

HISTOIRE DES VERTEBRES 7
3.2.7 . La classe des mammifères.
3.2.7.1 Introduction
Ce sont des homéothermes dont les phanères épidermiques forment des poils. Leur peau possède des glandes cutanées et de nombreuses glandes sébacées sont associées aux poils où elles sécrètent le sébum pour que le poil luise.
Les glandes sudoripares : elles sécrètent un liquide riche en azote et en ions : elles contribuent à une excrétion de vapeur qui permet un dégagement de calories pour assurer une partie de la régulation thermique.
Les glandes mammaires : elles produisent une sécrétion (le lait) pour la nutrition du jeune.
Les mammifères sont apparus en même temps que les dinosaures. Ils ont été obligés d’attendre que ces reptiles s’éteignent pour pouvoir se développer.
On a trois lignées différentes de mammifères qui se sont maintenues jusqu’à nos jours.
a\ L’ordre des Monotrèmes (protothériens, cloaque commun).
C’est un petit groupe qui vit en Nouvelle Zélande. Les ornithorynques sont des mammifères sans dent et avec un bec corné. Ils sont plus ou moins aquatiques.
Les Echidnés sont terrestres, primitifs, reptiliens, pondent des œufs et leur homéothermie est imparfaite (entre 25 et 36°C). Les canaux des glandes mammaires ne se regroupent pas : chacun a son orifice. Le lait coule le long des poils et le jeune lèche ce lait sur le corps maternel.
Pour les prochains groupes, le cloaque est cloisonné.
b\ L’ordre des marsupiaux (Métathériens).
Les canaux galactophores se regroupent en mamelles. La vie intra utérine est courte. Le placenta est incomplet et ne permet pas de barrière immunologique : l’embryon est expulsé. Il va remonter sur la mère pour gagner la poche marsupiale qui est tendue par deux os dits « pelviens ». Là, le jeune saisie une mamelle reste dans cette poche jusqu’au terme.
On trouve les marsupiaux surtout en Australie et en Amérique du sud. Avant, on les trouvait partout.
c\ L’ordre des placentaires (les Euthériens).
C’est un groupe qui regroupe autour de 4000 espèces.
3.2.7.2 Origine et évolution
A) Quelles sont les caractéristiques des Mammifères
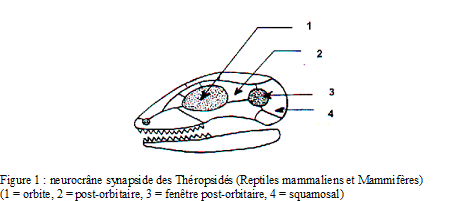
Neurocrâne
o neurocrâne synapside (une ouverture temporale unique),
o 2 condyles occipitaux permettent l’articulation à la première vertèbre cervicale (atlas),
o fort volume de la boîte crânienne en rapport avec le développement de l’encéphale : à tailles corporelles égales, le cerveau d’un mammifère est plus volumineux que celui d’un reptile, en particulier les hémisphères cérébraux et le cervelet,
o présence d’un palais osseux séparant la cavité buccale en un étage olfactivo-respiratoire et un étage masticateur et autorisant respiration et mastication simultanées.
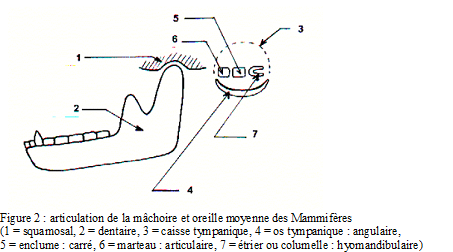
Splanchnocrâne
o mâchoires puissantes,
o constitution de la mâchoire inférieure (mandibule) par 1 seul os (dentaire),
o articulation de la mâchoire inférieure entre squamosal (appelé aussi temporal) et dentaire,
o oreille moyenne à trois osselets (marteau, enclume et étrier).
Dentition
o thécodonte (dents implantées dans des alvéoles de la mâchoire),
o hétérodonte (dents différenciées et spécialisées en incisives, canines, prémolaires et molaires),
o diphyodontie (2 dentitions successives : la dentition définitive remplace la dentition lactée).
Colonne vertébrale à vertèbres différenciées (présence de côtes et d’un diaphragme)
Membres parasagittaux sous le corps (sauf chez l’Ornithorhynque chez lequel les membres sont transversaux)
Tégument et formations annexes
o pelage associé au maintien de la température corporelle à un niveau élevé,
o glandes lactéales : chez tous existe une nutrition lactée du jeune
o des soins sont portés aux jeunes.
