
ORGANOGENESE 2
4.1.1.8 Les fibres nerveuses
Classiquement on dénomme fibre nerveuse,
un axone recouvert par sa gaine,
La gaine est dérivée de la crète neurale et correspond aux cellules de Schwann dans le système nerveux périphérique. Elle dérive du spongioblaste et correspond aux oligodendrocytes dans le système nerveux central.
Néanmoins dans les nerfs mixtes périphériques il est clair que les prolongements dendritiques des cellules sensorielles (prolongement allant de la périphérie jusqu'au corps cellulaire dans le ganglion spinal ; voir figure page 66) sont également recouverts par des gaines.
La définition classique est donc en partie erronée.
Par ailleurs, les fibres nerveuses forment des regroupements qui vont constituer :
---> les NERFS du système nerveux périphérique
---> les FAISCEAUX du système nerveux central
Les gaines nerveuses selon le mode de différenciation aboutissent à 2 types de fibres :
- LES FIBRES AMYELINIQUES, les plus simples (deux types, avec ou sans gaine de Schwann ).
- LES FIBRES MYELINISEES, plus complexes mais ayant de meilleures performances pour la transmission nerveuse
( deux types avec ou sans gaine de Schwann).
4.1.1.8.1 Fibres nerveuses myélinisées avec gaine de Schwann
Se rencontrent dans le système nerveux périphérique
Composition chimique de la myéline
Rapport L/P : 70/30 prépondérance des lipides
- Phosphatidyl-choline
- Cérébrosides ( glycolipides contenant du galactose)
- Sphyngomyéline ( Choline phosphorylée )
- Cholestérol
Voir biologie cellulaire.
Conséquences : Plus de lipides excluent l’eau et les éléments hydrosolubles (Na+ et K+ entre autres ) bon isolant
Protéines : composition caractéristique aux points de vue :
- Quantitatif : moins de protéines
- Qualitatif : moins d’hétérogénéité
Ainsi, il n’y a pas de protéines canaux qui pourraient faciliter le passage des ions.
Essentiellement :
• Protéines basiques solubles dans les solvants acides.
• Protéines assez rares qui ont la propriété d’être solubles dans les solvants organiques ( Protéines protidolipidiques)
Les protéines restantes sont des enzymes, des glycoprotéines.

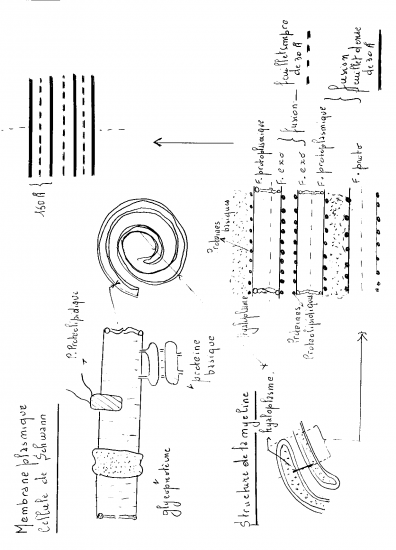
Formation et structure de la gaine de myéline
Dans ces fibres le cytoplasme de la cellule de Schwann s'enroule autour de l'axone. Au cours de ce processus les feuillets membranaires de la cellule s'unissent et forment un complexe lipido-protidique appelé gaine de myéline
