4.3.2.2 Myotome

4.3.2.2.1Constitution de la fibre musculaire striée et sa contraction
C'est le massif cellulaire responsable de la formation des fibres cellulaires striées constituant les parois du corps.
Le caractère strié : résultat de l'arrangement de 2 types de myofilaments formés de protéines contractiles :
- Les myofilaments épais : assemblage de myosine. La myosine est une protéine présentant une queue et une tête assez massive
Les queues ont une action purement mécanique et s'agglutinant entre elles ( filaments épais ) tandis que les têtes restent libres ( possibilités de mouvement appréciables ) . C'est dans la tête que se trouvent les structures vraiment intéressantes intervenant dans la transformation de l'énergie chimique en énergie mécanique
Elle porte :
- un groupement enzymatique capable de fixer et d'hydrolyser l'ATP.
- Un site d'accrochage de l'actine, par l'intermédiaire de la quelle s'effectue la transmission de la force mécanique.
- Myofilaments fins : assemblage de trois protéines :
- Actine : la principale. C'est une protéine globuleuse. Elle ne possède pas de site d'activité de l'ATPase mais chaque molécule globulaire est solidement liée à une molécule d'ATP qu'elle renferme. Ces molécules se polymérisent pour former un cordon à double brin ( polymérisation réalisée grâce à l'utilisation de l'ATP )
- Tropomyosine : elle est formée de longue tiges constituées chacune d'une double hélice d'une longeur approximativement égale à une queue de molécule de myosine.
- Troponine : protéine composée de trois sous unités à grande affinité pour le calcium, c'est la protéine la moins abondante dans le muscle. Une de ses sous unités constitue le dispositif par l'intermédiaire duquel le calcium contrôle la contraction en activant et désactivant l'actine
Les muscles sont constitués par l'association de fibres musculaires.Celles-ci sont en fait des syncitiums de cellules fusionnées (figure 2 ) . Elles se présentent comme des fuseaux longs et fins d'un diamètre de 10 à 100 µm et d'une longueur pouvant aller jusqu'à 30 cm. Chaque fibre peut contenir plusieurs centaines de noyaux et est entourée d'une robuste membrane (sarcolemme) formée par l'association de la membrane plasmique et d'une épaisse lame basale
riche en glycoprotéines et en fibres conjonctives, l'ensemble constituant une architecture à haute résistance mécanique.
Figure 1
La presque totalité de l'intérieur des fibres est remplie de faisceaux d'éléments contractiles: les myofibrilles, longs cylindres de 1 à 2 µm de
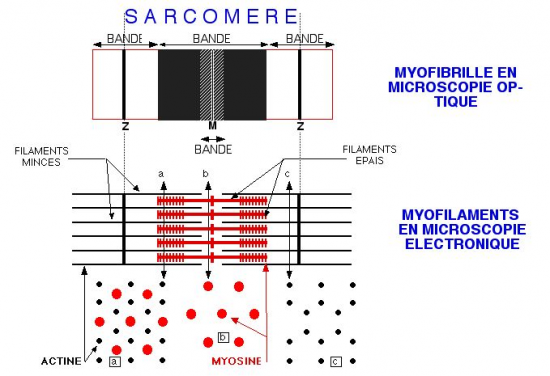
diamètre présentant une striation transversale périodique caractéristique qui est à l'origine de la dénomination de muscle strié (figures 1 et 2 ). Cette striation est due à un arrangement particulier des deux types de filaments contractiles: la myosine et l'actine (figure 2). Ces filaments sont organisés dans des éléments répétitifs appelés sarcomères limités par des membranes dites Z. Chaque sarcomère présente une alternance de filaments épais (myosine) et de filaments plus minces (actine), ces derniers étant attachés aux membranes Z. L'empilement de ces filaments rend compte de l'organisation en bandes A (foncées, anisotropes) et I (claires, isotropes) des cytologistes (figure 1).
