

ORGANOGENESE 3
4.1.2 L'épiblaste
Il est à la base:
1° de l'épiderme de la peau
4.1.2.1 Définition et caractéristiques ( exemple homme )
C’est un épithélium pluristratifié
- Cellules jointives à cohésion renforcée par des desmosomes
- Pas de vascularisation,les cellules sont nourries par la lymphe intersticielle.
- Innervation, terminaisons nerveuses libres intercellulaires
- Couches profondes se multipliant activement
- Couches superficielles : desquamation
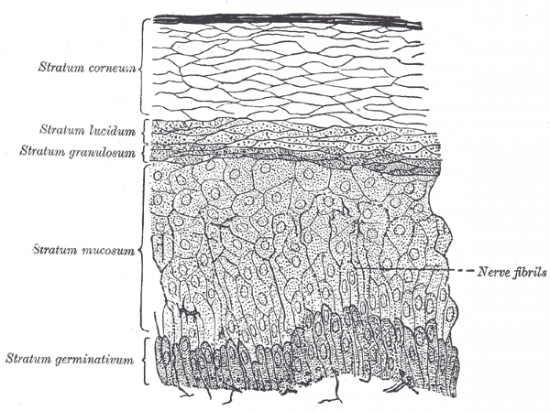
C’est la partie visible de la peau. Il a une épaisseur en moyenne de 1/10e de nm. Il est formé de cellules aplaties et juxtaposées, c’est le tissu épithélial.
L’épiderme n’est pas lisse, on observe des crêtes et des sillons. Par exemple les extrémités des doigts sont caractéristiques et différentes pour chaque personne, ce sont les empreintes digitales :
La couche germinative
La couche germinative assure par les mitoses de ses cellules le renouvellement de l'épiderme ; ses cellules, cubiques ou prismatiques, contiennent de nombreux grains de mélanine phagocytés qui permettent à l'épiderme d'assurer son rôle de protection de la lumière et qui sous-tendent le rôle de régulation de la pigmentation cutanée qu'ont les kératinocytes.
La couche à épines
Dans la couche à épines, les cellules commencent à s'aplatir, mais le noyau et les organites cytoplasmiques sont intacts, les filaments intermédiares de kératine groupés en faisceaux denses, les desmosomes normaux.
La couche granuleuse
Dans la couche granuleuse, la cellule est très aplatie, le noyau commence à dégénérer et surtout apparaissent au sein des trousseaux de filaments de kératine de nombreux grains de kératohyaline et des kératinosomes.
• La molécule constituant les grains de kératohyaline est la profilagrine, qui, dans la couche cornée, se transforme en filagrine qui la matrice du cytoplasme des cornéocytes.
• Les kératinosomes (ou corps d'Oadland) sont de petits organites ovalaires, entourés d'une membrane et présentant un aspect lamellaire ou strié périodique (d'où leur nom de granules lamellaires). Ils synthétisent dans les cellules de la couche granuleuse une substance déversée par exocytose dans les espaces intercellulaires de la couche cornée qui apparaîssent ainsi remplis d'une sorte de cément intercellulaire fait du matériel lamellaire qui était contenu dans les kératinosomes (phospholipides et glycolipides, qui se tranforment en céramides, cholestérol et acides gras libres).
La couche cornée
Enfin, dans la couche cornée, le kératinocyte (qui prend maintenant le nom de cornéocyte) est complètement aplati, le noyau et les organites cytoplasmiques ont totalement disparu et le cytoplasme est rempli de trousseaux fibrillaires formés à partir des filaments de kératine et des grains de kératohyaline. Les membranes plasmiques sont devenues très denses et épaisses et les desmosomes (qui prennent alors le nom de cornéodesmosomes) sont profondément modifiés, avec une ligne dense extra-cellulaire très épaisse ; en superficie de la couche cornée, les cornéocytes, se détachent de l'épiderme (desquamation) après la lyse du cément intercellulaire et des cornéodesmosomes (principalement sous l'action d'une enzyme sécrétée par les kératinosomes, la stéroïde-sulfatase).