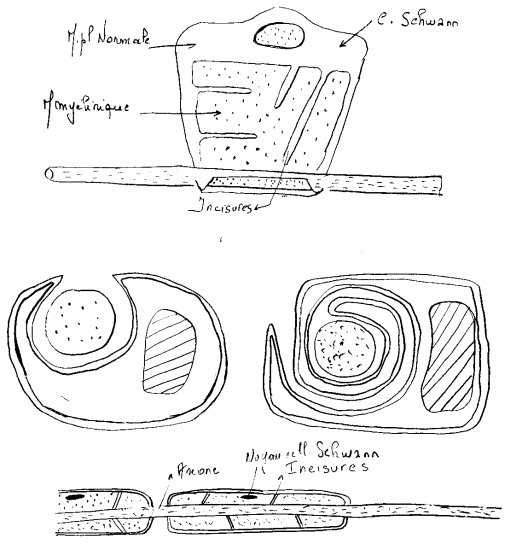
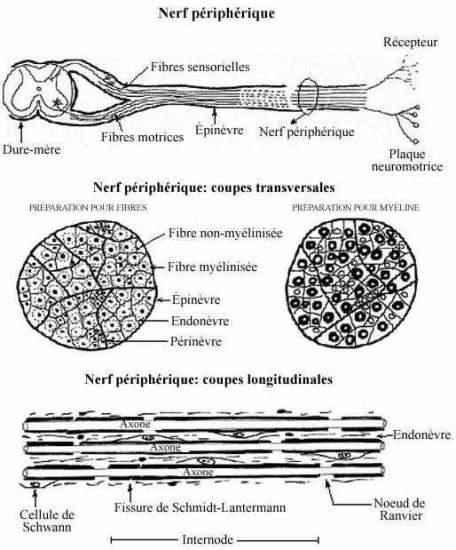
Comme nous le verrons sur les schémas ultérieurs, la gaine de myéline présente des interruptions, les étranglements ou noeuds de Ranvier, qui correspondent à l'espace entre deux cellules de Schwann adjacentes. La gaine de myéline présente aussi des zones moins compactes et obliques appelées incisures de Schmidt-Lanterman ; elles correspondent à des décollements des couches concentriques de la gaine de myéline.
La myéline n'est pas autre chose qu'un empilement de nombreuses couches de membranes cellulaires modifiées et appartenant à la cellule de Schwann.
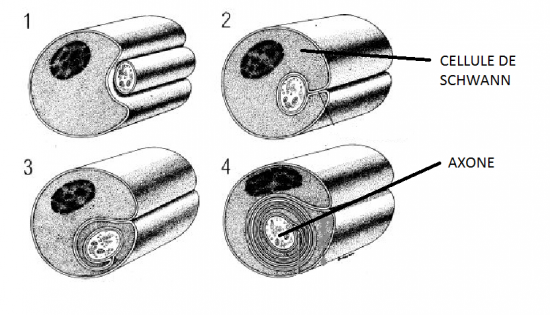
La cellule de Schwann appartient à la névroglie périphérique.
Le péricaryon émet des prolongements qui engainent l’axone puis s’enroulent sur lui.
La myéline présente des stries denses et obscures et des stries claires.
4.1.1.8.2 Fibres nerveuses myélinisées sans gaine de Schwann
Elles se rencontrent dans la substance blanche du système nerveux central et dans le nerf optique
Origine de la myéline
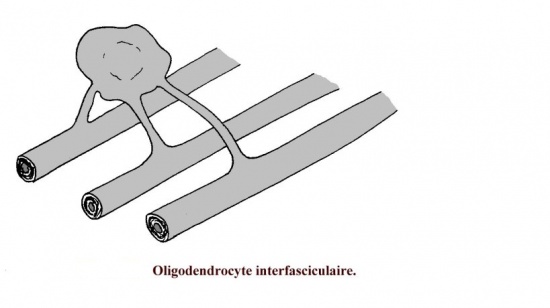
Enroulement d’un prolongement lamellaire d’un oligodendrocyte interfasciculaire
Ce type cellulaire appartient également à la névroglie ou cellules gliales de la substance blanche ou grise.
Définition de la névroglie
Ensemble de cellules non nerveuses d’origine ectodermique qui établissent d’étroit contact avec les neurones et leurs prolongements.
La névroglie forme une matière interstitielle. Elle prend aussi l'aspect d'épithélium de revêtement dans les cavités du système nerveux, par exemple le canal de l'épendyme dans la moelle ou les ventricules dans les hémisphères cérébraux. En outre, c'est la névroglie qui est à l'origine des gaines de SCHWANN. Contrairement aux apparences, ce tissu de soutien n'est pas un tissu conjonctif car il a la même origine embryonnaire que les neurones. En bref, c'est un tissu nerveux de soutien, de liaison et de revêtement. La névroglie joue un rôle trophique car elle est le milieu intermédiaire entre les vaisseaux et les neurones. De plus elle joue un rôle sécrétoire, car elle forme des pelotons épithéliaux appelés plexus choroïdes et elle intervient dans la sécrétion du liquide cérébro - spinal ( ou liquide céphalo-rachidien ).
