

HISTOIRE DES VERTEBRES 4
3.2.5 La classe des Reptiles
3.2.5.1 Introduction
Les reptiles se sont définitivement affranchis du milieu aquatique pour l’œuf amniotique ( voir organogénèse).
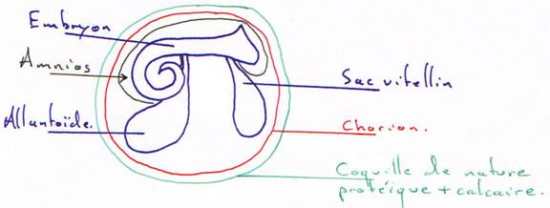
Le développement larvaire est remplacé par le développement de l’embryon dans l’amnios.
La respiration est pulmonaire ; la peau se recouvre de phanères : les phanères épidermiques cornés qui donneront des écailles cornées.
3.2.5.2 Origine et évolution des Reptiles
Il est assez facile de distinguer un amphibien fossile et un reptile par leur squelette : les reptiles montrent des os crâniens de dimensions réduites, une mâchoire articulée au crâne par l’intermédiaire du carré, des vertèbres sacrées soudées entre elles et l’absence du sillon auditif typique des amphibiens. Cependant, si on remonte aux débuts de l’évolution des reptiles, on constate que les différences entre les amphibiens Labyrinthodontes et les premiers reptiles s’amenuisent de plus en plus au point qu’il est possible d’envisager une relation de descendance.
3.2.5.2.1 Les Anthracosaures : à l’origine des Reptiles et des Amniotes
On ne connaît pas de véritable forme de transition entre les amphibiens Labyrinthodontes et les Reptiles mais il existe un groupe d’animaux semblant posséder des caractères de transition : les Seymouriamorphes (des Amphibiens Labyrinthodontes du type anthracosaure) dont le représentant le plus connu est Seymouria.
Seymouria vivait au Permien inférieur alors que les Reptiles, apparus au Carbonifère, étaient déjà très répandus. Il ne peut donc pas être considéré comme l’ancêtre des Reptiles ni comme une forme de transition mais il permet de donner une représentation de ce que furent ces formes de transition.
Gisement : couches permiennes (l’île de Seymour en Amérique du Nord, mais aussi Texas et Russie)
Physionomie : petits animaux à aspect de lézards (taille < 1m)
Squelette : il ne présente pas de caractères intermédiaires entre amphibiens et reptiles mais il réunit une mosaïque de caractères de poisson (2), d’amphibien Labyrinthodonte (18) et de reptile (11) de sorte qu’il s’agit déjà d’un reptile pour certains et encore d’un amphibien pour d’autres (thèse admise par une majorité de paléontologues).
• Quelques uns des caractères d’amphibiens Labyrinthodontes
* dents à structure labyrinthique,
* crâne à sillon auditif attestant d’une ligne sensorielle latérale,
* voûte crânienne épaisse,
* ceinture pelvienne.
• Quelques uns des caractères de reptiles
* choanes allongées en fente,
* 1 seul condyle occipital,
* vertèbres à arcs neuraux très développés et insertion des muscles thoraciques de type reptilien,
vertèbres sacrées soudées entre elles.
Biologie : Seymouria et les Seymouriamorphes subissaient la métamorphose typique des Amphibiens comme le montrent les formes juvéniles (présence d’un arc branchial et d’un sillon sensoriel attestant d’une ligne latérale sensible aux variations du courant de l’eau) ; ils sont donc encore très inféodés au milieu aquatique pour leur reproduction et pour les stades précoces du développement.