Physiologie
o ventilation pulmonaire efficace (existence d’une cage thoracique et d’un diaphragme) autorisant un métabolisme élevé lié à l’endothermie,
o endothermie (le maintien de la température corporelle élevée est assuré par le pelage limitant la déperdition de chaleur, le métabolisme élevé et la physiologie de thermorégulation), besoins métaboliques élevés couverts par une alimentation abondante, amniotes.
Viviparité (sauf chez les Monotrèmes qui sont ovipares)
B) Les Mammifères actuels : aperçu de la diversité
* Protothériens ou Monotrèmes (Ornithorhynque, Echidné d’Australie, Tasmanie et Nouvelle Zélande)
* Thériens (ou Mammifères modernes) subdivisés en :
- Métathériens ou Marsupiaux (Australie, Tasmanie, Nouvelle Guinée et Amériques) à vie intra-utérine brève et poche marsupiale
- Euthériens ou Placentaires à développement embryonnaire intra-utérin long (gestation)
NB : les Théropsidés sont les vertébrés à affinités mammaliennes; ils sont tous synapsides et regroupent les Reptiles mammaliens (exclusivement fossiles) et les Mammifères.
C) Identification paléontologique d’un Mammifère
Elle n’est possible qu’à partir de l’étude des éléments durs minéralisés fossilisables (os, dents).
De nombreux mammifères fossiles ne sont connus que par leurs dents, un fragment de mâchoire ou mieux encore leur crâne. Les marsupiaux se distinguent des placentaires par la formule dentaire, la constitution du plancher crânien, la mandibule à processus angulaire et l’épipubis à rôle de soutien de la poche marsupiale.
I – L’arbre phylétique des Mammifères
A) Origine reptilienne des Mammifères : les reptiles mammaliens
1) Les Pélycosauriens. Cet ordre apparu au Carbonifère supérieur (- 290 Ma) s’éteint au Permien moyen et ce sont des dépôts continentaux du Permien inférieur de Russie et d’Amérique du Nord –Texas– qui offrent les meilleurs gisements.
Physionomie générale : aspect de grands lézards à membres courts (1,5 à 3 m selon les espèces)
Des caractères reptiliens indiscutables
articulation de la mâchoire inférieure au crâne de type reptile
membres transversaux
Des caractères mammaliens :
Crâne synapside (caractère commun aux Théropsidés : Mammifères et Reptiles mammaliens)
ébauche de palais osseux
membres antérieurs à autopode dirigé vers l’avant
denture thécodonte et hétérodonte (canines nettes bien différenciées)
Diversité : grand nombre d’espèces (Varanosaurus, Edaphosaurus, Ophiacodon, Sphenacodon) avec des espèces herbivores et des espèces carnivores ; chez les espèces carnivores, l’hétérodontie est plus marquée (canines très nettes) que chez les herbivores ;
Exemple : Dimetrodon (jusqu’à 3 m de long) est une espèce carnivore du Permien inférieur du Texas dotée d’un grand voile cutané dorsal soutenu par des apophyses vertébrales très allongées (ce voile cutané jouait probablement un rôle thermorégulateur : absorption ou évacuation de chaleur selon les besoins de l’animal ?).
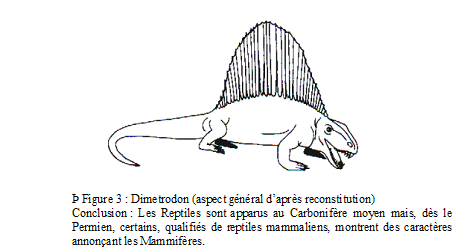
Les Pélycosauriens s’éteignent au Permien moyen supplantés par leurs descendants : les Thérapsidés.
2) Les Thérapsidés. Cet ordre apparu à la base du Permien supérieur est plus évolué que celui des Pélycosauriens ; il est le groupe de Vertébrés dominant du Permien supérieur au Trias ; ses derniers représentants s’éteignent au Jurassique moyen, il y a 160 Ma. Il présente une très large distribution à la surface du globe mais les principaux gisements sont situés dans des dépôts continentaux du Permien supérieur et du Trias d’Afrique du Sud (Karoo), du Niger, de Russie (Oural) et des Amériques.
Ils montrent une nette évolution vers la forme mammalienne
grande extension du dentaire aux dépens des autres os de la mandibule,
articulation de la mâchoire inférieure au crâne intermédiaire entre le type reptile et le type mammifère,
denture thécodonte nettement hétérodonte,
muscles masticateurs puissants révélés par leurs zones d’insertion,
membres parasagittaux relevés et rapprochés du corps,
corps élevé au dessus du sol.

