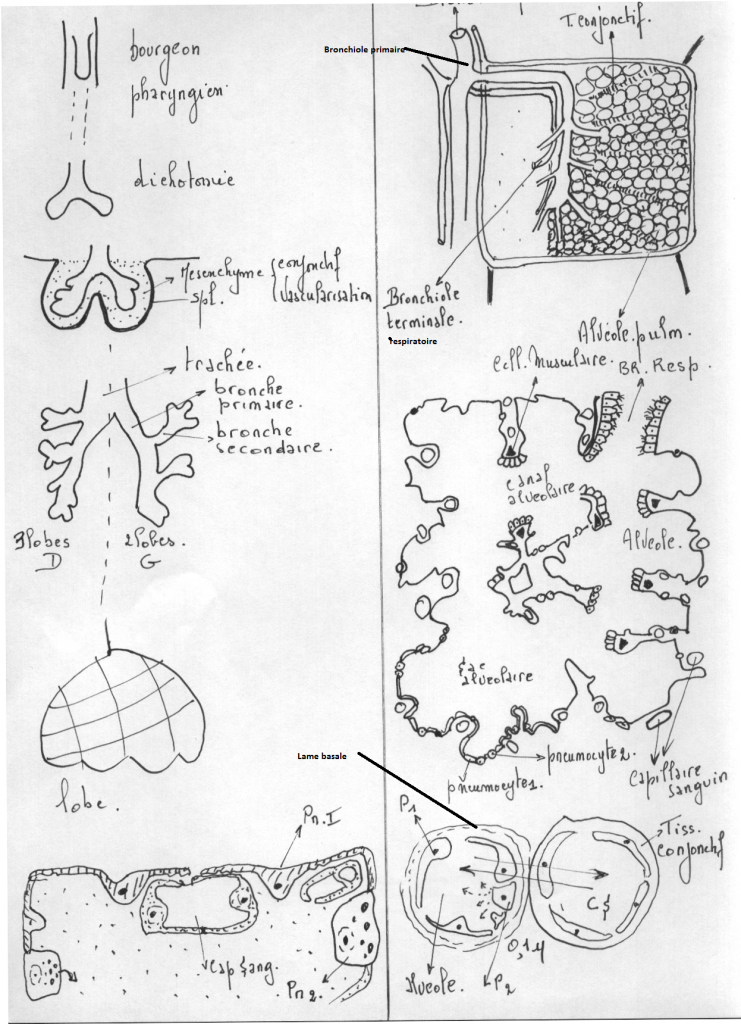
Postérieurement au pharynx, une évagination du plancher du tube digestif antérieur se dessine. Ce rudiment de la trachée bifurque en rudiments des bronches. Les rudiments des bronches s'allongent et se ramifient, leurs ramifications elle-mêmes s'embranchant et ainsi de suite. Ces embranchements des bronches et le tissu mésodermique qui leur est associé (dont des vaisseaux sanguins) constituent les poumons.
En fait,le développement de l’évagination repousse la splanchnopleure dont se détache un mésenchyme qui apportera le vascularisation.
La vessie natatoire des poissons se développe de façon comparable, mais sans ramifications.
La splanchnopleure refoulée constitue la plèvre viscérale.
La ramification , au fur et à mesure de son avancée individualise les bronches primaires, les bronches secondaires, les bronchioles primaires, les bronchioles respiratoires ( portant des alvéoles ) , les canaux alvéolaires portant les sacs alvéolaires cloisonnés en 2 ou 4 alvéoles.
4.3.2.2 Anatomie des poumons de mammifères
Les poumons sont deux organes pairs qui occupent la cavité thoracique. Du fait de la position du coeur, le poumon droit est plus développé que le poumon gauche. Le droit comprend 3 lobes (4 chez d'autres Mammifères que l'Homme), et le gauche, deux; chaque lobe reçoit une branche de la bronche primaire. La plèvre interne est étroitement appliquée à la surface pulmonaire.