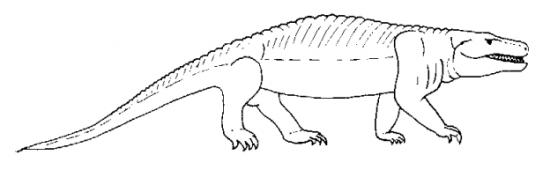
Figure 5 : Seymouria (Labyrinthodonte anthracosaure du Permien)
En conclusion : si l’on admet que les Anthracosaures sont à l’origine des reptiles et donc des premiers amniotes, il faut considérer les reptiles comme un groupe monophylétique mais ceci, loin d’être unanimement admis, est l’objet d’un vif débat soumis à la découverte de nouveaux fossiles ou au réexamen de fossiles déjà inventoriés.
Dans tous les cas, les Reptiles supplantent les Amphibiens dès le Permien.
3.2.5.2.2 Les innovations des Reptiles
Les plus anciens reptiles fossiles proviennent de terrains carbonifères (environ 340 MA pour Hylonomus du Carbonifère moyen du Canada). Les reptiles sont plus évolués que les amphibiens car ils ont complètement abandonné le milieu aquatique pour leur ponte grâce à une grande innovation : l’œuf amniotique qui possède une coquille protectrice rigide et qui renferme les réserves nutritives abondantes (vitellus et ovalbumine) ainsi que le milieu liquide (amnios et liquide amniotique) indispensables au bon développement de l’embryon.
Ainsi, l’embryon se développe bien dans un milieu aquatique mais au sein d’un œuf pondu à terre. Des œufs fossiles amniotes sont bien connus et fournis par des couches permiennes. La respiration pulmonaire perfectionnée va de pair avec l’existence d’une peau protégée d’une couche cornée écailleuse (la peau a perdu sa fonction respiratoire si essentielle chez les Amphibiens).
D’autres perfectionnements tels que la thermorégulation, l’osmorégulation se sont sans doute mis en place progressivement.
3.2.5.2.3 Caractères généraux des Reptiles actuels et fossiles
Squelette : animaux tétrapodes (corps soutenu par un squelette interne et 4 membres),
cou individualisé et tête mobile (articulation du crâne à la colonne vertébrale par atlas et axis),
présence d’un palais osseux séparant un étage buccal dévolu à la nutrition et un étage
olfactivo-respiratoire (l’animal peut déglutir sans s’arrêter de respirer) ;
Reproduction : fécondation interne efficace,
oviparité (ovoviviparité chez des vipères et des ichthyosaures),
oeuf amniote (développement direct dans l’oeuf sans métamorphose) ;
Biologie et physiologie : peau sèche kératinisée et peu glandulaire limitant la déshydratation,
ectothermie (la température corporelle dépend de la température extérieure mais ce caractère est discuté chez certains dinosaures),
cœur imparfaitement cloisonné conduisant au mélange des sangs (les crocodiles ont le cœur presque entièrement cloisonné et cela semble être le cas pour certains dinosaures),
bonne coordination entre perception sensorielle et activité motrice (cerveau évolué avec néopallium).
Tous ces caractères réunis chez le même animal définissent tous les reptiles, actuels comme fossiles, au delà de leur indéniable diversité.
3.2.5.2.4 Les différentes lignées évolutives
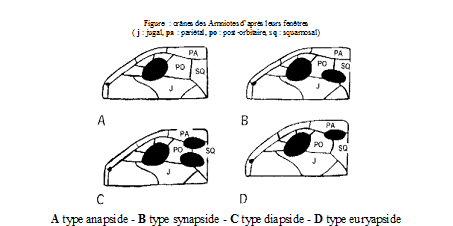
L’évolution des Reptiles s’étend sur près de 300 millions d’années. La classification de WILLISTON y distingue plusieurs lignées selon le nombre et la position des fenêtres temporales :
• Crâne anapside : aucune fenêtre temporale (cas des tortues et des reptiles primitifs tels que les Cotylosauriens)
• Crâne diapside : deux fenêtres temporales (cas de la grande majorité des reptiles, y compris les Dinosaures, Ptérosaures et les Lépidosauriens qui conduisent aux lézards et serpents actuels)
• Crâne euryapside (ou parapside) : une petite fenêtre temporale en position haute (cas des reptiles aquatiques du Mésozoïque tels que Plésiosaures, Mosasaures et Ichtyosaures)
• Crâne synapside : une seule fenêtre temporale en position basse (cas des reptiles mammaliens)
Au sein de ces lignées, les reptiles mammaliens regroupent les Pélycosauriens et les Thérapsidés (tous fossiles) qui sont à l’origine des Mammifères.
