

ORGANOGENESE 1 GENERALE ET ANATOMIE COMPAREE VERTEBRES
ORGANOGENESE
4. L’ORGANOGENESE
Après la gastrulation comportant le processus de neurulation ( voir rubrique embryologie descriptive ), on se trouve en présence d’un embryon triploblastique qui est :
- libre chez les Vertébrés à oeuf hétéro-lécithe.
- appliqué sur le vitellus chez les Vertébrés à oeuf télo-lécithe.
- enfermé dans le trophoblaste chez les Vertébrés à oeuf alécithe.
But de l’organogénèse : Que vont devenir les trois feuillets de l’embryon.

4.1 L’ectoblaste

4.1.1 Le neuroblaste
4.1.1.1 rappels neurulation
Evolution du tube neural
Les deux extrémités du tube neural ne se referment pas immédiatement mais demeurent soudées à l’ectoderme général et ouvertes sur l’extérieur, les neuropores antérieur et postérieur. Chez les amniotes les neuropores permettent le passage de liquide amniotique dans le neurocoele pendant un certain temps. Normalement ils se referment, le postérieur puis l’antérieur. Si le neuropore postérieur ne se referme pas, il en découle une pathologie appelée spina bifida, dont la sévérité dépend de la quantité de moelle épinière demeurant ouverte. Si le neuropore antérieur ne se referme pas, il en résulte une anencéphalie, léthale car le cerveau ainsi exposé dégénère et la voûte crânienne ne se forme pas. L’anencéphalie caractérise 0,1% des grossesses humaines.
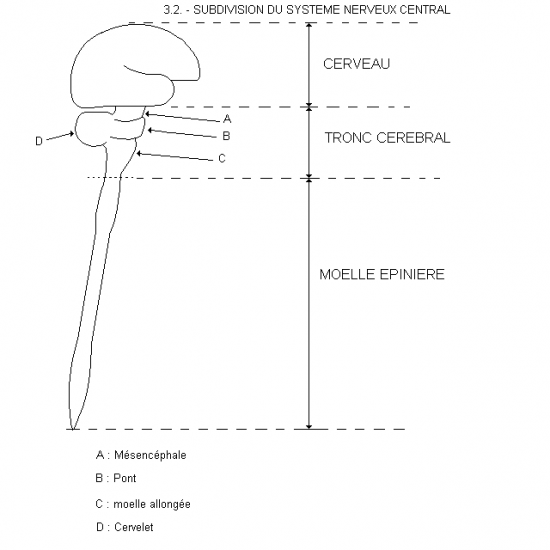
Des changements locaux du tube neural se produisent. Dans la portion céphalique le tube neural, élargi dès sa formation, se resserre en deux endroits et délimite 3 vésicules primaires: prosencéphale, mésencéphale, rhombencéphale. Postérieurement, le tube neural demeure un simple tube s’amincissant caudalement, la future moelle épinière.
Au moment où la portion caudale du tube neural se forme émergent les vésicules optiques, deux évaginations latérales du prosencéphale. Deux pliures ventrales se dessinent: la flexure cervicale, entre le rhombencéphale et la moelle épinière, et la flexure céphalique, entre le mésencéphale et le rhombencéphale. L’embryon devient replié sur lui-même. Ces repliements, du moins cela est connu de la flexure cervicale, ne sont pas intrinsèques au tube neural mais passifs; ils obéissent au mouvement autonome de repliement du coeur, dont nous discuterons dans un autre chapitre.
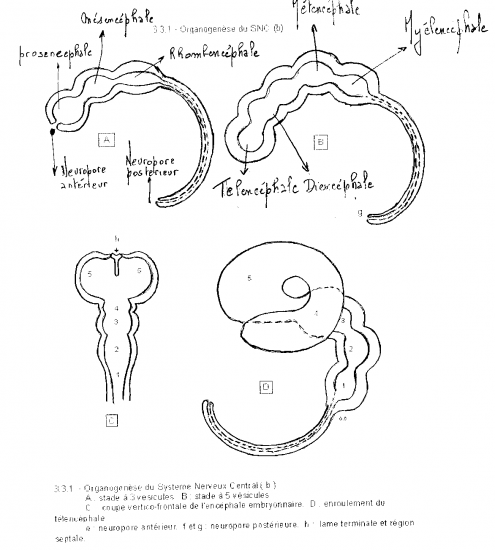
Par constriction, le rhombencéphale se subdivise en deux: myélencéphale et métencéphale.
Le myélencéphale deviendra la moelle allongée, continuation rostrale relativement peu différenciée de la moelle épinière, et le métencéphale donnera naissance au cervelet et structures pontiques associées.