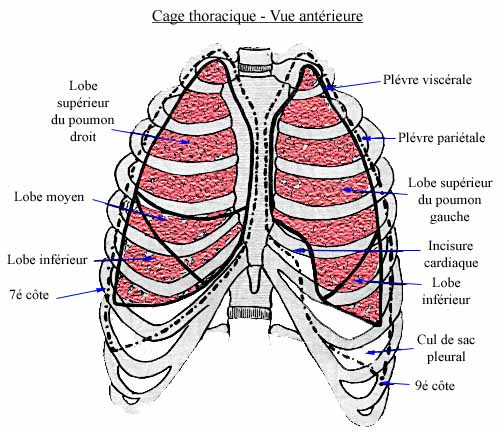
Poumons humains dans la cage thoracique
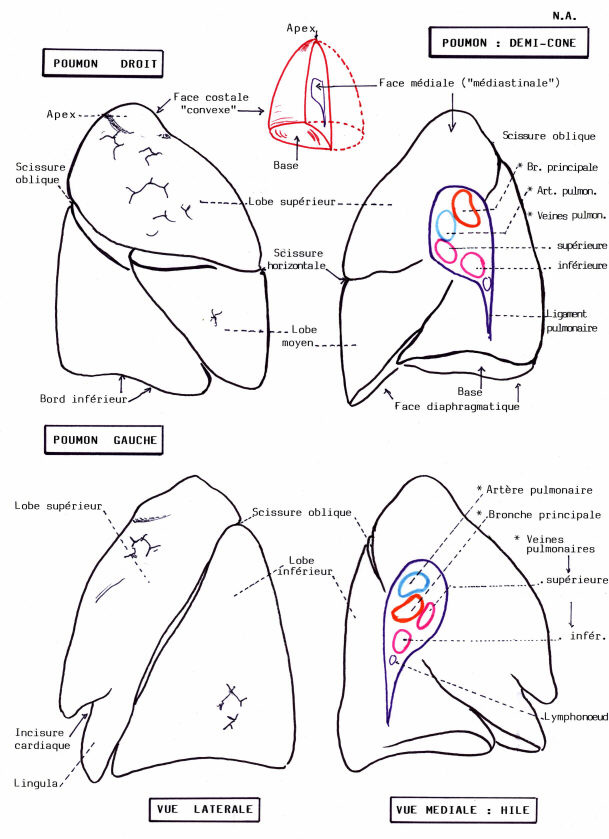
Chez les enfants, les poumons sont rose pâle, coloration due à l'irrigation sanguine. Avec l'âge, les poumons deviennent gris, à cause de l'accumulation des particules de carbone et autres substances minérales dans les cellules phagocytaires des septa conjonctifs
Cet assombrissement des poumons est particulièrement net chez les citadins, les fumeurs, et ceux qui sont exposés à la pollution de l'air.
Chacun des cinq lobes des poumons est divisé par de fins septa conjonctifs en un grand nombre de petits lobules, très visibles chez l'embryon, et moins chez l'adulte, sauf à la surface, surtout lorsque leur contour est souligné par les dépôts de carbone. Chaque lobule est pourvu d'une bronchiole.
Les bronches et les bronchioles.
La trachée se divise en deux branches, les bronches primaires qui pénètrent dans les poumons au hile, et s'y divisent à nouveau en deux à gauche, en trois à droite. Ces branches donnent encore les bronches secondaires, qui se dichotomisent en plusieurs ordres successifs (23-25 chez l'Homme).
Cette arborisation pulmonaire est caractéristique de l'espèce; des modélisations mathématiques de données morphométriques montrent des angles de dichotomie et des rapports de diamètres des conduits-fils déterminés le long de l'arbre, et correspondant à un modèle de fractales. Ce type de travaux est fort intéressant pour l'approche de la mécanique morphogénétique.
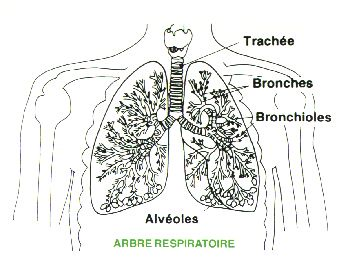
Chaque bronchiole terminale (intralobulaire) se poursuit en 1, 2, 3 ou plus bronchioles respiratoires, chacune donnant 2 à 11 canaux alvéolaires qui aboutissent au sac alvéolaire et à l'alvéole. On décrit aussi un atrium qui relie le sac alvéolaire et le canal.