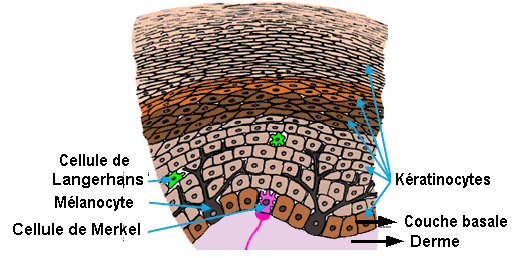
L’épiderme contient :
des mélanocytes, sécrétant la mélanine et issus des crètes neurales,
Dans l'épiderme, les mélanocytes sont situés principalement dans la couche basale. Ils ont un aspect étoilé et leurs prolongements cytoplasmiques s'insinuent entre les kératinocytes. Ils sont dépourvus de systèmes de jonction inter-cellulaire avec les cellules voisines.
En microscopie optique, les mélanocytes ne sont identifiables qu'avec des colorations argentiques ou par des techniques immunocytochimiques (HMB 45, anticorps anti-protéine S100, par exemple).
La mélanine est le pigment produit par les mélanocytes au niveau d'organites cytoplasmiques, les mélanosomes, ovoïdes mesurant 0,2 à 0,6 µm. Les mélanosomes résultent de la fusion entre des vésicules, contenant de la tyrosinase, de la dopachrome tautomérase (ou TRP2) et de la DHICA oxydase, dérivées de l'appareil de Golgi et des vésicules contenant les composants structurels des mélanosomes produites par le réticulum endoplasmique granulaire.
Ces organites contiennent un matériel fibrillaire ou lamellaire présentant une périodicité caractéristique. Quatre stades de différenciation sont classiquement décrits pour les mélanosomes.
Les stades I et II correspondent à des organites non mélanisés (parfois appelés prémélanosomes).
Les mélanosomes de stade I ont un contenu dont la structure filamentaire est encore assez mal définie. Au contraire, les mélanosomes de stade II se remplissent d'une structure interne filamenteuse. La mélanine commence à s'accumuler dans les mélanosomes de type III. Dans les mélanosomes de type IV, l'accumulation de pigments est telle que la structure interne n'y est plus visible. Les mélanosomes sont des vésicules apparentées aux lysosomes.
La biochimie de la synthèse de la mélanine n'est pas encore parfaitement connue.
On décrit deux types de pigments mélaniques : l'eumélanine qui est noir-marron et la phémélanine qui est jaune orangée. Ces pigments sont produits à partir de la L-tyrosine qui est hydroxylée en L-Dopa par la tyrosinase. La L-Dopa est oxydée en dopaquinone par cette même enzyme. La dopaquinone réagit avec la cystéine entrant dans la voie de synthèse de la phémélanine. Si la quantité de cystéine est faible dans la cellule, la dopaquinone s'oxyde spontanément en dopachrome et suit la voie de la synthèse de l'eumélanine.
La tyrosinase est l'enzyme clé de la synthèse de la mélanine.
La synthèse de la mélanine est soumise à des régulations complexes, en particulier par des hormones et des cytokines (alpha-MSH, FGF basique, HGF , insuline) ainsi que par certaines prostaglandines.
Les mélanocytes synthétisent de nombreuses cytokines (IL1alpha, IL1-bêta, IL3, IL6, TNF-alpha, GM-CSF) ; en période néo-natale, ils synthétisent également des leucotriènes dont l'action n'est pas connue.
Les mécanismes intimes du transfert des mélanosomes des mélanocytes aux kératinocytes sont incomplètement compris.
La mélanine est, en grande partie, responsable de la couleur de la peau et des phanères.
Le nombre de mélanocytes varie selon la localisation des régions cutanées chez un même individu. Ainsi, leur densité est de 2000/mm2 pour la peau de la face et de 1000/mm2 pour celle du corps. Par contre, leur nombre est sensiblement identique dans toutes les populations humaines (caucasiennes, négroïdes et mongoloïdes), la différence de couleur s'expliquant par la qualité et la quantité de pigments que ces cellules produisent. Chez les populations noires, les mélanosomes produits sont plus larges, leur contenu mélanique plus dense, ils restent isolés pendant tout le cycle de leur maturation. Au contraire, chez les populations blanches, les mélanosomes, dont les caractères s'opposent à ceux des précédents, sont associés dans des vésicules limitées par une membrane. Ainsi, chaque mélanocyte contient 5 fois plus de mélanosomes chez un sujet asiatique que chez un individu de race blanche et 8 à 10 fois plus de mélanosomes chez un sujet noir que chez un blanc.