Les reptiles mammaliens constituent avec les Mammifères l’ensemble des Théropsidés incluant tous les vertébrés à affinités mammaliennes ; les Théropsidés sont tous synapsides.
3.2.5.2.5 Aperçu de la diversité des Reptiles du Secondaire
Le règne des Reptiles s’intercale entre celui des Amphibiens (durée voisine de 80 millions d’années) et celui des Mammifères (débuté il y a 65 millions d’années) ; il s’étale sur les 225 millions d’années du Permien et de l’ère secondaire que l’on peut à juste titre qualifier d’ère reptilienne.
1. en milieu terrestre
: les Dinosauriens (Dinosaures)
Ils comptent les plus grands animaux terrestres ayant existé mais ils conservent cependant des membres parasagittaux. On y reconnaît 2 groupes distincts par leur ceinture pelvienne :
• les Saurischiens ou Sauripelviens (étym. : " reptiles à hanche de lézard ") à ceinture pelvienne de type reptile (bassin triradié) ;
• les Ornitischiens ou Avipelviens (étym. : " reptiles à hanche d’oiseau ") à ceinture pelvienne de type oiseau (bassin tétraradié : le pubis présente une ramification parallèle à l’ischion).
Figure : structure du bassin des Dinosaures (il : ilion ; is : ischion ; pu : pubis ; pp : prépubis)

A - Sauripelviens bassin triradié B - Avipelviens bassin tétraradié
a. les Saurischiens ou Sauripelviens
Ils regroupent des formes quadrupèdes – les géants du groupe – et des formes bipèdes.
On distingue les Sauropodes herbivores (ou phytophages) et les Théropodes carnivores prédateurs ou/et nécrophages (charognards). Exemples :
• Apatosaure ou Brontosaure (genre quadrupède)
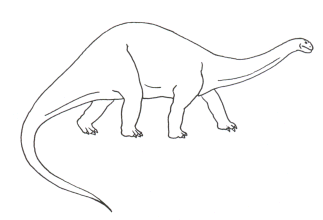
o environ 20 m de long pour un poids estimé de 35 tonnes (le poids de 6 éléphants adultes)
o extension spatio-temporelle : Jurassique supérieur d’Amérique du Nord
o documents : squelettes entiers, empreintes de pas
o régime alimentaire : herbivore mais il ne pouvait pas mâcher (implantation des dents uniquement à l’avant de la bouche)
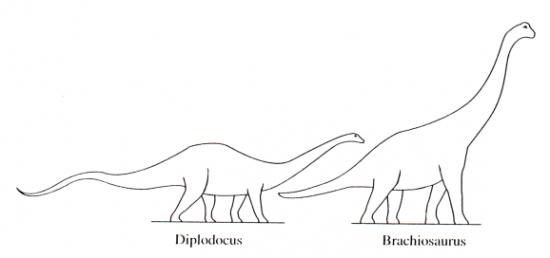
o milieu et mode de vie : animal forestier. Sur l’argument de la position haute des narines – façon hippopotame –il fut autrefois installé en marécages, le mode de vie semi-immergé étant supposé compenser partiellement son poids, les narines hautes autorisant la respiration sous l’eau. Cependant, certains firent remarquer la similitude de ses membres avec ceux des éléphants et des rhinocéros et ils montrèrent que l’immersion rendait impossible la respiration de ces animaux du fait de la pression exercée par l’eau sur la cage thoracique. L’animal fut dès lors considéré comme forestier ; il devait s’y nourrir à la manière des girafes atteignant les feuillages à l’aide de son long cou. Cette thèse bien admise aujourd’hui est généralisable aux genres voisins : Diplodocus, Brachiosaurus.