La névroglie des centres nerveux comporte la névroglie épithéliale (épendyme, paroi ventriculaire et plexus choroïdes) et la névroglie interstitielle (astrocytes, oligodendrocytes et microglie).
Les épendymocytes forment un épithelium cubique ou prismatique simple revêtant les cavités ventriculaires du névraxe.

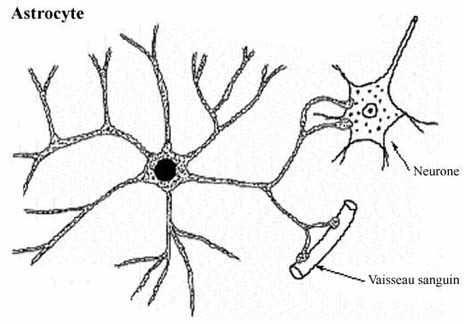
Les astrocytes sont des cellules étoilées avec de multiples expansions cytoplasmiques entourant les axones et les neurones voisins. Ils possèdent également des pieds vasculaires se terminant sur les capillaires (sustratum de la barrière hématoencéphalique). D'autres prolongements astrocytaires sont en relation avec les espaces lepto-méningés. En microscopie électronique, les astrocytes se caractérisent par leur richesse en filaments intermédiaires (contenant des quantités variables de protéine gliofibrillaire acide ou GFAP) et par leur richesse en grains de glycogène.
On distingue classiquement :
-les astrocytes de type 1 ou protoplasmiques, situés dans la substance grise, riches en GFAP et possédant des prolongements onduleux et courts
-les astrocytes de type 2 ou fibrillaires, situés dans la substance blanche, pauvres en GFAP et possédant des prolongements fins et longs.

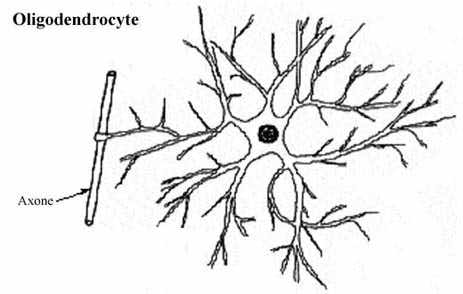
Les oligodendrocytes sont de petites cellules avec un halo clair correspondant à une étroite couronne cytoplasmique. Leurs prolongements sont plus courts et plus fins que ceux des astrocytes. On distingue :
-les oligodendrocytes satellites de la substance grise, associés aux péricaryons des neurones
-les oligodendrocytes interfasciculaires de la substance blanche, disposés entre les fibres nerveuses. Les oligodendrocytes de la substance blanche sont responsables de la myélinisation des fibres nerveuses à ce niveau (fibres nerveuses myélinisées sans gaine de Schwann).
Les cellules microgliales sont de petites cellules allongées, pouvant jouer un rôle phagocytaire.

La névroglie périphérique comporte les cellules de Schwann et les cellules satellites du ganglion spinal.
a - les cellules de Schwann entourent les fibres nerveuses dans les nerfs périphériques.
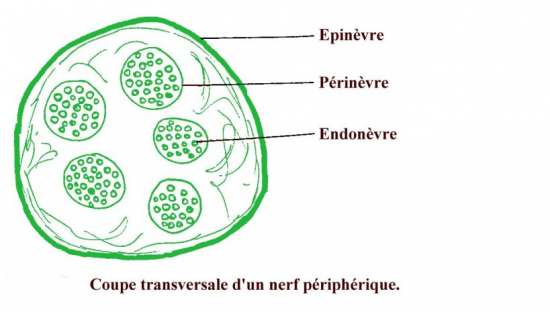
les nerfs périphériques contiennent de nombreuses fibres nerveuses, groupées en faisceaux et entourées de tissu conjonctif. L'organisation architecturale permet de distinguer :
-l'épinèvre, tissu dense périphérique limitant le nerf
-le périnèvre, tissu dense entourant les fascicules nerveux
-l'endonèvre, entourant individuellement chaque fibre nerveuse et riche en capillaires sanguins.
les cellules de Schwann entourent un ou plusieurs axones dans des dépressions de leur membrane plasmique. Leur organisation permet de distinguer deux types de fibres nerveuses périphériques : les fibres amyéliniques et les fibres myélinisées.