L'exposition solaire entraîne une stimulation de la mélanogénèse et une augmentation du nombre des mélanocytes soit par différenciation de mélanoblastes quiescents, soit par division cellulaire de la cellule mature. Les mécanismes d'action des rayonnements ultra-violets (UV) ne sont pas exactement connus
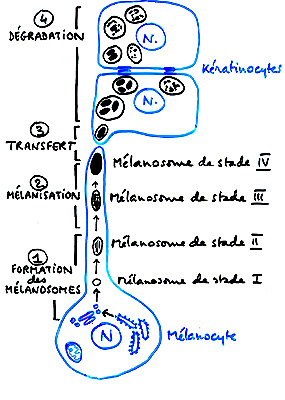
Chargement en mélanine des kératinocytes
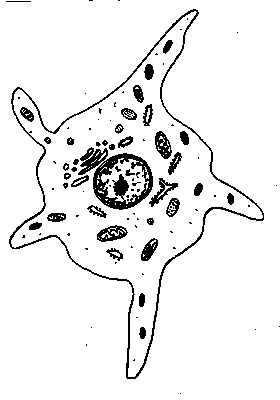
Mélanocyte
des cellules de Langerhans,
Les cellules de Langerhans font partie du groupe des cellules dendritiques. Elles dérivent des cellules souches hématopoïétiques situées dans la moelle osseuse et sont présentes dans tous les épithéliums pavimenteux stratifiés des mammifères. Elles sont en particulier dispersées entre les kératinocytes de la couche à épines de l'épiderme, la E-cadhérine jouant un rôle probablement important dans leur adhérence aux kératinocytes. La microscopie électronique permet de distinguer les cellules de Langerhans des mélanocytes, en mettant en évidence dans leur cytoplasme d'une part, l'absence de prémélanosomes et de mélanosomes et d'autre part, la présence de petits organites discoïdes pathognomoniques (granules de Birbeck).
Les cellules de Langerhans initient et propagent les réponses immunes dirigées contre les antigènes appliqués sur la peau. Elles sont capables d'ingérer des particules étrangères, y compris des micro-organismes. Après avoir capté l'antigène, les cellules de Langerhans activées quittent l'épiderme et gagnent les ganglions lymphatiques satellites où elles présentent les déterminants antigéniques aux lymphocytes T. Le GM-CSF induit la prolifération et la différenciation des précurseurs des cellules de Langerhans, ainsi que leur activation. Plusieurs marqueurs immunocytochimiques permettent de les identifier (CD1a par exemple).
de Merkel
Situées, de façon dispersée, dans la couche germinative, entre les kératinocytes basaux, au contact d'une terminaison nerveuse libre, les cellules de Merkel sont caractérisées en microscopie électronique par la présence dans leur cytoplasme de très nombreuses vésicules à centre dense entouré d'un halo clair. Les cellules de Merkel sont des cellules neuro-endocrines qui expriment des marqueurs neuronaux (chromogranine, synaptophysine, nombreux neuropeptides) et des marqueurs épithéliaux (nombreuse kératines, notamment la K20, qui, au niveau de la peau et de ses annexes, serait spécifique des cellules de Merkel). Les cellules de Merkel sont des mécanorécepteurs qui auraient également des fonctions inductives et trophiques sur les terminaisons nerveuses de l'épiderme et sur les annexes cutanées.
et des Kératynocytes.
Les molécules des filaments intermédiaires des kératinocytes sont des kératines (appelées aussi cytokératines ou alpha-kératines). On connaît actuellement une vingtaine de kératines différentes chez l'homme. Certaines kératines sont dites dures et sont spécifiquement retrouvées dans les ongles et les cheveux. D'un point de vue biochimique, on distingue les kératines de type 1 (ou kératines acides) et les kératines de type 2 (ou kératines neutres/basiques). L'assemblage des kératines se fait par hétérodimères formés entre une kératine 1 et une kératine 2. Seuls ces hétérodimères sont capables de se polymériser en filament intermédiaire. Tous les épithéliums contiennent des filaments intermédiaires de kératine (notamment K5 et K14), mais l'épiderme contient en plus plusieurs kératines différentes quasi-spécifiques de certaines couches (K1, K2, K10 et K11 sont quasi-spécifiques des couches supra-basales) et/ou de certaines régions (K9 est spécifique des paumes et des plantes).