• Tyrannosaure (genre bipède)
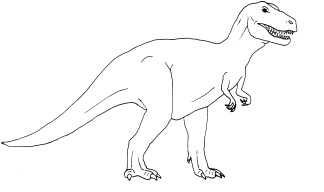
o 6 m de haut pour 12 m de long
o extension spatio-temporelle : Crétacé supérieur d’Amérique du Nord et d’Asie
o documents : squelettes entiers, empreintes de pas
o régime alimentaire : carnivores prédateurs plutôt que charognards, dents toutes semblables pointues à bords crénelés de 15 à 20 cm de long
o milieu et mode de vie : membres antérieurs réduits inaptes à la locomotion et membres postérieurs puissants ; les empreintes rapprochées des pieds et de la queue indiquent que ceux-ci devaient former une sorte de trépied au repos alors que les empreintes de pieds éloignées et l’absence d’empreinte de la queue laissent imaginer une course bipède rapide avec la queue relevée.
b. les Ornitischiens ou Avipelviens (ils sont tous phytophages)
• Stégosaure (genre quadrupède)
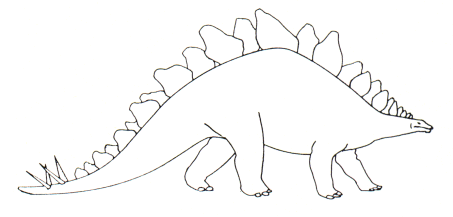
o 9 m de long
o extension spatio-temporelle : Jurassique d’Amérique du Nord
o documents : squelettes complets montrant un crâne très petit, 2 rangées de plaques osseuses dermiques dressées sur le dos et disposées en quinconce et des pointes osseuses (1 m de long environ) sur l’extrémité de la queue.
o régime alimentaire : herbivore
o milieu et mode de vie :
* les moulages endocrâniens montrent un cerveau réduit, guère plus large que la moelle épinière, d’où ont été déduits chez cet animal des mouvements simples et lents ainsi qu’une prédominance de l’activité réflexe;
* il a été attribué aux plaques osseuses dorsales un rôle défensif mais leur structure et l’expérimentation vont dans le sens d’un rôle thermorégulateur (le sang devait y circuler dans un vaste réseau de lacunes et permettre soit l’évacuation de l’excès de chaleur corporelle soit l’absorption de la chaleur solaire) ;
* les pointes osseuses caudales devaient jouer un rôle défensif (prédateurs tenus à distance et/ou blessés par les balancements de la queue).
• Tricératops
(genre quadrupède)
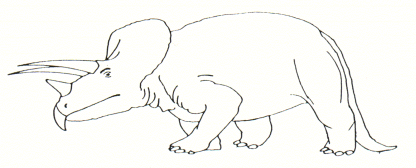
o 9 m de long pour un poids estimé de 11 tonnes
o extension spatio-temporelle : Crétacé supérieur d’Amérique du Nord
o documents : squelettes complets, présence d’un bec corné, de 3 cornes (2 frontales et 1 nasale) à rôle sans doute défensif sinon dissuasif et d’un couvre-nuque protecteur (?) de l’arrière tête et du cou
o régime alimentaire : herbivore
o milieu et mode de vie : l’abondance des fossiles dans les gisements laisse supposer une vie en troupeau.