Le mésencéphale ne se subdivise pas; il comprendra une portion du tronc cérébral (tegmentum) et le tectum chez les sous-mammaliens ou les corps quadrijumeaux chez les mammifères.
Le prosencéphale se subdivise en deux par constriction: diencéphale et télencéphale.
Le diencéphale formera le thalamus, le sous-thalamus, l’hypothalamus et l’épithalamus. C’est au diencéphale, plus précisément au thalamus, que sont reliées les vésicules optiques.
Le télencéphale formera les noyaux profonds et les hémisphères cérébraux.L’épithélium de la plaque neurale, de la gouttière neurale et du tube neural initial est simple cylindrique ou pseudostratifié.
4.1.1.2 Différentiation des neurones
Toutes les cellules sont en activité mitotique intense. Les cellules du tube neural se divisent tellement rapidement que l’épithélium devient stratifié. Certaines cellules cessent éventuellement de se diviser et migrent vers la périphérie du tube.
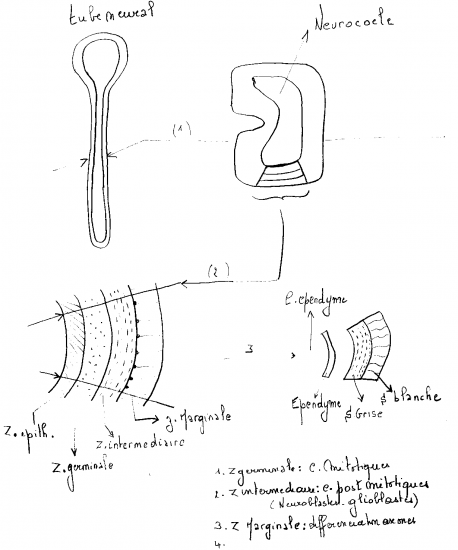
La localisation préférentielle des cellules mitotiques et postmitotiques autour du neurocoele définit deux zones dans la paroi du tube neural: zone germinative et zone intermédiaire ou manteau. Les cellules postmitotiques de la zone intermédiaire, encore non différenciées, appartiennent à deux lignées: neuroblastes, précurseurs des cellules nerveuses proprement dites, et glioblastes, précurseurs des cellules gliales. La zone germinative s’amincit à mesure qu’un plus grand nombre de ses cellules deviennent post-mitotiques et enrichissent la zone intermédiaire.
La zone germinative se réduit éventuellement à une seule assise de cellules qui deviennent ciliées: l’épendyme. Le neurocoele devient le canal épendymaire dans la moelle épinière et les ventricules dans le cerveau.
Dans la zone intermédiaire, les neuroblastes se différencient en neurones. D’abord, chacun élabore un prolongement cytoplasmique unique, l’axone, dirigé vers la périphérie du tube neural.
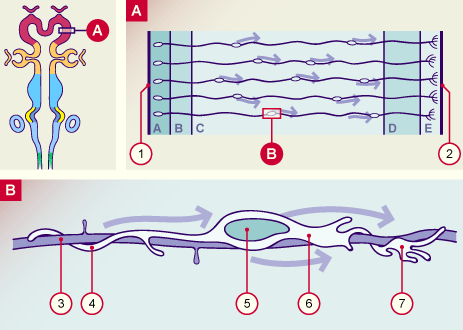
1 membrane limitante interne (cavité épendymaire)
2 membrane limitante externe (pie-mère)
3 prolongement cellulaire spécialisé de la glie radiaire
4 processus postérieur du neurone en migration
5 noyau du neurone en migration
6 neurone en migration
7 processus antérieur du neurone en migration
A Zone ventriculaire
B Zone subventriculaire
C Zone intermédiaire
D Plaque corticale
E Zone marginale
La plupart des neurones sont guidés dans leur migration de la zone ventriculaire germinative au cortex par les cellules gliales radiaires. Les études faites dans diverses espèces de mammifères montrent qu’une cellule gliale radiaire guiderait environ 130 neurones. Sur l’image du bas un neuroblaste en migration sur un prolongement de la glie radiaire a été schématisé. Observer le processus antérieur qui correspond au cône de croissance.
Le grand nombre d’axones émis à la périphérie du manteau créent une zone additionnelle au tube neural, la zone marginale. Elle s’épaissit à mesure qu’y parviennent les axones des neurones de la zone intermédiaire. Les neurones continuent leur cytodifférenciation, notamment par l’élaboration d’autres prolongements cytoplasmiques, multiples, les dendrites.