Diversité : au sein de l’ordre des Thérapsidés peuvent être distingués plusieurs sous-ordres dont les Anomodontes herbivores et les Thériodontes carnivores.
Les Anomodontes ne possèdent pas de palais osseux véritable (il est absent ou rudimentaire) ; on y distingue les Dinocéphales (ex : genre Moschops de la taille d’un rhinocéros) et les Dicynodontes dont la taille va de 0,3 à plusieurs mètres (ex : genre Lystrosaurus à dentition réduite aux canines et présence d’un bec corné). Les Anomodontes sont très spécialisés et ils s’éteignent sans postérité ; ils ne sont pas les ancêtres des Mammifères.
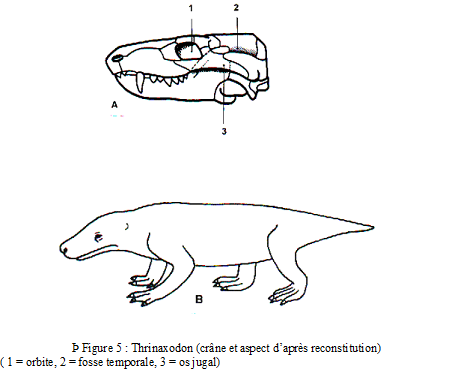
Les Thériodontes regroupent les Gorgonopsiens, les Thérocéphales et surtout les très évolués Cynodontes (tels Thrinaxodon et Cynognathus) qui apparaissent comme les précurseurs directs des Mammifères par les caractères suivants :
Essai de reconstitution
Cynognathus (longueur # 1m) présente un aspect voisin de celui du blaireau mais avec une tête plus large et une queue plus courte. Doté de puissantes mâchoires à denture de Mammifères, il se nourrissait en fouillant le sol à la recherche de petits animaux et de végétaux.
Caractères mammaliens indiscutables des Cynodontes
vaste volume de la boîte crânienne,
2 condyles occipitaux,
existence d’une véritable voûte palatine séparant la cavité buccale en étage respiratoire et étage alimentaire,
articulation de la mâchoire inférieure entre dentaire et squamosal chez les plus évolués mais certaines espèces (Diarthrognathus du Trias supérieur) montrent la présence des deux types d’articulation (reptilienne entre carré et articulaire et mammalienne entre dentaire et squamosal),
très nette hétérodontie (dents post-canines dotées de cuspides émoussées autorisant le broyage des os et la mastication des chairs) comparable à celle des Mammifères,
différenciation de la colonne vertébrale en vertèbres dorsales et vertèbres lombaires.
Caractères mammaliens hypothétiques ou spéculatifs des Cynodontes
respiration et mastication simultanées,
fort développement du cervelet (siège des commandes motrices) en rapport avec une activité élevée,
existence d’un diaphragme autorisant une respiration plus efficace et donc un métabolisme élevé,
présence de dépressions dans les os maxillaires comme chez les mammifères munis de vibrisses (étaient-ils pourvus de poils ? étaient-ils homéothermes ?),
la denture des jeunes, plus simple que celle des adultes, est dépourvue de dents masticatrices (diphyodontie ? jeunes nourris par les parents ? présence de mamelles ?).
Important : de nombreux reptiles et reptiles mammaliens se sont éteints lors de la crise biologique de la fin du Trias ; ils ont laissé le champ libre aux dinosaures du Jurassique et du Crétacé. Plusieurs causes peuvent être invoquées pour cette crise :
o un impact d’origine extraterrestre (ou plusieurs?) correspondant une chute d’astéroïde (au Québec, le cratère du Manicouagan d’âge Norien et d’environ 70 kilomètres de diamètre en serait un témoignage),
o des éruptions volcaniques avec émissions de gaz et de cendres modifiant et/ou assombrissant l’atmosphère (épanchements des trapps basaltiques du Karoo en Afrique du Sud et du New-Jersey sur la côte Est des USA datés respectivement de –220 MA et – 205 MA),
o un réchauffement climatique d’âge Norien avec dépôts d’évaporites (évaporites continentales du Trias supérieur) et baisse de la salinité des eaux marines,
o une régression marine mondiale généralisée depuis le Trias supérieur suivie d’une transgression (peut-être dysoxique ?) dès le Jurassique inférieur.
B) Les premiers Mammifères véritables au Mésozoïque
Les premiers mammifères indiscutables apparaissent à la fin du Trias (Rhétien vers - 200 Ma) en même temps que les plus anciens dinosaures et sont certainement issus des Thérapsidés cynodontes. Nous ne savons pas si ces premiers mammifères étaient vivipares et s’ils allaitaient leurs jeunes mais les fossiles indiquent qu’ils avaient la démarche, les mâchoires et la dentition des mammifères actuels. Ils étaient probablement endothermes.