-les fibres nerveuses amyéliniques sont constituées par des faisceaux d'axones associés à une même séquence de cellules de Schwann.
-les fibres nerveuses myélinisées sont constituées par un seul axone associé à une même séquence de cellules de Schwann (fibres myélinisées avec gaine de Schwann). Cette organisation est surtout bien visible sur les préparations de nerf dissocié ou en microscopie électronique.
b -les cellules satellites forment une couronne de cellules aplaties entourant le corps cellulaire des neurones pseudo-unipolaires du ganglion spinal.
Remarque : alors que la cellule de Schwann peut se déplacer au cours de l’enroulement, la cellule oligodendrocyte est ancrée fermement par ses nombreux prolongements.
C’est donc la myéline nouvellement synthétisée qui doit pousser les nombreuses couches déjà formées , faisant ainsi avancer l’extrémité de la gaine
4.1.1.8.3 Fibre nerveuses amyéliniques avec gaine de Schwann encore appelées fibres de Remak
Au niveau du système nerveux symphatique
Il s’agit d’un faisceau de fins cylindraxes cheminant dans une gaine commune , succession ininterrompue de cellules de Schwann
Une même cellule de Schwann peut entourer plusieurs fibres nerveuses, plusieurs axones, presque les emprisonner, avec une seule couche de membrane plasmique. Autrement dit, l'enroulement ne se fait pas. Nous avons donc des fibres nerveuses non myélinisées. Ce qui veut dire que dans un nerf, l'une à côté de l'autre, nous avons des fibres nerveuses fortement myélinisées et des fibres nerveuses peu myélinisées. Quand on dit "non myélinisées", c'est à la limite de la vérité puisque de toutes façons, dans le système nerveux périphérique, il y a au-moins une membrane de la cellule de Schwann autour de la fibre nerveuse : c'est le début de la myélinisation mais qui ne va pas plus loin.
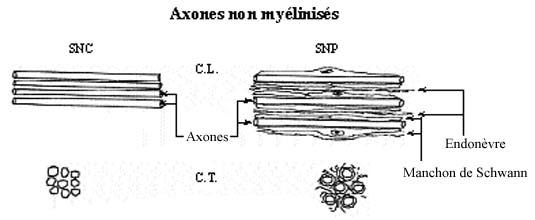
4.1.1.8.4 Fibres sans myéline, sans gaine de Schwann
Ce sont les fibres nues.
• Arborisations terminales d’un axone ou d’un dendrite
• Substance grise de l’encéphale ou de la moëlle
4.1.1.9 synapses
4.1.1.9.1 Les synapses interneuronales chimiques
Les synapses polarisent la transmission de l’influx nerveux dans les circuits de neurones.
L’influx nerveux se propage toujours dans un sens défini.
Définition
La synapse interneuronale est une région de contact entre deux neurones qui va assurer la transmission par voie chimique de l’influx nerveux d’un neurone à l’autre.
Classification
1° Premier groupe : l’élément présynaptique est toujours un axone :
• Synapse axo-somatique
• Synapse axo-axonique
• Synapse axo-dendritique

2° Second groupe : L’élément présynaptique, dendrite ou pérycaryon.