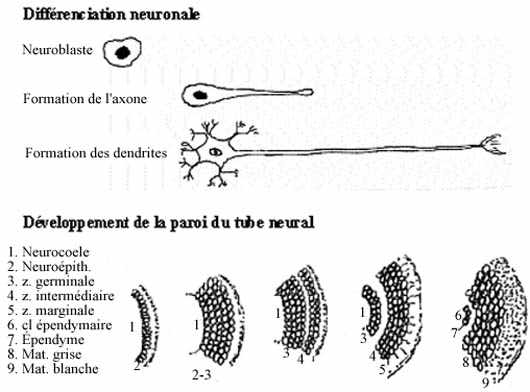
Certains glioblastes de la zone intermédiaire migrent vers la zone marginale et se différencient en oligodendrocytes qui forment un manchon isolant autour des axones: la myéline. Ce manchon à forte teneur lipidique donne une apparence blanche à la zone marginale, qui deviendra la matière blanche. Les glioblastes demeurant dans la zone intermédiaire se différencient en astrocytes qui protègent les neurones. La zone intermédiaire devient la matière grise (par "défaut", i.e., par manque de myéline). Les épendymocytes ne se divisent que pour pourvoir au remplacement des cellules gliales. Les neurones mûrs qui meurent ne sont pas remplacés.
Dans la moelle épinière, le myélencéphale et le métencéphale, les trois zones: épendyme, matière grise et matière blanche, se trouvent concentriques autour du canal épendymaire ou du ventricule. Dans la moelle, la matière grise prend la forme caractéristique d’un H. La moitié dorsale de ces régions, appelée plaque alaire chez l’embryon, est surtout consacrée aux fonctions sensorielles, i.e., la réception de l’information, et devient la corne dorsale. La moitié ventrale, appelée plaque basale, vouée aux fonctions motrices devient la corne ventrale. Les neurones qui s’y retrouvent sont généralement de plus grande taille. La racine ventrale, collection d’axones quittant le SNC pour innerver les muscles squelettiques, prend origine des motoneurones de la corne ventrale. Le terme "corne" est réservé à la moelle épinière.
Le taux des divisions mitotiques puis le mode de migration des neurones post-mitotiques et leur différenciation varient quelque peu d’un niveau à l’autre de la moelle épinière et, pour un niveau donné, tout autour du neurocoele. Ainsi l’épaisseur et la forme des zones intermédiaire et marginale ne sont pas tout à fait les mêmes aux niveau cervical, thoracique, lombaire, sacré ou coccygéen de la moelle, ou bien dans la plaque alaire et la plaque basale.
Dans les régions du cerveau rostrales au métencéphale (més-, di-, tél-encéphale), la façon dont se déroulent les migrations cellulaires et la croissance différencielle du tissu font que les matières grise et blanche s’entremêlent plus ou moins; elles ne sont pas concentriques autour du ventricule. Elles peuvent s’organiser en couches alternées, comme dans le cortex cérébral, le toit optique ou le cortex cérébelleux (ce dernier étant métencéphalique); ailleurs, des neurones à fonction commune se regroupent en amas appelés noyaux, séparés d’autres noyaux à fonctions différentes par des faisceaux d’axones. L’organisation des neurones en noyaux commence déjà à s’observer dans le rhombencéphale.
Les neurones établissent entre eux des contacts fonctionnels spécialisés appelés synapses qui utilisent des médiateurs chimiques appelés neurotransmetteurs.
Les vésicules optiques, évaginations des parois latérales du diencéphale, demeurent rattachées au diencéphale par les pédoncules optiques. Elles sont à l’origine de la rétine de l’oeil. Le développement de l’oeil entier sera étudié au chapitre suivant.
La formation du système nerveux comprend non seulement les étapes de divisions cellulaires, de migration et de différenciation (incluant la formation des synapses et la différenciation chimique - le choix de neurotransmetteurs), mais aussi des étapes de mort cellulaire, i.e., l’élimination sélective d’une proportion des cellules produites, de même que l’élimination d’axones et dendrites ainsi que de synapses établies.
4.1.1.3 Développement ultérieur de la crête neurale: SNP, squelette crânio-facial
Le développement de la crête neurale signifie grandement celui du système nerveux périphérique. À cause de sa nature périphérique, il nécessite la connaissance du développement et de la constitution adulte des cibles innervées: muscles striés squelettiques permettant les articulations du squelette, muscles lisses et leur organisation en organes, glandes exocrines. Le développement de la crête neurale ne signifie pas que celui du SNP, toutefois. Une grande proportion du squelette de la tête provient en effet de la crête neurale céphalique, non segmentée. Bien que neurectodermique d’origine, nous parlerons de sa formation dans le cadre du développement squeletto-musculaire du mésoderme. Nous nous contenterons, ici, d’énumérer les dérivés de la crête neurale.