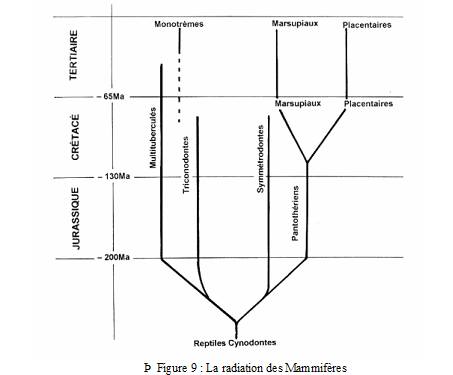
Plusieurs lignées peuvent être reconnues mais la structure de la paroi latérale du neurocrâne, l’organisation de la ceinture scapulaire permettent de distinguer deux ensembles :
• les Protothériens représentés par les Triconodontes, Docodontes et Multituberculés
• les Thériens représentés par les Symmétrodontes et Pantothériens.
1) Les lignées sans postérité
Ce sont les groupes dominants de la faune mammifère du Mésozoïque ; animaux de la taille d’une musaraigne pour les plus petits, quelques espèces atteignent la taille du lapin voire du blaireau.
Les Triconodontes apparaissent au Trias supérieur vers -200 Ma et s’éteignent avant la fin du Crétacé vers -75 Ma. Ce sont des animaux de petite taille (15 cm pour un poids estimé de 20 à 30 g) à aspect de musaraigne. Leur répartition très large est pangéenne (Morganucodon du Rhétien du Pays de Galles, Mégazostrodon du Jurassique inférieur d’Afrique du Sud et Sinoconodon du Jurassique inférieur du Yunan chinois).
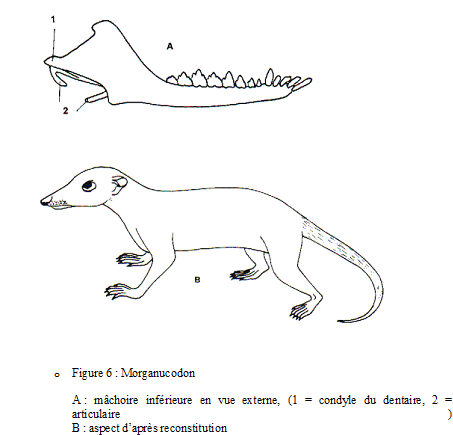
Ils présentent d’indiscutables caractères mammaliens :
o mâchoire inférieure constituée du seul os dentaire,
o articulation de la mâchoire inférieure au crâne de type mammifère entre dentaire et squamosal,
o vaste volume crânien (cerveau plus développé que celui des Cynodontes),
o denture hétérodonte avec I, C, PM et M (molaires à 3 cuspides alignées) et dentitions successives,
o régime insectivore déduit de l’aspect des PM et M à nombreux tubercules,
o pelage et endothermie sont déduits de la petite taille (un animal de petite taille présente un fort rapport surface / volume donc une forte déperdition thermique impérativement compensée par un métabolisme élevé, donc une alimentation abondante or tout ceci suppose une activité soutenue incompatible avec des températures corporelles basses et fluctuantes),
o grandes facultés sensorielles attestées par le grand volume des capsules olfactives, optiques et otiques,
o mœurs nocturnes (?) : ils échappent ainsi par leur petite taille aux dinosaures prédateurs et occupent de nombreuses niches écologiques.
Chez les Triconodontes, la denture est très bien connue grâce aux nombreux crânes fournis par le gisement du Yunan et elle peut être suivie de façon complète du jeune à l’adulte. Sinoconodon montre une denture à renouvellement multiple fonctionnelle dès le plus jeune âge comme chez les reptiles alors que Morganucodon présente un seul renouvellement de la denture comme chez les mammifères allaitant (Marsupiaux et Placentaires). Ceci est un argument en faveur de l’allaitement chez le seul Morganucodon et en faveur de l’acquisition de l’allaitement par certaines des espèces triconodontes.
Les Multituberculés constituent un groupe d’extension mondiale apparu au Jurassique supérieur. Dominant la faune mammalienne au Crétacé où ils représentent à eux seuls le tiers de la diversité, ils sont touchés par la crise Crétacé–Tertiaire (la moitié des Multituberculés disparaît alors que tous les Placentaires survivent) et ils s’éteignent fin Eocène (-35 Ma) à l’occasion de la " Grande coupure ".Ce sont des animaux à régime phytophage de la taille du mulot caractérisés par des incisives en lame coupante, l’absence de canines et des molaires à nombreux tubercules disposés en rangées parallèles.