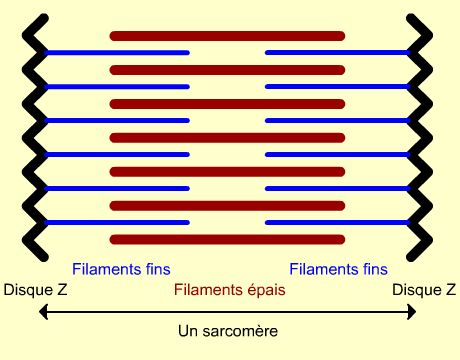 Sarcomère
SarcomèreCoupe transversale d'une myofibrille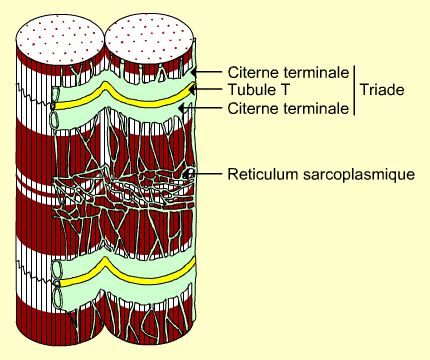
L'espace laissé libre entre les myofibrilles est occupé par du cytoplasme (appelé ici sarcoplasme) dans lequel on retrouve les organites et les inclusions classiques: reticulum endoplasmique (ici sarcoplasmique), mitochondries, noyaux, gouttelettes lipidiques, etc. On y trouvera également un pigment respiratoire particulier: la myoglobine (voir plus loin). Le reticulum sarcoplasmique est organisé de façon très particulière, entourant les myofibrilles (figures 1 et 2). Son organisation est par ailleurs en rapport étroit avec la disposition périodique des sarcomères: à intervalles réguliers, il présente des manchons dilatés formant des structures dites citernes terminales qui jouxtent un réseau d'invagination de tubules dits transverses. Un tubule et les citernes l'entourant de part et d'autre forment ce qu'il est convenu d'appeler une triade. A ce niveau, la membrane du tubule et celle du reticulum forment des ensembles histologiquement assez confus (figure 2) et considérés comme les zones de couplage électrophysiologique au niveau desquels la contraction est initiée à partir du message nerveux.On trouve donc un tubule transverse en étroite association avec deux citernes terminales, formant ce que l'on appelle une triade (Figure 2). Cette organisation joue un rôle important dans le couplage excitation-contraction 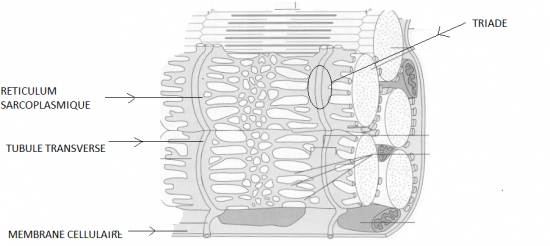
les nombreuses myofibrilles disposées parallèlement remplissent l'essentiel du volume cellulaire, ne laissant que peu de place au sarcoplasme (cytoplasme d'une cellule musculaire). De façon remarquable, les sarcomères des différentes myofibrilles sont situés au même niveau selon l'axe longitudinal. Ainsi, vue en microscopie photonique, une cellule musculaire apparait régulièrement zébrée transversalement sur toute sa longueur, d'où le nom de muscle strié Notons que les bandes sombres ne correspondent pas aux disques Z mais aux filaments épais et les bandes claires correspondent aux zones ne comportant que des filaments fin (donc à cheval sur deux sarcomères). Vue en microscopie électronique, on peut préciser cette striation au niveau ultrastructural avec la visualisation des disques Z, bandes A (zone des filaments épais, bandes sombres visibles en microscopie photonique), bandes I (zone ne présentant que des filaments fins, bandes claires visibles en microscopie photonique), zone H (zone centrale du sarcomère sans filaments fins) et strie M (zone d'attachement des myosines têtes-bèches).
La contraction des fibres musculaires est due aux interactions des filaments d'actine et de myosine et à leur glissement les uns par rapport aux autres.Ce mécanisme fait intervenir différentes protéines outre l'actine et la myosine. Il nécessite par ailleurs du Ca2+ et de l'ATP. Il s'agit d'un processus complexe dont certains aspects restent encore obscurs à l'heure actuelle. Il ne sera pas considéré en détail ici, faisant l'objet d'une section de la rubrique physiologie animale. Nous n'en donnerons donc qu'unrésumé très bref Lors de la contraction d'un sarcomère, la longueur des deux catégories de filaments reste invariable: le mouvement provient en fait du glissement des filaments d'actine le long des filaments de myosine.