Dérivés de la crête neurale:
- cellules nerveuses et gliales des ganglions sensoriels (crâniens et spinaux)
- cellules nerveuses et gliales des ganglions autonomes (sympatiques, parasympathiques)
- médulla de la surrénale
- cellules pigmentées de l’épiderme (mélanocytes, xanthophores, iridophores)
- cellules neuro-endocrines des voies digestives et respiratoires
- cellules parafolliculaires de la thyroïde, sécrétrices de calcitonine
- cellules de Type I du corps carotidien
- odontoblastes de la dent
- tissu conjonctif et squelettique de la tête
- tissu conjonctif des glandes dérivés de l’endoderme et de l’ectoderme pharyngiens
Tout en se divisant, les cellules de la crête neurale migrent vers leur site de résidence définitive. La direction de la migration semble déterminée par le substrat de migration, par contact sélectif des cellules avec la protéine fibronectine. Elles arrêtent leur migration et s’accumulent aux endroits dépourvus de fibronectine.
La destinée des cellules de la crête neurale dépend en partie d’elles mêmes, une détermination intrinsèque, mais aussi de l’environnement: tant du substrat de migration et des autres substances rencontrées lors de la migration que du site de résidence définitive dans l’embryon. Les parts intrinsèque et extrinsèque ne sont pas les mêmes pour toute la crête neurale. Les cellules de la crête neurale sont donc pluripotentes: une cellule peut se différencier en différents types selon sa localisation dans l’embryon; mais toutes n’ont pas le même degré de potentialité dans l’axe antéro-postérieur. Seules celles de la crête neurale céphalique peuvent produire le cartilage de la tête. La crête neurale crânienne transplantée dans la région thoracique formera le cartilage du tronc, qui est normalement produit par le mésoderme. La crête neurale thoracique transplantée dans la région crânienne ne forme pas de cartilage, qui est pourtant normalement formé par la crête neurale en cet endroit. Nous reviendrons sur ces notions au dernier chapitre.
Outre le tissu nerveux même, le système nerveux comprend d’autres tissus. Au cours du développement du tube neural, des cellules mésenchymales locales et le matériel intercellulaire associé forment trois couches autour de lui, les méninges, dont l’épaisseur et la solidité augmentent de l’intérieur vers l’extérieur. La méninge la plus interne, la pie-mère, est la plus délicate et s’insinue dans les fissures du SNC pour lui apporter les capillaires sanguins; la moyenne ou arachnoïde-mère véhicule les vaisseaux sanguins de taille intermédiaire; la dure-mère constitue une capsule conjonctive typique et solide. L’espace sous-dural renferme du fluide et du tissu adipeux qui servent d’amortisseur au SNC. Méninges et vaisseaux sanguins dérivent du mésoderme. Le tissu mésodermique n’entre jamais en contact direct avec les cellules nerveuses, incluant leurs prolongements (axones et dendrites), mais en est toujours séparé par des cellules gliales.
4.1.1.4 Chronologie du développement du système nerveux
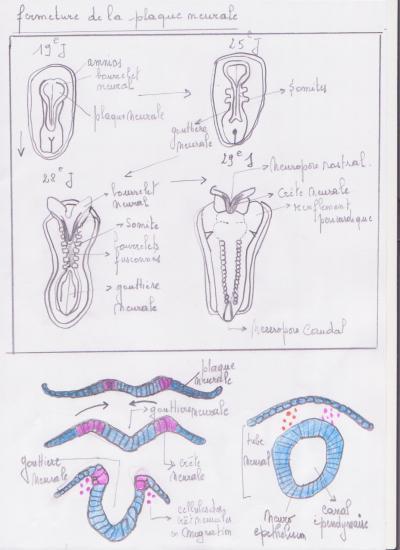
Neurulation : C’est le développement du tube neural primitif. ( rappel )
La plaque neurale et la gouttière neurale apparaissent au 17ème jour de la vie intra-utérine. La fermeture de la gouttière neurale apparaît au début du 21ème jour. La fermeture du neuropore antérieur ( future lame terminale ) se fait au 26ème jour et la fermeture du neuropore postérieur au 28ème jour. L’absence de fermeture du neuropore postérieur est à l’origine d’une malformation congénitale appelée spina bifida.
Développement de l’encéphale :
Le stade à trois vésicules se situe au début du 25ème jour. Le stade à cinq vésicules commence au 32ème jour. A deux mois et demi de la vie intra-utérine, les vésicules télencéphaliques recouvrent et encerclent complètement la vésicule diencéphalique. Dans l’épaisseur des parois du télencéphale se développent des formations cellulaires, progressivement volumineuses qui constituent le futur striatum
Parallèlement au développement morphologique, les structures fonctionnelles se mettent en place, en établissant les premiers réseaux synaptiques dans la substance grise.