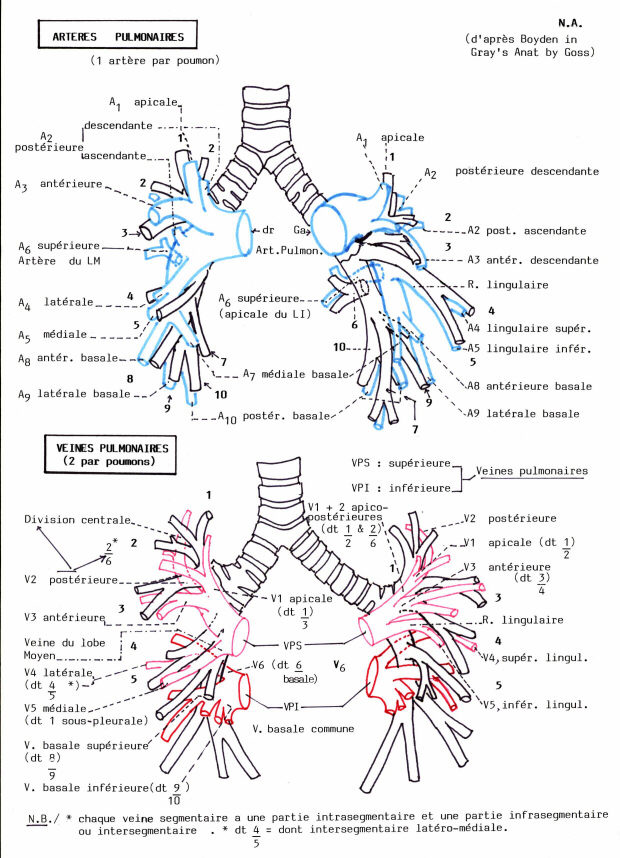
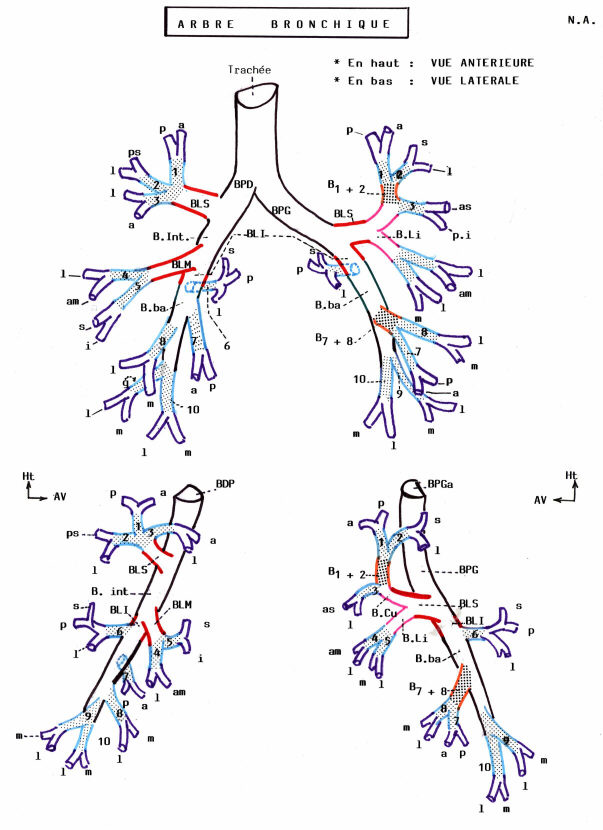
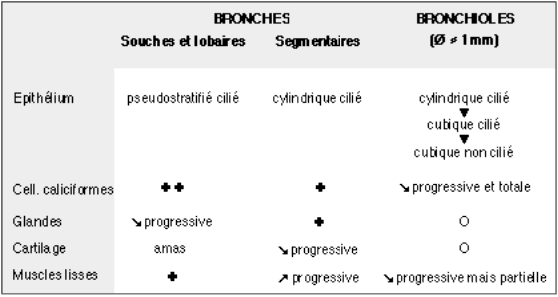
Hors des poumons, les deux bronches souches ont la même structure que la trachée bien que leur calibre soit inférieur et que les anneaux cartilagineux soient complets. Dans les poumons, les anneaux cartilagineux sont fragmentés. Au fur et à mesure que les bronches se subdivisent, la structure de leur paroi se modifie et ces modifications sont résumées dans le tableau précédent.