• Iguanodon
(genre bipède)
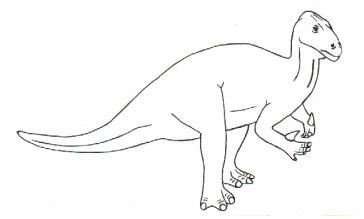
o 3 ou 4 m de haut pour 9 m de long, aspect de Tyrannosaure
o extension spatio-temporelle : Crétacé inférieur d’Europe
o documents : squelettes complets très bien conservés (Iguanodon de Bernissart – Belgique)
o régime alimentaire : herbivore consommateur de fougères et de prëles (dents à surface plane dotée de replis verticaux comparables à ceux observés chez les vaches et les chevaux)
o milieu et mode de vie : coureurs rapides ; l’abondance des fossiles dans les gisements laisse également supposer une vie en troupeau.
2. en milieu marin :
• Ichthyosaures (du Trias au Crétacé) (étym. " reptiles poissons ")
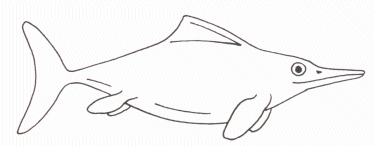
o 1 à 8 m selon les genres,
o corps hydrodynamique (aspect de dauphin) couvert d’écailles et porteur de nageoires dorsales et caudales
o membres transformés en palettes natatoires (hyperdactylie et hyperphalangie, le tout enveloppé dans un conjonctif commun)
o nage par ondulations du corps aidé des nageoires dorsales et caudales, palettes natatoires à rôle stabilisateur
o nourriture : les proies sont des poissons et des céphalopodes capturés à l’aide du museau long et fin, bouche garnie de nombreuses dents coniques semblables (certains spécimens montrent des écailles de poissons et des rostres de bélemnites conservés à l’emplacement du tube digestif).
o reproduction : certains spécimens présentent dans le ventre les squelettes de 1 ou 2 petits ichthyosaures (ce sont toujours des petits appartenant à la même espèce mais il ne s’agit pas de cannibalisme car ils ne sont pas endommagés par l’action de dents ou par la digestion ; il s’agit donc d’embryons) ; cela atteste d’une viviparité (viviparité stricte ou ovoviviparité ?). Les plus gros embryons sont toujours disposés dans la même position
o – tête orientée vers la tête maternelle – et devaient donc naître à la manière des baleineaux en présentant la queue (ceci permet l’apprentissage des premiers battements de queue et évite la noyade instantanée qu’occasionnerait une naissance brutale tête en avant).
• Plésiosaures (Jurassique inférieur à Crétacé supérieur d’Europe et d’Asie)

o 1 à 15 m selon les genres,
o petite tête, cou long et grêle sur un corps massif
o queue courte et puissante
o membres transformés en palettes natatoires (hyperphalangie)
o nage lente à l’aide des membres transformés en palettes (à la manière des tortues)
o nourriture : poissons, céphalopodes.
• Mosasaures (Crétacé supérieur d’Europe)

o 4 à 10 m selon les genres,
o grosse tête et cou ramassé sur un corps long et souple, serpentiforme
o queue longue et puissante dotée d’une nageoire caudale
o membres transformés en palettes natatoires (hyperphalangie)
o nage par ondulations du corps, membres à rôle stabilisateur
o nourriture : poissons et céphalopodes.
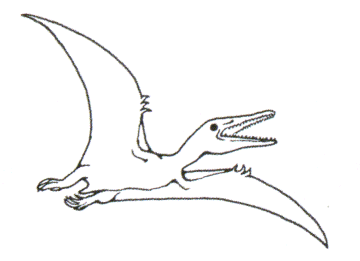
3. en milieu aérien : les Ptérosauriens (étym. " reptiles ailés ")
o De nombreux genres sont connus issus de gisements célèbres tels ceux de Cerin (France), Solenhofen (Allemagne) et du Kazakhstan ; là, des calcaires lithographiques ont fournis d’excellents fossiles.