Myélinisation
au niveau des nerfs périphériques : la myélinisation est effectuée par les gaines de SCHWANN. Elle commence au 4ème mois de la vie intra-utérine.
au niveau du système nerveux central : la myélinisation débute aussi au 4ème mois de la vie intra-utérine. Ainsi le faisceau vestibulo-spinal se myélinise au 6ème mois, le faisceau rubro-spinal au 7ème mois. Au niveau du faisceau pyramidal, la myélinisation se poursuit après la naissance et s’étend jusqu’à l’âge de 1 à 2 ans.
4.1.1.5 Exemple de chronologie du développement du système nerveux : homme
La plaque neurale et la gouttière neurale apparaissent au 17ème jour de la vie intra-utérine. La fermeture de la gouttière neurale apparaît au début du 21ème jour. La fermeture du neuropore antérieur ( future lame terminale ) se fait au 26ème jour et la fermeture du neuropore postérieur au 28ème jour. L’absence de fermeture du neuropore postérieur est à l’origine d’une malformation congénitale appelée spina bifida.
Développement de l’encéphale :
Le stade à trois vésicules se situe au début du 25ème jour. Le stade à cinq vésicules commence au 32ème jour. A deux mois et demi de la vie intra-utérine, les vésicules télencéphaliques recouvrent et encerclent complètement la vésicule diencéphalique. Dans l’épaisseur des parois du télencéphale se développent des formations cellulaires, progressivement volumineuses qui constituent le futur striatum
Parallèlement au développement morphologique, les structures fonctionnelles se mettent en place, en établissant les premiers réseaux synaptiques dans la substance grise.
Myélinisation
au niveau des nerfs périphériques : la myélinisation est effectuée par les gaines de SCHWANN. Elle commence au 4ème mois de la vie intra-utérine.
au niveau du système nerveux central : la myélinisation débute aussi au 4ème mois de la vie intra-utérine. Ainsi le faisceau vestibulo-spinal se myélinise au 6ème mois, le faisceau rubro-spinal au 7ème mois. Au niveau du faisceau pyramidal, la myélinisation se poursuit après la naissance et s’étend jusqu’à l’âge de 1 à 2 ans.
4.1.1.6 Histologie du système nerveux
4.1.1.6.1 Morphologie des neurones
Le neurone est constitué d’un corps cellulaire appelé "péricaryon" (càd. l’endroit où se trouve le noyau) et garni d’une multitude de prolongements ... De nombreux prolongements petits, courts, ramifiés qui sont les dendrites et des prolongements plus longs, généralement uniques qui sont les axones. Il est important ici de se rappeler que l’information peut être véhiculée le long des neurones, mais toujours dans le même sens ... Au niveau des dendrites, la dépolarisation va migrer de manière centripète vers le corps / préricaryon du neurone. Ces informations vont y être rassemblées / ordonnées / intégrées. Tandis que, au niveau de l’axone, cette dépolarisation va aller de manière centrifuge pour aboutir à la terminaison axonique qui est toujours une synapse.
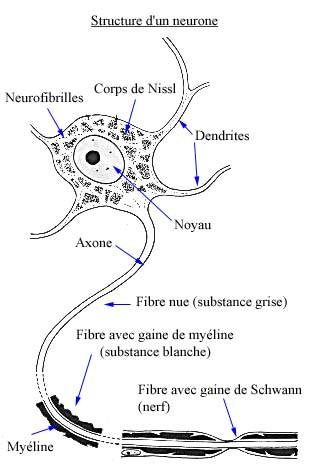
En bref, le neurone est constitué d’un corps cellulaire ( Pérycaryon ) et d’expansions cytoplasmiques plus ou moins ramifiées : les dendrites et l’axone.