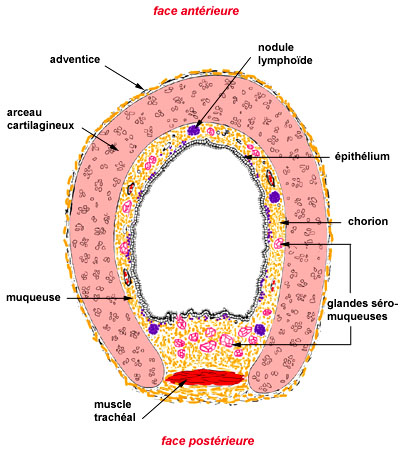
L’épithélium est encore pseudostratifié dans les bronches lobaires; il devient cylindrique cilié dans les bronches segmentaires et le reste dans ses subdivisions; dans les bronchioles dont le diamètre maximal est d’environ 1 mm, l’épithélium devient cubique puis perd ses cils. Les cellules caliciformes, encore nombreuses dans les bronches segmentaires, se raréfient progressivement et n’existent plus dans les dernières bronchioles.Dans le chorion, les glandes et les nodules de cartilage diminuent petit à petit et les bronchioles en sont dépourvues. Par contre, les cellules musculaires s’agencent en une musculeuse incomplète, le muscle de Reissessen, dès les bronches souches, augmentent relativement dans les bronches segmentaires et persistent en petit nombre jusque dans les dernières bronchioles.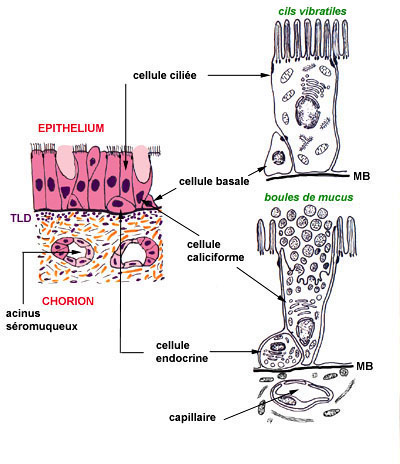
Le lobule
L'unité fonctionnelle est le lobule, constitué des structures desservies par une branche d'une bronchiole terminale : la bronchiole respiratoire, à laquelle succèdent le canal alvéolaire, l'atrium, le sac alvéolaire, l'alvéole. Le lobule comprend aussi des vaisseaux sanguins et lymphatiques, une innervation et l'emballage conjonctif. Chez le nouveau-né, le lobule est petit, la bronchiole respiratoire n'est pas encore développée, et les alvéoles ne sont encore que des diverticules de la paroi des canaux alvéolaires.innervation et l'emballage conjonctif. Chez le nouveau-né, le lobule est petit, la bronchiole respiratoire n'est pas encore
développée, et les alvéoles ne sont encore que des diverticules de la paroi des canaux alvéolaires
En coupe histologique, cette portion respiratoire du poumon apparaît comme un entrelac d'espaces clairs limités par de minces septa, et ponctué de canaux à parois épaisses : bronches et vaisseaux. Il faut mettre en oeuvre des techniques spéciales de fixation, d'injections, de coupes épaisses,... pour avoir une idée satisfaisante de la structure pulmonaire.
La bronchiole respiratoire
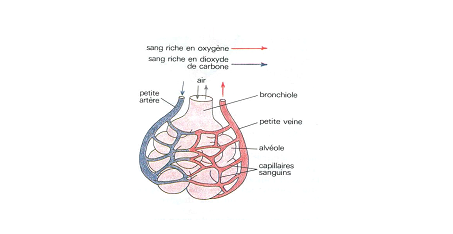
Une bronchiole terminale se divise en plusieurs branchioles respiratoires, dont le nom provient de la présence de quelques alvéoles pulmonaires disposées le long du conduit, du côté opposé à la branche de l'artère pulmonaire qui le longe.
La structure épithéliale se modifie. Déjà, dans la bronchiole terminale, les cellules ciliées ne sont plus accompagnées de cellules caliciformes à mucus mais d'un nouveau type de cellules sécrétrices, dénommées cellules de Clara.
Le seul caractère morphologique constant de la cellule de Clara est son pôle apical qui bombe entre les cils des autres cellules. Il contient de nombreuses mitochondries ovoïdes et des granules denses volumineux. Le noyau de la cellule est basal, souvent irrégulier, et entouré de réticulum endoplasmique rugueux, certains subdivisent les cellules de Clara en trois groupes. Il s’agit vraisemblablement d’états fonctionnels différents.
La paroi conjonctive élastique comporte un treillis de muscles lisses, mais ne contient plus de cartilage.