Ils tiennent probablement une place écologique comparable à celle des Rongeurs actuels. Certaines espèces à vastes orbites sont considérées comme diurnes alors que d’autres seraient nocturnes (orbites réduites mais capsules olfactives développées). La présence d’un pelage est déduite des poils découverts dans les coprolithes de leurs prédateurs.
Les Symmétrodontes s’éteignent avant la fin du Crétacé (-75 Ma). Petits animaux de la taille d’une musaraigne, ils sont connus (Kuehneotherium) par des mandibules et des dents (molaires à tubercules disposés en triangle).
Les Docodontes s’éteignent fin du Jurassique (-100 Ma) ; leurs post-canines possèdent des cuspides disposées en triangle (molaire tribosphénique) comme chez les Thériens.
2) Les lignées conduisant aux Mammifères actuels
Les Pantothériens apparaissent au Jurassique moyen et conduisent aux Thériens (Marsupiaux et Placentaires actuels) dont les premiers représentants sont datés du sommet du Crétacé inférieur de Mongolie (Albien à – 105 Ma).

Animaux de petite taille, sans doute arboricoles et nocturnes, ils forment un groupe diversifié (on y reconnaît plusieurs familles) caractérisé par des molaires larges dont les cuspides en vis à vis s’emboîtent (système de pilon et mortier). La mandibule est dotée d’un processus angulaire permettant l’insertion du masséter ; la mastication comporte une composante transversale et le régime alimentaire est partiellement carnivore. De bons gisements sont connus dans les terrains du Jurassique moyen et supérieur d’Europe (Angleterre) et d’Amérique du Nord (Wyoming).
3) La dichotomie Marsupiaux – Placentaires
C’est au cours du Crétacé (vers – 100 Ma) que l’isolement géographique de deux groupes de Pantothériens conduit aux premiers Marsupiaux apparus en Amérique du Nord (ou en Asie ?) et aux premiers Placentaires apparus en Asie (Mongolie). Marsupiaux et Placentaires apparaissent donc comme deux groupes frères dérivant d’un groupe ancestral commun : les Placentaires ne sont pas les descendants des Marsupiaux.
En résumé, le passage des Reptiles aux Mammifères s’effectue par des modifications profondes portant sur de nombreux caractères : on peut donc parler ici de transition évolutive.
4) L’origine géographique des Monotrèmes et des Marsupiaux
L’origine des Monotrèmes actuels est méconnue. Leurs plus anciens fossiles datent du Crétacé inférieur d’Australie (Steropodon à – 115 Ma) où le groupe est sans doute apparu puis s’est diversifié avant d’atteindre Madagascar et l’Amérique du Sud. Une parenté avec les Multituberculés est possible.
* L’origine géographique des Marsupiaux est encore l’objet de controverses.
Les plus anciens marsupiaux connus sont datés du Crétacé moyen du Texas (Holoclemensia à – 100 Ma, Alphadon à – 80 Ma), ce qui indiquerait une origine nord-américaine. Cependant, même pour les tenants de cette origine américaine, la patrie des Marsupiaux change souvent en fonction des découvertes paléontologiques les plus récentes et en quelques décennies, elle fut alternativement située en Amérique centrale, au Nord de l’Amérique du Sud ou au Sud de l’Amérique du Nord.
Pour d’autres, la patrie des Marsupiaux se trouverait en Asie (Mongolie) car il y existe une faune marsupiale très diversifiée dès le Crétacé supérieur (famille des Deltathéridés : Asiatherium, Deltatheridium à – 75 Ma Campanien).
C) Les Mammifères du Cénozoïque
Les mammifères modernes (Marsupiaux et Placentaires) sont très peu diversifiés pendant la majeure partie du Mésozoïque. Ils ne sont identifiés qu’à la fin du Crétacé inférieur mais, dès le début de l’ère tertiaire, le phylum se diversifie, supplante celui des Reptiles et conquiert la totalité des terres émergées.
Paléocène – Les Mammifères deviennent plus nombreux mais, à l’exception des Marsupiaux et Insectivores, les ordres actuels ne sont pas encore représentés. Existent alors des groupes de placentaires archaïques qui déclineront ou s’éteindront lors de la crise de la limite Eocène – Oligocène (la " Grande Coupure ").
• Les Amblypodes constituent une lignée qui s’éteindra sans descendance fin Eocène. Ce sont gros animaux herbivores mais leur denture montre des canines supérieures développées. Uintatherium (= Dinoceras) d’Amérique du Nord atteint la taille d’un gros rhinocéros ; il est doté de trois paires de protubérances osseuses sur la face antérieure du crâne.