4.1.1.6.1.1 . Le pérycaryon
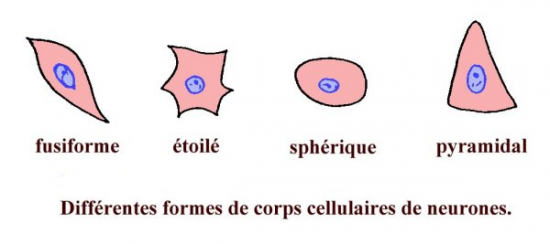
Celui-ci peut se présenter sous des formes variables :
- Forme étoilée ( cellules motrices de la corne antérieure de la moëlle )
- Forme pyramidale ( neurone du cortex )
- Forme sphérique ( cellules ganglionnaires)
- Formes globuleuse etc…
4.1.1.6.1.2 L’axone
Il n’y en a qu’un par neurone, il est ramifié à son extrémité, mais peut posséder des colatérales. On peut subdiviser l’axone en 4 régions :
• le cône d’implantation, en forme d’entonnoir il correspond à la région de départ de l’axone du corps cellulaire,
• le segment initial,
• le segment principal
• et la terminaison axonique ou neuronale
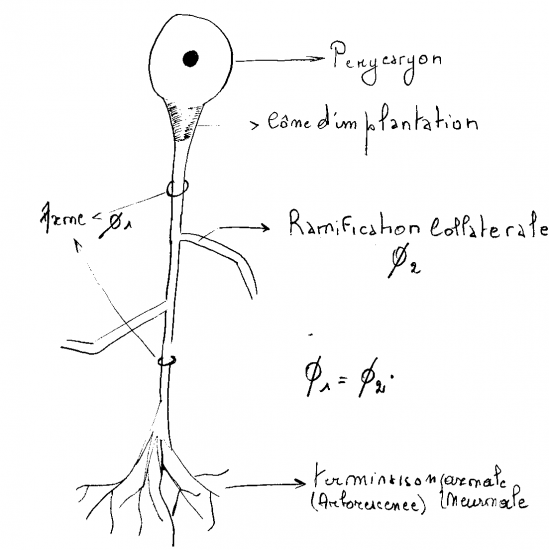
4.1.1.6.1.3 Les dendrites
Possédent des ramifications arborescentes nombreuses et de calibre irrégulier.
4;1.1.6.1.4 Classification morphologique des neurones
Classification basée sur le nombre de prolongements.
- Un seul prolongement : Cellules unipolaires ( cellules de la rétine ).
- Un seul prolongement : cellules pseudo-unipolaires ( ganglions rachidiens)
- Deux prolongements : cellules bipolaires ( épithélium olfactif)
- Nombreux prolongements : cellules multipolaires ( cellules étoilées des cornes de la moëlle épinière, cellules pyramidales du cortex cérébral).
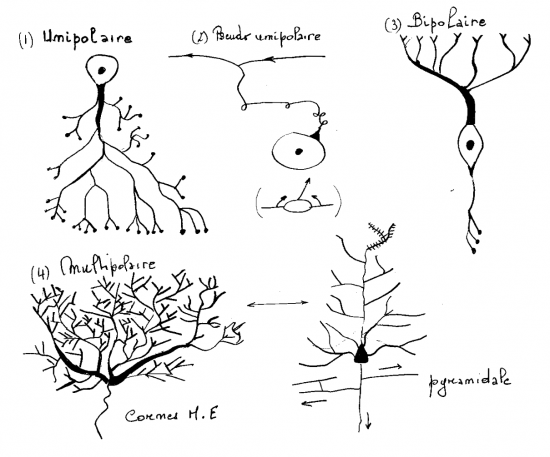

4.1.1.6.2 Cytologie du pérycaryon
PERICARYON = CENTRE FONCTIONNEL
NOYAU arrondi, riche en euchromatine, nucléole(s) hypertrophié(s)
dérépression génique, forte activité transcriptionnelle
CORPS DE NISSL = amas de réticulum granulaire et de Polysomes
Forte activité traductionnelle : synthèses protéiques intenses (enzymes hyaloplasmiques, turnover des protéines du cytosquelette, des canaux ioniques et récepteurs membranaires, sécrétion des neurotransmetteurs, etc)
APPAREIL DE GOLGI très développé
Nécessaire à la neurosécrétion et synthèses des vésicules synaptiques
MITOCHONDRIES
Fournir suffisamment d’énergie chimique utilisable pour les synthèses ( ATP…)
NEUROFIBRILLES
Une neurofibrille est faite de l’agrégation de deux neurofilaments torsadés. Les neurofilament sont constitués d’une molécule de méromyosine lourde.