o Leur étude indique que l’aile est formée par un repli de peau tendu entre le membre postérieur, le corps et le quatrième doigt hypertrophié du membre antérieur. Le squelette est constitué d’os pneumatiques : la cavité axiale est remplie d’air ce qui confère à l’animal une faible masse volumique favorable au vol. Les moulages endocrâniens montrent un cervelet développé (le cervelet renferme les centres de la motricité !) et des lobes optiques très développés. En conclusion, ces animaux dotés d’une bonne vision devaient être très actifs.
o Les empreintes de peau portent des traces de glandes sébacées : il en a été déduit l’existence de poils, poils évitant ou limitant la déperdition de chaleur en particulier au cours du vol.
o Cependant, le squelette des ailes est pauvre en insertions musculaires et l’articulation " humérus/scapulaire " ne permet pas un vol ramé efficace. On a imaginé ces animaux pendus au repos la tête en bas à la manière des chauves-souris et se libérant pour effectuer des vols essentiellement planés.
o Principaux genres connus : entre 0,5 et 12 m d’envergure selon les cas
Ptérodactyle :
Jurassique supérieur d’Afrique, taille d’un cygne mais moins de 1 m d’envergure, dents pointues et denture rappelant celle d’un insectivore, chasseur d’insectes et pêcheur de petits poissons.
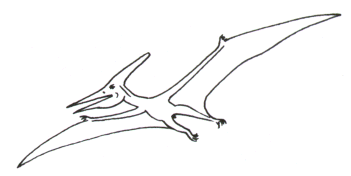
Ptéranodon :
Crétacé supérieur d’Amérique du Nord et d’Europe, taille d’un albatros mais 5 à 8 m d’envergure pour un poids estimé de 12 kg seulement, long bec sans dents, pêcheur de poissons à la manière du pélican (présence d’une poche sous le bec).
Quetzalcoatlus :
Crétacé supérieur du Texas, 11 à 15 m d’envergure pour un poids estimé de 65 kg ( le plus grand animal ayant jamais volé) ; doté d’un grand bec, il est selon les auteurs soit un charognard se nourrissant de carcasses de dinosaures soit un consommateur de poissons et de coquillages prélevés en eaux peu profondes.
3.2.5.2.6 Quelques questions soulevées par l’expansion des Reptiles au Secondaire
1. Sang froid ou sang chaud ?
Un peu de vocabulaire
• Certains vertébrés comme les poissons et les reptiles régulent leur température corporelle en utilisant le milieu ambiant : soleil pour se réchauffer et ombre pour se refroidir comme le pratiquent les lézards ; ils sont qualifiés d’ectothermes. Chez eux, la température corporelle peut varier de 20°C ou plus sans dommages pour l’organisme. Ce système a l’avantage d’être peu coûteux en aliments et oxygène mais présente un inconvénient majeur : l’activité de l’animal est tributaire de la chaleur ambiante.
• D’autres vertébrés comme les oiseaux et les mammifères maintiennent une température corporelle constante grâce à un système sophistiqué de thermorégulation interne ; ils sont qualifiés d’endothermes. Chez eux, une variation de la température corporelle de quelques degrés peut avoir de graves conséquences comme c’est le cas dans l’espèce humaine. Chez ces animaux, maintenir une température corporelle élevée et constante impose une forte dépense énergétique et nécessite donc des apports soutenus et réguliers en aliments et enoxygène. A l’inverse, l’activité de l’animal peut être maintenue malgré les fluctuations du climat.
• On disait à tort animaux à " sang chaud " ou " à sang froid " mais cette formulation doit être abandonnée car la température corporelle d’un ectotherme peut dépasser temporairement celle d’un endotherme
• Une autre approche plus ancienne distingue les vertébrés poïkilothermes à température corporelle variable et les vertébrés homéothermes à température corporelle constante. Ces définitions ne se recouvrent pas parfaitement, ainsi poïkilotherme n’est pas l’équivalent d’ectotherme et homéotherme n’est pas l’équivalent d’endotherme. A titre d’exemples, les poissons sont le plus souvent des ectothermes homéothermes : leur température corporelle varie très peu car elle se trouve régulée par la stabilité thermique de leur milieu de vie marin ; à l’inverse, colibris et chauve-souris sont des endothermes poïkilothermes capables de suspendre leur thermorégulation pendant les périodes nocturnes et hivernales.