Les canaux alvéolaires
La bronchiole respiratoire se disperse en 2-11 canaux alvéolaires. Ces tubes minces, tortueux, à paroi très discontinue, sont difficiles à visualiser en coupes ordinaires; ils sont plus évidents en coupe épaisse. Ils produisent plusieurs branches qui se divisent encore, et sont boursoufflés par les sacs très nombreux, on comprend que les ouvertures constituent la majeure partie de la surface du canal alvéolaire. Ce qui reste de la paroi est soutenu par du conjonctif, avec des faisceaux élastiques et du muscle lisse.
L’arbre bronchique contient aussi de petites cellules endocrines qui sont isolées dans son épithélium ou regroupées en petits amas appelés corps neuro-épithéliaux. Ceux-ci sont surtout localisés aux différents embranchements de l’arbre bronchique. Ils contiennent deux types de cellules : les unes sont en contact direct avec la lumière bronchique par leur pôle apical; les autres par contre, n’arrivent pas en contact avec celle-ci. La structure de ces cellules et leur sécrétion, comprenant des polypeptides ou des amines, expliquent leur appartenance au système APUD, qui sera vu avec le système endocrinien. Par l’action de leurs sécrétions sur les muscles lisses vasculaires et bronchiques, ces cellules endocrines participent à la régulation de la vascularisation et de la ventilation pulmonaire.
Les alvéoles
La microscopie électronique a définitivement mis en évidence la continuité de la paroi épithéliale limitant les alvéoles. Des cellules épithéliales dérivées de l'endoderme embryonnaire s'amincissent tellement au cours de la croissance du poumon, qu'elles ne sont plus visibles en microscopie photonique sauf à l'endroit où se situe le noyau; le reste de leur cytoplasme n'est plus qu'une couche d'une épaisseur inférieure à la limite de résolution du microscope photonique. L'épithélium repose sur une lame basale continue, qui le sépare de l'endothélium capillaire ( voir schéma).
Les pneumocytes
Les cellules épithéliales sont de deux types : celui qui vient d'être signalé, aminci (pneumocytes de type I, pneumocytes membraneux) et des cellules arrondies ou cubiques contenant des inclusions (pneumocytes de type II, pneumocytes granuleux).
Dans l’interstitium, espace entre l’épithélium et l’endothélium, on observe, suivant les endroits, de rares fibroblastes, des macrophages, des lymphocytes, des polynucléaires éosinophiles et des mastocytes. A l’origine de chaque alvéole, les fibres de collagène sont nombreuses et associées à quelques fibres élastiques.
Le surfactant
Le surfactant est un matériel lipoprotéique complexe, comparable à un détergent. Il diminue la tension superficielle du film liquide qui tapisse les cavités alvéolaires, ce qui empêche l’affaissement ou collapsus des alvéoles lors de l’expiration. Il renferme douze fois plus de phospholipides que de protéines. Les protéines les plus nombreuses sont des glycoprotéines et les phospholipides les plus importants sont des dérivés de la phosphatidylcholine ou lécithine.
Une fixation adéquate permet de distinguer les deux phases de l’enduit alvéolaire. L’épiphase est en surface un film de densité variable, considéré comme une couche monomoléculaire de phospholipides dépourvue de protéines. L’hypophase est aqueuse. Elle contient des protéines et occasionnellement des corps lamellaires. L’importance des deux phases varie en fonction des mouvements respiratoires..
Le pneumocyte de type II intervient dans la synthèse et l’excrétion des phospho-lipides de la partie active du surfactant. Ce n’est pas sa seule fonction. Il élabore aussi les glycoprotéines de l’hypophase, peut régénérer l’épithélium alvéolaire en se transformant en pneumocyte de type I et intervient par sa micropinocytose dans les échanges d’eau et d’électrolytes entre l’hypophase et l’interstitium.
Notons que l’hypophase n’est pas produite par les seuls pneumocytes de type II. Une part de ses protéines proviennent des cellules de Clara et son fluide est un transsudat à partir des vaisseaux.
Le surfactant n’est produit qu’au 7ème mois de la vie embryonnaire. Son insuffisance chez les nouveau-nés prématurés est la cause d’une détresse respiratoire.
L’endothélium capillaire
Les cellules endothéliales limitant les capillaires alvéolaires présentent une disposition que l'on ne trouve nulle part aileurs : deux zones structurellement - et fonctionnellement - différentes. Une zone avésiculaire (dépourvue de vésicules de pinocytose) située entre les cellules épithéliale de l'alvéole et une zone vésiculaire normale située vis-à-vis du tissu de soutien. La première est surtout concernée dans le transport bidirectionnel des gaz, tandis que l'autre remplit les fonctions endothéliales classiques, notamment les transferts de macromolécules vers les tissus avoisinants.