Diamètre 50 Angström
Remarque : les microfilaments présents sont faits d’actine et ont un diamètre variable
MICROTUBULES
INCLUSIONS : MÉLANINE, LIPOFUSCINE
MEMBRANE PLASMIQUE ET CONNECTIQUE : voir plus loin
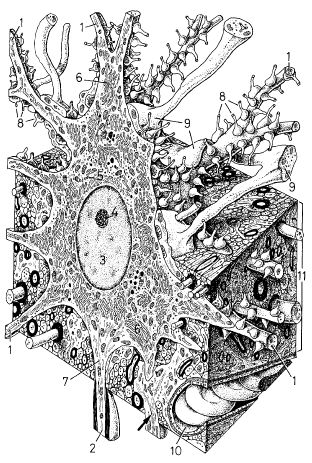
Légende :Coupe d’un pérycaryon
Plusieurs dendrites (1) et un axone myélinisé (2) se détachent du péricaryon de la cellule . Le noyau (3) riche en euchromatine possède un nucléole hypertrophié (4). L’appareil de Golgi est bien développé, avec plusieurs groupements dictyosomiques (5). Les citernes du réticulum endoplasmique granulaire se présentent sous forme de groupements qui constituent les corps de Nissl (6). Ils sont abondants dans tout le neuroplasme et même les dendrites. Ils sont par contre absents de l’axone et de son cône d’émergence , ou cône d’implantation (7) sur le corps cellulaire. Les mitochondries, les lysosomes et, à l’occasion, des grains de pigment (mélanine, lipofuscine) sont également présents.
Les neurofïlaments sont très nombreux dans le neuroplasme ; ils constituent le cytosquelette des cellules nerveuses.
Les centrioles sont absents du neuroplasme, puisque les cellules nerveuses ont totalement perdu leur capacité de division.
De nombreux boutons terminaux (8) et plusieurs prolongements astrocytaires (9) sont en contact avec le plasmalemme des cellules nerveuses. L’un de ces prolongements (flèche) contacte à la fois le neurone et un capillaire (10). Cette cellule pyramidale(cortex) a été dessinée de manière à se détacher de l’entassement du neuropile (11) que l’on a coupé en cube. Il est alors plus facile de suivre le trajet des prolongements des cellules gliales et nerveuses, et d’avoir une vision plus précise des unions cellulaires serrées qui constituent le tissu du SNC
(équivalent de grandissement : x 3 500

Légende : disposition des neurofilaments
Le noyau (1) est au centre du corps de la cellule nerveuse. Il est entouré par les dictyosomes de l’appareil de Golgi (2) anastomosés entre eux par de nombreux tubules fins (3). Les neurofibrilles (4) composées de neurofilaments, parcourent tous les secteurs du péricaryon et s’étendent dans les dendrites (5) et l’axone (6).
Le réseau fibrillaire confère une résistance mécanique nécessaire à l’architectue du neurone, mais il participe aussi au cheminement des structures vésiculaires et à celle des influx sans que le mécanisme soit totalement compris.
Les mitochondries (7) sont également très abondantes.
4.1.1.7 Organisation du système nerveux
Les métazoaires et en particulier les Vertébrés doivent coordonner les activités de leurs divers organes, ils disposent à ce titre de deux systèmes de communication :
- le système nerveux, chaine de cellules spécialisées appelées neurones, qui relient les organes entre eux
- le système endocrine, faisant intervenir des substances chimiques, les hormones , transportées par voie sanguine.
Quatre éléments interviennent dans la coordination nerveuse des Vertébrés :
- des récepteurs sensoriels capables de déceler certaines variations du milieu extérieur ou intérieur et de transmettre l'information correspondante à des centres nerveux.
- des centres nerveux chargés d'analyser cette information et d'élaborer une réponse.
- des organes effecteurs (muscles, glandes) chargés d'exécuter la réponse issus des centres nerveux.
- des nerfs reliant récepteurs sensoriels et organes effecteurs aux centres nerveux et par l'intermédiaire desquels l'information recueillie au niveau des brécepteurs sensoriels est transmise aux centres nerveux , et la réponse élaborée dans ces centres est communiquée aux organes effecteurs.
Centres nerveux et nerfs sont les deux constituants du système nerveux.
- Le point de vue anatomique
1. Le système nerveux comprend deux grands ensembles étroitement associés.
a) Le système nerveux central appelé ainsi non pas à cause de sa position qui est dorsale ( épineurien ), mais parce qu'il contient les centres nerveux. On l'appelle encore névraxe car il constitue un axe nerveux longitudinal s'étendant depuis la tête jusqu'à la queue, ou système cérébro-spinal car il se subdivise en :
- une partie antérieure cérébrale renflée, très différenciée , logée à l'intérieur du neuro-crâne : le cerveau ou encéphale
- une partie postérieure rachidienne ou spinale allongée et peu différenciée, logée à l'intérieur du canal rachidien de la colonne vertébrale : la moëlle épinière
b) Le système nerveux périphérique fait de nerfs mettant en rapport les centres du névraxe,d'une part avec les récepteurs sensoriels, d'autre part avec les organes effecteurs, suivant leur niveau d'émergence, ils se divisent en :
- nerfs crâniens issus de l'encéphale
- nerfs rachidiens ou spinaux issus de la moëlle épinière
On isole classiquement sous le nom de système nerveux végétatif un ensemble de centres et de nerfs, en rapport avec des fonctions de coordination de l'activité des viscères. Anatomiquement, ces centres sont inclus dans le névraxe et les fibres qui relient ces centres aux viscères accompagnent les nerfs crâniens ou rachidiens sur une partie de leur trajet. Elles s'isolent cependant pour former les nerfs symphatiques et parasymphatiques. Etant donnée l'importance du système nerveux végétatif, un chapitre particulier lui sera consacré.