• Synapse dendro-somatique
• Synapse dendro-dendritique
• Synapse somato-somatique
Structure de la synapse
Grande diversité de synapses , mais unité de structure
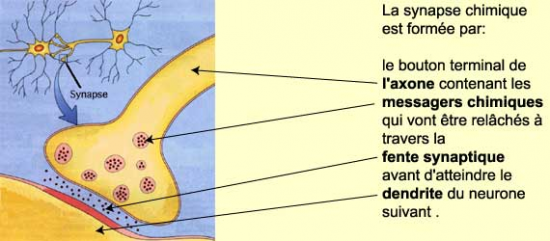
On distingue :
1) la zone présynaptique
- Vésicules présynaptiques
- Membrane et grillage présynaptique
Le grillage est un ensemble de projections denses à disposition hexagonale qui reposent sur la face interne de la membrane présynaptique.
Entre les projections denses , les espaces ont valeur de canaux ( diamètre 250 Angström ) ; les vésicules s’y engagent, atteignant ainsi la membrane postsynaptique
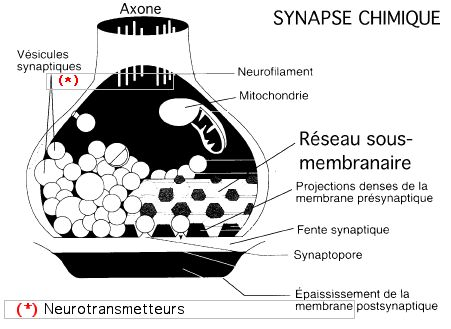
En dehors des mitochondries et du cytosquelette, les deux constituants les plus importants de l'élément présynaptique sont les vésicules synaptiques (dites aussi vésicules présynaptiques) et l'épaississement de la membrane présynaptique. Le feuillet interne de la membrane présynaptique apparaît en effet plus épais et plus dense aux électrons que le reste de la membrane plasmique du neurone. Cette densification membranaire correspond à une structure complexe appelée grille présynaptique, faite de l'arrangement régulier, trigonal, de projections denses reliées par de fins microfilaments et circonscrivant ainsi des emplacements où les vésicules synaptiques peuvent se loger individuellement. De petites dépressions (synaptopores) visibles à la face externe de la membrane présynaptique s'enfoncent en regard des emplacements vésiculaires situés sur l'autre face de la membrane.
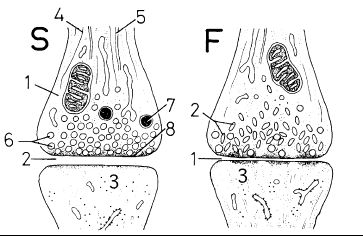
L'aspect général du renflement synaptique (1) est toujours identique. En bout d'axone on reconnait les microfilaments et microtubules (4, 5). La zone postsynaptique (3) est également très univoque. Mais on distingue différents types de synapses en fonction, soit de la nature vésiculaire (taille, forme, contenu), soit de l'aspect de la fente synaptique. Pour simplifier :
Synapses type S. Ce sont les plus répandues, avec des vésicules sphériques de taille variable. On distingue :
- les petites vésicules sphériques (6), à contenu clair ou dense selon le neurotransmetteur. Les plus répandues sont celles contenant de l'acétyl-choline. Les petites vésicules sont directement sécrétées dans la fente synaptique
- les grandes vésicules sphériques (7) avec un coeur dense séparé de la membrane par un halo clair périphérique. Elles contiennent préférentiellement des neuropeptides (et toujours de la chromogranine). Les grandes vésicules sont exocytées latéralement et non directement dans la zone synaptique.
Synapses type F. Dans ce cas les vésicules synaptiques sont applaties (2). Elles contiennent très souvent du GABA et correspondent à des synapses inhibitrices. Dans le type F la fente synaptique est différente ; elle est en particulier beaucoup plus étroite.
LA RICHE DIVERSITE DES NEUROTRANSMETTEURS EXPLIQUE LES CAPACITES D'ADAPTATION REGULATRICES A COURT TERME DU SYSTEME NERVEUX.
Nous fournissons ci dessous une ébauche de classification (en aucun cas à apprendre "par coeur").
1) LES NEUROTRANSMETTEURS «CLASSIQUES» (c'est-à-dire les plus anciennement connus) sont :
a) L'ACÉTYLCHOLINE,
b) LES MONOAMINES : catécholamines (dopamine, noradrénaline, adrénaline), sérotonine (5 hydroxy-tryptamine = tryptophane décarboxylé et hydroxylé), histamine ( = histidine décarboxylée).