• Les Créodontes
sont des carnivores semi-plantigrades au corps massif et à tête volumineuse. Ils différent des carnivores actuels par leur encéphale lisse et la position variable de la carnassière dans la denture. Les carnivores modernes (Fissipèdes) apparus à l’Oligocène en sont les proches descendants ; Cétacés et Pinnipèdes dérivent de ces carnassiers terrestres.
• Les Condylarthres
sont des herbivores ou omnivores à membres pentadactyles ; ils sont à l’origine des Ongulés actuels (Périssodactyles et Artiodactyles).
Eocène - La faune mammalienne acquiert un cachet moderne.
• Les Marsupiaux se dispersent à partir de l’Amérique du Nord ( ?) ; ils atteignent l’Australie dès l’Eocène basal par l’Amérique du Sud et l’Antarctique. En Australie et en Nouvelle Guinée, l’isolement géographique ultérieur les met à l’abri des Placentaires et entraîne là une vaste radiation adaptative. De l’Amérique du Nord, ils atteignent l’Europe durant l’Eocène (la Sarigue de Cuvier trouvée à Montmartre date de l’Eocène supérieur à –35 Ma) puis, à l’Oligocène, ils gagnent l’Afrique du Nord. En Asie, les Marsupiaux (Asiatherium, Deltatheridium) sont connus au Crétacé supérieur (Campanien).
• Les Placentaires se diversifient et se dispersent à partir de l’Asie. Les principaux ordres se développent (Insectivores, Lémuriens) ou apparaissent (Rongeurs simplicidentés, Edentés, Chiroptères, Siréniens, Carnivores fissipèdes, Cétacés et Ongulés) dérivés des Créodontes et des Condylarthres du Paléocène.

(P = Placentaires originaires de Mongolie, M = Marsupiaux originaires d’Amérique du Nord ; 1 = Eurasie, 2 = Afrique, 3 = Inde, 4 et 5 = Amériques du Nord et du Sud : 6 = Antarctique)
II – La radiation mammalienne cénozoïque et la dynamique des masses continentales
A) Expansions et confrontations des Placentaires et Marsupiaux
Au Crétacé inférieur (Albien à – 105 Ma) apparaissent les premiers Marsupiaux – en Amérique du Nord ( ?) – et les premiers Placentaires en Asie (Mongolie).
Le Crétacé supérieur est une période pendant laquelle les Marsupiaux connaissent une extension géographique mondiale qui les conduit en Amérique du Sud, Antarctique et Australie ; ils constituent alors le groupe dominant de Mammifères thériens, accompagné des protothériens Multituberculés.
Début Tertiaire, les Placentaires se diversifient à partir de l’Asie centrale et, dès le Paléocène, ils envahissent le Nouveau Monde par les voies Nord Pacifique et/ou Nord Atlantique. Ils supplantent désormais les Marsupiaux qui ne subsistent comme groupe dominant que sur les terres isolées : Amérique du Sud et Australie / Nouvelle Guinée.
Fin du Tertiaire (Pliocène supérieur), l’émersion de l’isthme de Panama met en communication les territoires Nord et Sud américains. L’Amérique du Sud est envahie par les Placentaires venus du nord ; l’arrivée des carnivores placentaires entraîne la disparition presque totale des Marsupiaux.
Au Pléistocène, l’Australie reçoit tardivement quelques Placentaires venus d’Asie par les îles du sud-est asiatique.
B) Les relations entre faunes d’Amérique du Nord et d’Amérique du Sud ("le grand échange")
L’Amérique du Sud est isolée au Nord pendant presque tout le Cénozoïque jusqu’à la mise en place de l’isthme de Panama il y a 2,5 Ma. Pendant ces 60 Ma d’isolement, il s’y diversifie une faune mammifère endémique de Marsupiaux dont aucune famille n’a de représentant en Amérique du Nord. Dans chaque Amérique, les mêmes niches écologiques sont occupées par des placentaires (au Nord) et des marsupiaux (au Sud) qui montrent les mêmes adaptations, les mêmes convergences de formes (ex : le carnivore marsupial Thylacosmilus et son pendant placentaire Machairodus).
Pendant l’Eocène : Amérique du Nord et Amérique du Sud sont isolées par des niveaux marins élevés. Des attaches temporaires permettent le passage de quelques Placentaires d’Amérique du Nord en Amérique du Sud. Là se développent des Ongulés singuliers (ex : Macrauchenia) et des représentants de l’ordre des Edentés (Paresseux, Tatous, Fourmiliers) en compagnie des Marsupiaux indigènes mais il n’existe pas à cette époque de carnivore sud-américain autre que marsupial.