2. Les récepteurs sensoriels . Ils sont souvent associés au système nerveux et étudiés avec lui. En principe, ils sont anatomiquement distincts du système nerveux central et reliés à lui par des nerfs sensoriels. Les photorécepteurs font cependant exception à cette règle, car ils représentent des territoires encéphaliques différenciés en plages sensorielles.
a) Les récepteurs sensoriels dits généraux sont dispersés dans l'organisme ( téguments, muscles, viscères ) et ne constituent jamais d'organes anatomiquement définis.
b) Les récepteurs sensoriels dits spéciaux sont au contraires groupés en organes sensoriels de forme définie ( rétine - oreille interne - organe olfactif ), auxquels peuvent éventuellement s'associer des structures annexes destinées à focaliser les stimuli sensoriels sur les récepteurs correspondant ( cristallin, oreille moyenne et externe ).
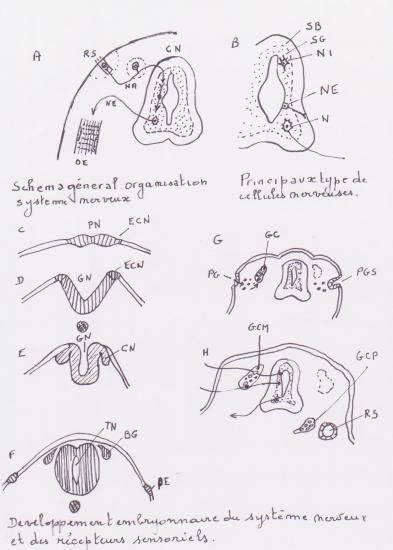
A . Schéma général de l'organisation du système nerveux
C.N. : Centre nerveux ; N.A. : Nerf afférent ; N.E. : Nerf efférent ; O.E. : Organe effecteur ; R.S. : Récepteur sensoriel.
B. Principaux types de cellules nerveuses
N. : Neurone ; N.E. : Cellule névroglique épithéliale ; N.I : Cellule névroglique intersticielle ; S.G : Substance grise ; S.B. : Substance blanche.
C-H . Développement embryonnaire du système nerveux et des récepteurs sensoriels. Coupes transversales très schématiques ( foetus humain ).
B.G. : Bandelettes ganglionnaires ; C.N : Crètes neurales ; E.C.N : Ebauche des crètes neurales ; G.C : Ganglions crâniens issus des bandelettes ganglionnaires ; G.C.M : Ganglions crânien mixte. Aux neurones ganglionnaires issus des crêtes neurales s'ajoutent des neurones ganglionnaires d'origine placodique. G.C.P : Ganglion crânien d'origine placodique : G.N. : Gouttière neurale ; P.E. : Placode épiblastique ; P.G. : Placode ganglionnaire ; P.G.S : Placode ganglionnaire et sensorielle ; P.N : plaque neurale ; Récepteur sensoriel d'origine placodique ; T.N. : Tube neural.
- Point de vue histologique
Comme nous l'avons vu ci-dessus :
Le système nerveux présente au niveau histologique une remarquable unité structurale liée à sa fonction essentielle : l'établissement de communications. L'élément cellulaire hautement spécialisé du tissus nerveux en rapport avec cette fonction est le neurone , caractérisé morphologiquement par ses prolongements cytoplasmiques, les fibres nerveuses, et fonctionnellement par sa capacité de réagir à une perturbation qui atteint sa surface (irritabilité ) et de transmettre l'excitation qui en résulte (conductibilité) à une autre cellule, nerveuse ou non, par l'intermédiaire d'une zone de contact, la synapse. Les neurones sont réunis par un tissus de liaison nourricier : la névroglie .
Sous-pages :
Date de dernière mise à jour : 30/03/2018
Commentaires
-
 I have noticed you don't monetize your website, don't waste your traffic, you can earn extra cash every
I have noticed you don't monetize your website, don't waste your traffic, you can earn extra cash every
month because you've got high quality content. If you want
to know how to make extra money, search for: best
adsense alternative Wrastain's tools
Ajouter un commentaire