Les kératinocytes subissent en permanence une évolution morphologique témoignant de leur kératinisation sous-tendant le rôle de barrière protectrice (mécanique et chimique) de l'épiderme. Cette évolution se fait de la profondeur vers la superficie et permet de distinguer sur une coupe d'épiderme quatre couches superposées de la profondeur vers la superficie : la couche germinative (ou basale), la couche à épines (ou spineuse), la couche granuleuse et la couche cornée (compacte, puis desquamante).

Nous verrons en anatomie comparée que les poils, ongles, cornes repris sous le vocable phanères sont des formations de l'épiblastes proches de l'épiderme.
2° Placodes :
Placodes optiques à l'origine du cristallin/cornée
L'oeuil comprend des formations ectodermiques et mésodermiques la rétine et l'iris, expansions de la vésicule cérébrale, le cristallin et la cornée sont dérivés de l'épiderme céphalique. Les enveloppes de l'œil, choroïde et sclérotique, sont mésodermiques.
Au stade bourgeon caudal, la vésicule antérieure du futur encéphale, le prosencéphale, forme latéralement deux expansions, les vésicules optiques ou placodes, qui sont réunies entre elles par la bandelette chiasmatique. Ces vésicules s'étranglent au niveau de leurs pédoncules qui deviendront les nerfs optiques.

Elle se creuse en cupules dont la concavité est tournée vers l'extérieur.Les cupules sont formées de deux feuillets accolés, l'un se différencie en rétine sensorielle tournée vers l'épiderme, il est doublé vers l'intérieur par le second qui devient le feuillet pigmentaire. La périphérie de la cupule formera l'iris. Les cupules optiques
s'appliquent contre l'ectoderme céphalique latéral qui s'épaissit en une ébauche, la placode cristallinienne à l'origine du cristallin. Cette placode s'invagine, se referme en vésicule cristallinienne et s'isole de l'épiderme. L'épiderme refermé au-dessus de la vésicule forme la cornée transparente. Les espaces entre cristallin et cupule optique sont envahis par du mésenchyme qui se différencie en corps vitré. Le globe oculaire acquiert sa rigidité grâce à la différenciation d'une enveloppe mésodermique à forte trame de collagène: la sclérotique, cartilagineuse ou porteuse d'éléments osseux dans certaines espèces (oiseaux).
s'appliquent contre l'ectoderme céphalique latéral qui s'épaissit en une ébauche, la placode cristallinienne à l'origine du cristallin. Cette placode s'invagine, se referme en vésicule cristallinienne et s'isole de l'épiderme. L'épiderme refermé au-dessus de la vésicule forme la cornée transparente. Les espaces entre cristallin et cupule optique sont envahis par du mésenchyme qui se différencie en corps vitré. Le globe oculaire acquiert sa rigidité grâce à la différenciation d'une enveloppe mésodermique à forte trame de collagène: la sclérotique, cartilagineuse ou porteuse d'éléments osseux dans certaines espèces (oiseaux).
L’ectoderme situé au-dessus du cristallin devient la cornée transparente. Elle est formée de cellules transparentes avec des espaces interstitiels réduits.
Elle ne contient pas de cellule glandulaire. Le rayon de courbure de la cornée transparente est très précis. Il est plus tard doublé d’un mésenchyme sous épithélial et d’une trame régulière de collagène élaborée par des fibroblastes. Cette différenciation peut être provoquée sur n’importe quel épiderme compétent de neurula au contact d’un cristallin ou d’un fragment de rétine. L’induction doit alors être permanente pour maintenir la transparence; après ablation du cristallin, la cornée s’opacifie.Deux autres types de placodes apparaissent :
les placodes olfactives à l'origine de l'épithélium olfactif
La morphogenese du systeme olfactif peut se diviser en trois grandes etapes : la formation de la placode olfactive, de la cavite nasale et enfin la maturation de l’epithelium olfactif embryonnaire . Ces principales etapes sont similaires chez lesmammiferes et les oiseaux, cependant leur synchronisation varie d’une espece a l’autre. (Brunjes et Frazier, 1986., Farbmann, 1992., pour revue Ayer- le Lievre et al., 1995,2000.)