Pendant l’Oligocène : l’abaissement du niveau marin met en place une guirlande d’îles dont profitent des rongeurs nord-américains pour atteindre l’Amérique du Sud. Là, ils se diversifient en rongeurs sud-américains typiques (cabiais, maras, chinchillas et cobayes).
Au Pliocène supérieur : le rattachement des terres par l’isthme de Panama ouvre la voie au " Grand Echange ". Il est marqué par la migration massive Nord-Sud de placentaires. Des espèces différentes adaptées aux mêmes niches écologiques entrent en compétition mais les espèces placentaires issues d’Amérique du Nord, peut-être mieux adaptées, disparaissent en moins grand nombre de sorte que les placentaires immigrants supplantent les groupes sud-américains (Placentaires ongulés indigènes, Marsupiaux carnivores et Marsupiaux en général). Une migration Sud-Nord beaucoup plus discrète conduit des animaux sud-américains tels que Opossum et Tatou en Amérique du Nord.
C) La crise Eocène-Oligocène ou " grande coupure " ( – 34 Ma)
A la fin de l’Eocène, les Mammifères atteignent leur diversité maximale (122 familles) et tous les types adaptatifs connus actuellement sont déjà présents. La crise voit la disparition de groupes archaïques dès la seconde moitié de l’Eocène et l’essor des groupes modernes dès la base de l’Oligocène mais le nombre de famille (70) reste bien inférieur à celui des temps éocènes. S’éteignent ou déclinent les Multituberculés, les Condylarthres, les Uintathères et les Marsupiaux de l’hémisphère Nord. La nouvelle vague est dominée par les Rongeurs modernes (ils occupent la niche écologique des Multituberculés), les carnivores Fissipèdes (Canidés, Vivérridés et Mustélidés) et les Ongulés.
Un changement climatique serait la cause de cette crise.
L’Eocène précoce est une période planétaire chaude (la plus chaude de tout le Cénozoïque) alors que l’Eocène moyen et tardif est marqué par un net refroidissement et une sécheresse marquée aux hautes latitudes. En effet, les rapports isotopiques de l’oxygène établis sur des coquilles d’invertébrés marins indiquent une chute de 10°C de la température moyenne des eaux du Pacifique ; l’étude des paléoflores éocène et oligocène de la côte ouest nord-américaine indiquent une chute de 8°C. Ainsi, l’Antarctique se couvre de glace et les zones boisées de type tropical se trouvent limitées à une mince bande alors que les forêts tempérées à conifères et arbres caducifoliés se développent dans les zones à climat contrasté. Ce changement de flore a pu perturber le régime alimentaire des phytophages, de leurs prédateurs et donc perturber les réseaux trophiques.
Plusieurs causes ont été invoquées pour ce changement climatique :
• un changement d’inclinaison de l’axe terrestre (celui-ci passant de la verticale à une inclinaison de 23°) aurait entraîné une augmentation des surfaces soumises aux faibles rayonnements solaires et donc un refroidissement général ;
• une perturbation des courants océaniques
aurait amené des eaux froides polaires en domaine intertropical (l’ouverture du passage entre Norvège et Groenland dans l’hémisphère Nord et l’ouverture du passage entre Antarctique et Australie dans l’hémisphère Sud auraient modifié les circulations océaniques et atmosphériques planétaires).
Conclusion générale : deux points peuvent être soulignés ici
• l’addition de changements minimes (micro-évolution) au fil des générations (effet amplificateur du temps) a permis un événement majeur (macro-évolution) : l’émergence des premiers mammifères à partir d’un vivier ancestral reptilien ;
• l’origine reptilienne et la phylogénie des Mammifères initialement fondées sur des bases paléontologiques et systématiques ont été confirmées, au moins dans leurs grandes lignes, par l’établissement des phylogénies moléculaires bâties à partir des séquences de molécules telles que l’hémoglobine, l’insuline, les ARN ribosomiques…...
et ces 2 points s’inscrivent bien dans la Théorie synthétique de l’Evolution.
3.2.7.3 Classification
La diversité des Mammifères Protothériens ou « Monotrèmes »
Deux genres ; Ornythorynchus ou Ornythorynche et Tachyglossus ( deux espèces), Echidnés.
La diversité des Marsupiaux ou Métathériens
Ne sont plus représenté qu’en Australie, en Tasmanie , en nouvelle – Guinée et en Amérique du Sud et du nord
En Amérique du Nord : La Sarigue de Virginie , en Amérique du Sud : Les Sarigues et les Coenolestes, deux petits groupes carnivores.