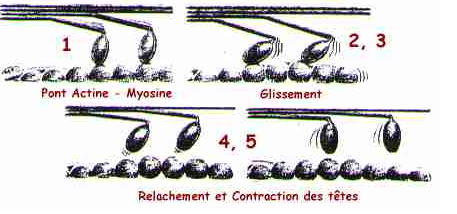
Les filaments de myosine sont des assemblages protéiques polymériques formés de parties a - hélicoïdales et de têtes globulaires. Les têtes sont orientées vers l'extérieur de l'assemblage,sortant de celui-ci en formant une spirale. Le nombre de têtes par tour de spire de même que le nombre de molécules de myosine constituant la fibre sont variables. Les filaments de myosine seront donc plus ou moins long et plus ou moins épais selon les types de muscles et les espèces considérés. Les têtes sont par ailleurs mobiles et leur angulation par rapport à l'axe du filament peut varier. C'est ce mouvement qui est en fait à la base du mouvement musculaire.
Les filaments d'actine sont des polymères fibreux formés de l'association de monomères d'actine globulaire. Ils entourent, en nombres variables suivant les espèces et les types de muscles, les filaments de myosine. Chez les vertébrés, on compte en général 6 actines par myosine; ce nombre peut aller jusqu'à 12 dans certains muscles d'arthropodes. Les filaments d'actine présentent à intervalles réguliers des sites de fixation pour les têtes de myosine. Dans le muscle au repos, ces sites sont normalement masqués par des associations de protéines régulatrices (tropomyosine et troponines, cfr. physiologie animale).
La contraction démarre avec l'arrivée sur le muscle d'ondes de dépolarisation provenant de signaux nerveux. L'organisation des jonctions neuromusculaires est variable. Certains muscles n'en présentent qu'une par fibre, d'autres une grande quantité . Les ondes de dépolarisation se propagent le long des membranes
sarcoplasmiques et pénètrent à l'intérieur des fibres musculaires, jusqu'au voisinage du reticulum sarcoplasmique par les invaginations du système de
tubules transverses (figure2). Elles induisent à ces endroits des entrées de Ca2+ et des libérations de modulateurs qui vont entraîner un relarguage massif de Ca2+ des citernes du reticulum sarcoplasmique entourant chaque faisceau de myofibrilles (voir physiologie animale). L'importante augmentation de Ca2+ dans le sarcoplasme provoque alors des changements d'organisation des protéines régulatrices des filaments d'actine ce qui rend accessible leurs sites de fixation aux têtes globulaires des filaments de myosine.
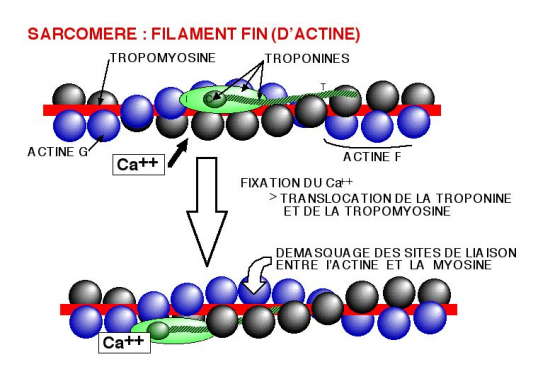
L'Actine F est associé à deux filaments de tropomyosine qui se mettent dans les sillons de l'hélice.
Au repos, la tropomyosine masque les sites d’interaction actine-myosine.
Par ailleurs, et lorsque les concentrations de calcium intracellulaire sont faibles, une autre molécule globulaire,
la troponine, est associée à la fois à l'actine F et à la tropomyosine.
La troponine est une molécule Calcium dépendante. La fixation de Calcium provoque un changement
de conformation qui rend accessible l'actine aux têtes de myosines.