Schémas complémentaires
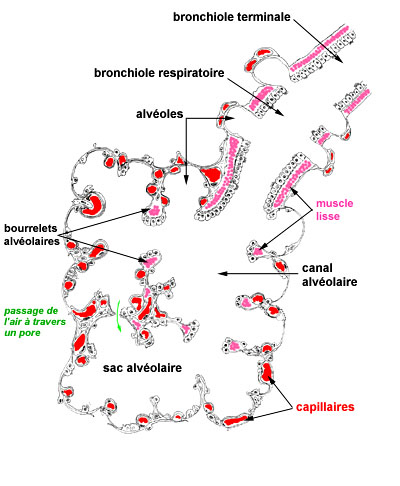
STRUCTURE DE L'ACINUS PULMONAIRE
Précisions sémantiques
l'acinus pulmonaire est l'unité morphofonctionnelle du parenchyme respiratoire; il est centré sur une bronchiole respiratoire qui se divise en quelques canaux alvéolaires; chaque canal alvéolaire se poursuit par 2 ou 3 sacs alvéolaires dans lesquels s'ouvrent de multiples alvéoles; la bronchiole respiratoire gère le fonctionnement de l'acinus pulmonaire qui est invalidé quand la lumière bronchiolaire est obstruée
. les bronchioles respiratoires sont des segments courts qui ont la même structure histologique que les bronchioles terminales; elles en diffèrent par la présence d'alvéoles pulmonaires s'ouvrant directement dans leur paroi, ce qui fait qu'en plus de la conduction de l'air, elles sont impliquées dans les échanges gazeux qui s'effectuent au niveau des alvéoles
. les canaux alvéolaires, larges et sinueux, n'ont pas de paroi propre : ils sont définis par les orifices des alvéoles et par les bourrelets alvéolaires; les bourrelets alvéolaires sont des épaississements annulaires comportant un épithélium cubique (cellules de Clara), des fibres élastiques et des fibres musculaires lisses circulaires dans un interstitium conjonctif; le bourrelet alvéolaire est une sorte de sphincter dont la contraction ferme le territoire alvéolaire situé en aval
. les sacs alvéolaires sont la confluence de plusieurs alvéoles dont ils ont la structure
. les alvéoles pulmonaires, chez l'Homme, sont de l'ordre de 300 millions; leur surface est de 100 m2 dont 60 à 80 sont le siège d'échanges gazeux entre l'air et le sang; les alvéoles sont l'aboutissement des voies aériennes; ce sont des cavités plus ou moins polygonales de 0,1 à 0,3 mm de diamètre (diamètre variable au cours du cycle respiratoire); ils s'ouvrent essentiellement dans les sacs alvéolaires, mais aussi dans les canaux alvéolaires et dans les bronchioles respiratoires; la paroi commune à deux alvéoles est la cloison interalvéolaire.
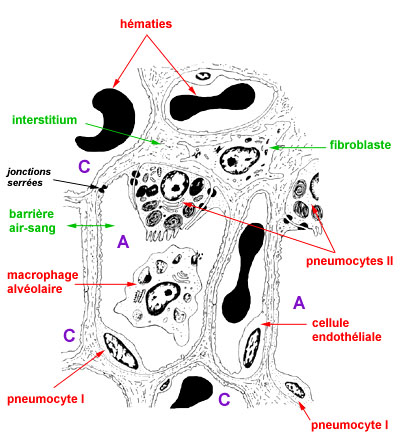
le schéma précédent montre l'aspect ultrastructural de 2 alvéoles (A) cernés par 6 capillaires (C), le septum interalvéolaire (interstitium) avec un fibroblaste, les cellules endothéliales et les pneumocytes I et II, un macrophage libre dans l'alvéole; on repère les corps lamellaires des pneumocytes II, les jonctions serrées (zonula occludens) entre les pneumocytes, et les lysosomes, phagolysosomes et corps résiduels du macrophage
Date de dernière mise à jour : 05/07/2021
Commentaires
-
 Loving the info on this site, you have done outstanding job on the content.
Loving the info on this site, you have done outstanding job on the content.
Ajouter un commentaire