La majorité des représentants de la sous-classe habitent les territoires de l’Australie
La sous classe des Marsupiaux est ventilée en 10 familles :
Didelphidés : Opossums
Dasyuridés: Diable de Tasmanie ou Sarcophile, Loup marsupial.
Thylacinidés : loup de Tasmanie
Myrmécobiidés : Fourmilier marsupial
Notoryctidés : Taupe marsupiale
Péramélidés : Bandicoot
Péroryctidés : bandicoot géant
Coenolestidés : Coenolestes, Rat opposum
Phalangéridés : Certains Opossums
Phascolomyidés ( Wombatidés) : Le Wombat ou Phascolome
Macropodidés : les Kangourous , Wallabyes.
Phascolarctidés : Koalas
Microbiothéridés : Colocolo
Acrobatidés : acrobate
Burramyidés :Possum pygmée
Potoroïdés : Rat-kangourou
Pseudocheïridés (Phalanger à queue annelée)
Tarsipédidés :Souris à miel
Petauridés : Possum rayé, etc.
Phalangéridés : couscous, etc.
Burramyidés : Possum pygmée
Certains auteurs parlent de l’Ordre des Marsupiaux et non de la Sous classe … question de conventions…
Les Marsupiaux sont apparus comme nous l’avons vu, au Crétacé supérieur Austral ( GONDWANA) avec des formes peu différentes des Sarigues actuelles. Ils se répandent sur tout l’hémisphère Nord . L’ apparition quasi simultanée des Euthériens ne leur permet de se développer que si ils sont isolés géographiquement de ceux-ci. Si les Marsupiaux américains du Nord sont peu différents du type primitif, les Marsupiaux australiens ont put développer un certain nombre de spécialisations en rapport avec les niches écologiques disponibles. En conclusion, en l’absence d’une pression de concurrence avec les Euthériens, ils ont évolué jusque nos jours et développé des adaptations parallèles à celles ces Euthériens :
Chaque type marsupial a son équivalent parmi les Euthériens de l’hémisphère nord
Carnassiers, coureurs, grimpeurs, insectivores, fouisseurs, arboricoles etc…
Sous forme de schémas
Eres géologiques
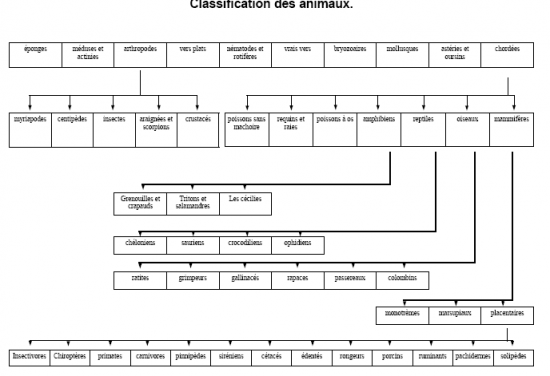
La diversité des animaux
La diversité des vertébrés
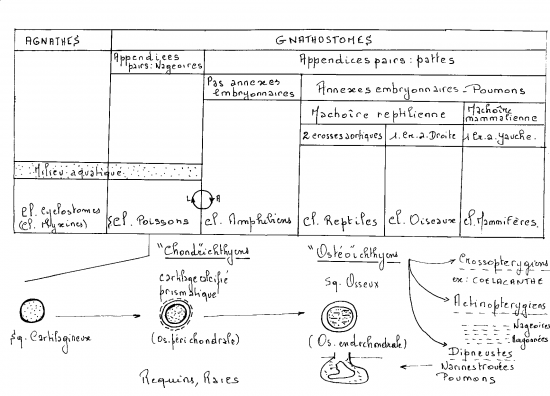

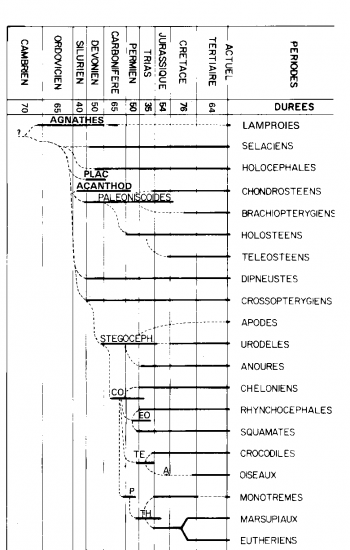
La diversité des Mammifères

Date de dernière mise à jour : 30/03/2018
Ajouter un commentaire