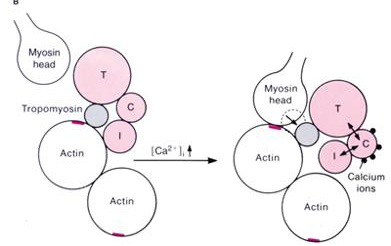
L'interaction actine - myosine et la présence de Ca2+activent alors une ATPase située dans les têtes de myosine: l'hydrolyse d'ATP qui s'ensuit produit l'énergie nécessaire à leur mouvement . Après s'être fixées aux filaments d'actine, les têtes de la myosine activées pivotent ainsi d'environ 10 nm. Le mécanisme intime de ce mouvement n'est pas encore compris. Dans un modèle proposé en 1971, Huxley et Simmons (Nature, 233, 533) considèrent sur les têtes de myosine une série de quatre sites d'accrochage à l'actine, chacun étant activé successivement et présentant une affinité pour son site supérieure à celle
du précédent.
4.3.2.2.2 organisation du muscle et classification
la masse musculaire est structurée et emballée dans du tissus conjonctif principalement à base de fibres de collagène :
- autour de la fibre: lame appelée endomysium
- Autour du muscle: lame appelée épimysium
- les fibres musculaires sont reliées par bottes à l'aide du périmysium 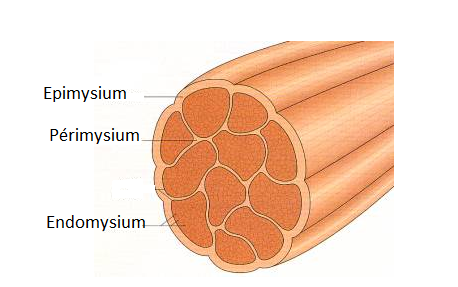 Une première classification des muscles se base sur leur fonction :
Une première classification des muscles se base sur leur fonction :
1) Les muscles somatiques qui sont axiaux ou appendiculaires ( squelettiques ) ils sont issus du myotome et sont commandés par la volonté, ils sont dits volontaires
2) Les muscles viscéraux qui sont viscéraux stricts ou cardiaques ils dérivent de la splanchnopleure et sont involontaires.
Les muscles somatiques sont striés comme le muscle cardiaque, les muscles viscéraux stricts sont dits "lisses" ( peu de fibres peu organisées ). Tous les muscles sont d'origine mésoblastique.
4.3.2.2.3 . Les différences muscle squelettique - muscle cardiaque
L'organisation ultrastructurale du muscle strié squelettique et cardiaque n'est pas très différente. Néanmoins, on peut distinguer quelques spécificités.
Première différence : les cellules musculaires squelettiques sont des cellules syncitiales polynucléées pouvant atteindre plusieurs centimètres de long. Au contraire, les cellules musculaires cardiaques ne sont pas issues de la fusion de plusieurs cellules souches musculaires et sont donc uninucléées (Figure 4). Leur taille est donc généralement sans commune mesure avec celle de leurs homologues squelettiques. L'unité mécanique est obtenue par un attachement des cellules disposées selon l'axe longitudinal par l'intermédiaire de disques intercalaires riches en desmosomes (voir figure 4). Par ailleurs, les cytoplasmes des cellules cardiaques sont en communication directe via de nombreuses jonctions communicantes.
|
|
Figure 4 : Vue en microscopie photonique de cellules
musculaires cardiaques de singe. Les cellules musculaires ont globalement
une forme rectangulaire, le noyau étant visible chez plusieurs d'entre
elles. Les astérisques correspondent aux disques intercalaires (zones
d'attachement entre deux cellules). Source Biodidac (http://biodidac.bio.uottawa.ca/) |
Deuxième différence : si l'organisation structurale du matériel contractile est globalement similaire dans les deux cas, les cellules cardiaques
sont souvent ramifiées et les myofibrilles sont plus courtes que dans les cellules musculaires squelettiques.
Enfin, troisième différence : le reticulum sarcoplasmique est considérablement moins développé dans les cellules musculaires cardiaques par rapport à la situation observée dans les cellules musculaires squelettiques, cette observation laissant entrevoir des différences fonctionnelles.
Il existe principalement trois différences fonctionnelles entre muscle squelettique et muscle cardiaque.