Qu’en était-il des dinosaures et autres reptiles du secondaire ?
Dans la faune actuelle, les reptiles sont à ranger clairement parmi les ectothermes. Peut-on sans contestation ni discussion généraliser cela aux reptiles du secondaire qui ont traversé une longue évolution (près de 160 MA) marquée par une très grande diversité des genres, tailles et milieux de vie ?
• Le climat au Jurassique - Crétacé
. Durant l’ère secondaire, le climat est nettement plus chaud que l’actuel et le gradient thermique latitudinal est faible ; on ne connaît pas de calotte glaciaire à cette époque. Cette douceur favorable peut expliquer la présence des dinosaures sur tous les continents jusqu’à 60° de latitude nord et sud ; de nombreux reptiles du secondaire devaient être de simples ectothermes.
• Et les dinosaures géants
? Chez eux, le rapport " surface/volume " est relativement faible de sorte que, même avec un métabolisme bas, la chaleur produite par les tissus profonds est équilibrée par la faible déperdition thermique cutanée. Ces grands dinosaures herbivores devaient être des ectothermes homéothermes et l’usage a consacré les termes d’ homéothermie liée à la masse ou encore de gigantothermie.
• Les ichthyosaures et autres reptiles marins
? Les deux raisonnements précédents peuvent leur être appliqués. Ils bénéficient de la douceur climatique et de la température favorable des eaux marines et peuvent être considérés comme des ectothermes homéothermes (homéothermie liée au milieu de vie). De plus, les espèces les plus volumineuses devaient réaliser une gigantothermie (10 à 15 m de long et un corps massif à faible rapport " surface/volume ").
• Les dinosaures polaires
: migration ou hibernation ? Dans les années 60, les campagnes de prospection pétrolière d’Alaska permirent la découverte d’ossements de dinosaures dans des roches sédimentaires du Crétacé. Or au Crétacé, cette région se trouvait déjà au nord du cercle polaire. Dans les années 90, d’autres dinosaures ont été découverts dans l’hémisphère sud largement au delà du cercle polaire : Jurassique et Crétacé de l’Antarctique, Crétacé inférieur du Sud de l’Australie. Ces découvertes ont nourri un vif débat sur la thermorégulation de ces dinosaures polaires. Pour les partisans de l’endothermie, même en absence de calotte glaciaire, l’hiver polaire devait être rigoureux. Ces dinosaures, tous de petite taille, devaient donc être adaptés au climat froid puisque seul un endotherme peut supporter sur de longues périodes des basses températures. Quelle adaptation ? L’endothermie vraie ? elle suppose un fort métabolisme et une prise de nourriture abondante et régulière. L’hibernation ? elle permet de résoudre le passage de la mauvaise saison mais les preuves manquent. La thermorégulation sociale à la manière des manchots ? Là encore, les preuves manquent. Les adversaires de l’endothermie font remarquer qu’il n’y a pas de preuves indiscutables de la présence de neige ou de glace aux pôles à cette époque, donc pas de preuve d’un hiver rigoureux. En outre, les dinosaures polaires auraient très bien pu migrer vers des latitudes plus clémentes au gré des disponibilités en nourriture comme le font aujourd’hui les rennes. Le débat reste ouvert, soumis aux découvertes futures.
• La présence de phanères ? En 1996, des fossiles de dinosaures théropodes d’une qualité exceptionnelle ont été découverts dans le Crétacé supérieur du nord-est de la Chine : outre le squelette, les parties molles et en particulier la peau étaient bien observables…Or cette peau présente un fin revêtement filamenteux. Qu’il s’agisse de poils ou de duvet, cette couverture épidermique a pu contribuer à l’endothermie en réduisant la déperdition de chaleur.