2) LES PURINES : ATP, adénosine
3) LES ACIDES AMINÉS
- les uns excitateurs, comme le glutamate (50 % des synapses du SNC sont glutamatergiques) et l'aspartate,
- les autres inhibiteurs, comme le GABA (1/4 à 1/3 des synapses du SNC sont GABAergiques) et la glycine.
4) LES NEUROPEPTIDES. Ils se comportent plutôt comme des neuromodulateurs. Ils exercent une action de régulation au niveau de nombreux récepteurs extrasynaptiques, plutôt qu'au niveau de sites purement synoptiques.
On distingue :
- LES NEUROPEPTIDES NON OPIOÏDES. Les principaux sont : l'ocytocine et la vasopressine, les tachynines ou neurokinines (substance P, neurokinine A, neurokinine B, neuropeptide K, neuropeptide g), les endothélines, le vaso-intestinal peptide (VIP), la somatostatine, le neuropeptide Y, le calcitonin gene-related peptide (CGRP), la bradykinine, la cholécystokinine et autres peptides de la famille de la gastrine, la galanine, etc.
- LES PEPTIDES OPIOÏDES. Ce sont des agonistes endogènes naturels des récepteurs aux opiacés.
5) LE MONOXYDE D'AZOTE (NO) est également considéré comme un neurotransmetteur.
2) La fente synaptique
Cet espace de 250 à 300 Angström est rempli par un matériel extra cellulaire composé en partie de glucosamine glycane.

3) La zone post synaptique
Elle est fréquemment épaissie et sous tendue par des tonofilaments de kératine.
Le neurotransmetteur libéré dans la fente synaptique se fixe sur les récepteurs ionotropiques ou métabotropiques de la membrane postsynaptique.
Les récepteurs ionotropiques (ou récepteurs-canaux)
Leur ouverture est contrôlée par un neurotransmetteur. L'ouverture des canaux sodium, récepteurs de l'acétylcholine ou du glutamate, entraîne l'entrée de Na+ dans l'élément post-synaptique et par voie de conséquence une dépolarisation de la membrane de la cellule-cible et donc une excitation neuronale (synapses excitatrices). L'ouverture des canaux chlore, récepteurs du GABA ou de la glycine, entraîne une hyperpolarisation de la membrane de la cellule-cible et donc une inhibition neuronale (synapses inhibitrices).
Les récepteurs métabotropiques
A la différence des récepteurs ionotropiques, les récepteurs métabotropiques sont séparés des canaux ioniques dont ils règlent le fonctionnement, le couplage étant assuré par une protéine membranaire de la famille des protéines G.
La stimulation de certains neurones post-synaptiques entraîne la production de NO
NO (monoxyde d'azote ou oxyde nitrique) est produit grâce à la présence d'une enzyme, la NO-synthétase, qui peut être détectée par immunocytochimie. C'est par simple diffusion que NO est libéré à travers la membrane du neurone et qu'il pénètre dans le neurone receveur. Son rôle exact est inconnu.
Le plus souvent l'élément post-synaptique est un dendrite (synapses axo-dendritiques) ou un corps cellulaire (synapses axo-somatiques)
Les ramifications dendritiques de certains neurones (comme les cellules pyramidales du cortex cérébral et les cellules de Purkinje du cortex cérébelleux) sont couvertes de très nombreuses petites protrusions, appelées épines dendritiques, qui constituent autant d'éléments post-synaptiques différenciés.
4.1.1.9.2 Les synapse interneuronale électriques

Dans une synapse électrique, les membranes plasmiques des cellules pré- et postsynaptiques sont réunies par des connexons. Il s'agit de pores dynamiques à 6 sous unités ménageant des tunnels trans-cellulaires. Le flux d'ions à travers les tunnels permet à l'influx électrique de passer directement d'une cellule à l'autre.
Date de dernière mise à jour : 30/03/2018
Ajouter un commentaire