La première est extérieure aux cellules musculaires elles-mêmes : la commande nerveuse des muscles squelettiques est assurée par les nerfs moteurs, ce qui correspond à une commande volontaire (hors mouvement réflexe). A l'inverse, la régulation nerveuse du muscle cardiaque est assurée par les nerfs issus du système sympathique cardioaccélérateur et parasympathique cardiomodérateur (nerf vague ou pneumogastrique X) du système nerveux autonome, ce qui correspond à une commande involontaire. Sans oublier que les cellules cardiaques se contractent rythmiquement en absence de toute influence nerveuse sous l'impulsion des cellules pace-maker.
La seconde correspond à une différence dans le couplage excitation-contraction Dans le muscle squelettique, l'augmentation de la concentration intracellulaire en calcium (responsable de la contraction) est non seulement le résultat d'un influx de calcium extracellulaire mais surtout d'un efflux de calcium provenant du reticulum sarcoplasmique. Dans le muscle cardiaque, cette augmentation de calcium intracellulaire est essentiellement due à un un influx de calcium extracellulaire, le reticulum sarcoplasmique étant moins développé.
Enfin troisièmement, le muscle cardiaque n'est pas tétanisable. La tétanie se caractérise par un plateau de contraction de puissance maximum par suite d'une stimulation à une fréquence ne permettant pas au muscle de se relâcher entre deux contractions. Or, la période réfractaire absolue (période durant laquelle une cellule excitable qui vient d'être stimulée n'est pas en mesure de répondre à une nouvelle stimulation) est beaucoup plus importante pour les cellules cardiaques que pour les cellules musculaires squelettiques. La cellule musculaire cardiaque a le temps de se relâcher avant d'être en mesure d'être à nouveau stimulée. Il est donc impossible d'obtenir une sommation des contractions. Il en résulte que le muscle cardiaque n'est pas tétanisable au contraire du muscle squelettique.Notons également qu'entre les cellules musculaires cardiaques se trouvent des synapses électriques permettant une grande rapidité dans l'apparition du potentiel d'action musculaire.
A la lumière de la comparaison entre muscle squelettique et muscle cardiaque, il apparaît que leurs points communs permettent de les classer dans une même catégorie, celle des muscles striés, mais que leurs différences, aussi bien structurales que fonctionnelles, imposent de les séparer en deux sous-catégories.
Il est ainsi classique de les différencier en parlant de muscles squelettiques volontaires et de muscle cardiaque involontaire. Pourtant, si elle est essentielle,
cette différence n'est pas le fait des cellules musculaires elles-mêmes mais des mécanismes de commande (présence ou non de cellules pace-maker
et origine de l'innervation afférente). Il est donc important que cette différence physiologique ne masque pas les autres différences qui, elles, sont directement liées à la structure et au fonctionnement des cellules musculaires.
4.3.2.2.4 Les muscles lisses,
Contrairement aux muscles striés, se composent de cellules musculaires fusiformes uninuclées.Ces fibres musculaires non-striées se contractent lentement et involontairement (activité contractile spontanée). Ils sont sous le contrôle du système nerveux autonome parasympathique dont le neurotransmetteur est l'acétylcholine Chaque cellule, qui a des extrémités pointues et bifides, ne possède qu'un seul noyau bien visible, en général central et allongé, et situé
dans la partie la plus large de la cellule. Elles sont délimitées par le sarcolemme, combinaison de la membrane plasmique et d'une lame basale externe. Le cytoplasme (appelé sarcoplasme pour les cellules musculaires) apparaît assez homogène. Cependant, on distingue à l'intérieur des myofibrilles (=myofilaments) orientés selon le grand axe de la cellule. Les myofilaments sont composés des deux protéines contractiles : les fins d'actine et les épais de myosine. Des myofilaments fins entourent un myofilament épais, et la contraction a lieu en présence d'ATP. Le réticulum sarcoplasmique n'a pas la régularité de celui des fibres striées, et ne sert pas de réservoir de calcium.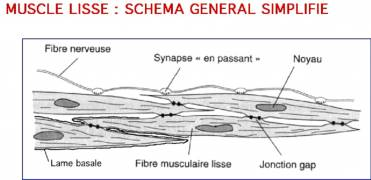






 Sarcomère
Sarcomère








 Une première classification des muscles se base sur leur fonction :
Une première classification des muscles se base sur leur fonction : 