La formation de la placode olfactive consiste en deux evenements principaux :la determination de la zone placodale presomptive et la neurogeneseles premieres etudes concernant les etapes precoces de la morphogenese du systeme olfactif ont ete realisees chez l’embryon d’oiseau par des experiences de xenogreffes cailles/poulets (Couly et Le Douarin, 1985 ; 1987). En effet les cellules de caille possedent une heterochromatine etroitement associee au nucleole, contrairement aux cellules de poulet dans lesquelles l’heterochromatine est repartie dans l’ensemble du noyau (LeDouarin, 1973). Une telle difference permet donc de suivre chez les chimeres caille/poulet par coloration specifique du noyau ou par immunohistochimie (anticorps anti-QCPN) le devenir des cellules de caille lors du developpement de l’embryon de poulet hote. Ainsi differentes regions de l’extremite anterieure de la plaque neurale d’embryons de poulet ont ete substituees par des regions similaires d’embryons de cailles (Couly et Le Douarin, 1985). Les etudes concernant le devenir des cellules de cailles dans de tels embryons ont permis de localiser a l’extremite anterieure de la plaque neurale le territoire presomptif de la placode olfactive. Cette localisation a pu par la suite etre precisee par l’excision de la partie externe de l’extremite anterieur du bourrelet neural aboutissant a une ebauche prosencephalique normale mais a l’absence de la placode olfactive (Ayer-Le Lievre et al., 1995).

Territoire des placodes olfactives d’un embryon de poulet (stade 3 somites) (A). Identification des bourrelets neuraux rostro-lateraux (1) et lateraux (2) de l’extremite cephalique de l’embryon (B) (modifie d’apres Couly et Le Douarin 1985).
Plus precisement, chez le poulet au stade E1 (de 0 a 3 somites), juste avant la fermeture du tube neural, le territoire presomptif des placodes olfactives se situe au niveau de la region anterieure du bourrelet neural (Couly et Le Douarin 1985, 1987, 1988. Ayer-Le Lievre et al., 1995). Au stade E2 (a partir du stade 13 de Hamburger et Hamilton), un epaississement bilateral de la zone placodale est observe. Suite aux differents mouvements morphogenetiques de cette region, les placodes se retrouvent en position ventrale, proche de la partie anterieure du tube neural, dans la future region faciale. La placode va, au stade E3 (Stade 18-19 de H&H), s’invaginer en doigt de gants pour donner naissance par la suite aux cavites nasales.

On peut encore signaler les placodes auditives et adéno-hypophysaires
4.2 Le Chordoblaste
Tigelle cylindrique axiale ( notochorde ) formant le squelette axial primaire. Elle est bien développée chez les embryons de tous les Vertébrés et persiste chez certains Vertébrés inférieurs ( Poissons condrichtyens, Cyclostomes ) . Ce chordoblaste fait partie embryologiquement d'un ensemble plus vaste appelé Chordomésoblaste
Cette tigelle se forme lors de l'évolution de la ligne primitive, chez l'homme au 18ème jours





Images tirées de : http://www.poly-prepas.com/images/files/L0%20Embryogenese.pdf
4.3 Le mésoblaste
4.3.1 Evolution générale
Le mésoblaste est constitué de deux ailes latérales qui s'étalent entre ecto et entoblasde sur toute la longueur de l'embryon, excepté les membranes pharyngiennes et cloacales.

On peut schématiser l'évolution du mésoblaste comme dans le dessin suivant :
Grossièrement, le mécanisme de l'évolution est le suivant :
1 - Clivage du mésoblaste
Feuillet pariétal : somatopleure
Feuillet viscéral : spalchnopleure
Entre les deux feuillets : coelome
2 - Etranglement
- mésoblaste dorsal : zone des somites
- mésoblaste interne : zone des néphrotomes
- mésoblaste ventral : zone des lames latérales
Rq le coelome ne subsistera qu'au niveau des lames latérales
3 - segmentation : au niveau des zones des somites et des néphrotomes

4.3.2 Evolution des somites
Une somite est un massif cellulaire creux comprenant trois parties :
1. Le dermatome
2. Le sclérotome
3. Le myotome 
Date de dernière mise à jour : 30/03/2018
Ajouter un commentaire