2. Gigantisme
Les Sauropodes ont compté dans leurs rangs les plus grands animaux terrestres de tous les temps : Diplodocus (Jurassique moyen - Amérique du Nord) atteignait 26 m de longueur et Brachiosaurus (Jurassique supérieur – Tanzanie, Afrique du Nord et Amérique du Nord) 12 m de hauteur. Ces animaux au volume imposant fait de chair et d’os volumineux avaient des masses corporelles élevées : on estime à environ 90 tonnes le poids de Brachiosaurus soit le poids de 12 éléphants adultes.
• Comment estimer le poids d’un dinosaure ? L’estimation a été réalisée à partir de modèles réduits ; ceux-ci sont plongés dans un récipient plein d’eau et le volume d’eau collecté correspond au volume du modèle réduit. Il est alors simple d’obtenir le volume de l’animal, volume auquel il faut appliquer un coefficient multiplicateur : la masse volumique de l’animal. Là, les calculs divergent selon le masse volumique attribuée à 1 litre de dinosaure : plus ou moins de 1 kilogramme ? Variable selon les auteurs (elle se situe pour eux entre 0,7 et 1), elle conduit à des masses corporelles différentes selon les paléontologues.
• De telles masses corporelles étaient-elles mécaniquement supportables ? A mesure que s’accroissent les dimensions d’un animal, le volume corporel et donc la masse corporelle augmentent comme le cube de ces dimensions. Parallèlement, chez un tétrapode marcheur, le diamètre des pattes augmente proportionnellement à la masse corporelle. Il est possible d’établir un graphe donnant le diamètre des os de pattes (fémur, humérus) en fonction de la masse corporelle : la relation est linéaire et il est possible d’extrapoler pour des valeurs très élevées de la masse corporelle. Pour 140 tonnes environ, la limite supérieure du poids d’un animal terrestre est atteinte car le diamètre des os des pattes est tel que celles-ci se touchent. Les plus grands dinosaures n’ont pas atteint cette limite : ils étaient donc mécaniquement viables sur la terre ferme.
NB : ce graphe portant en échelles logarithmiques le diamètre des os de pattes en fonction de la masse corporelle peut être utilisé en sens inverse pour déterminer la masse corporelle à partir du calibre d’un os (humérus, fémur).
3. Capacité cérébrale, intelligence ?
Le tissu nerveux se décompose rapidement après la mort et il échappe le plus souvent à la fossilisation. Malgré cela, le système nerveux central des Vertébrés fossiles peut être étudié à l’aide de moulages endocrâniens ; ils renseignent sur le volume du cerveau et de ses différentes parties, sur les nerfs qui s’y raccordent (à partir des orifices autorisant leur passage) et sur la taille des organes des sens.
La relation entre les tailles du corps et du cerveau a été établie chez tous les vertébrés actuels : à taille corporelle égale, les oiseaux et les mammifères ont un cerveau environ 10 fois plus volumineux que celui des poissons osseux, des amphibiens et des reptiles. Sachant que plus le cerveau est développé, plus le comportement est souple et complexe, on peut appréhender la plasticité comportementale et l’activité générale d’un vertébré à partir de la taille de son cerveau.
Qu’en est-il pour les dinosaures? Les résultats indiquent que la taille relative de leur cerveau est comparable à celle des poissons osseux, des amphibiens et des reptiles actuels ; on peut en déduire une plasticité comportementale et une activité générale comparables à celle de ces animaux. La stupidité et la lourde lenteur longtemps attribuées aux dinosaures doivent être abandonnées.
Cependant, au sein même des dinosaures existent de fortes disparités. Chez les Sauropodes et dans la lignée des Stégosaures le cerveau présente une très faible taille relative alors que chez certains dinosaures carnivores bipèdes, celle-ci atteint le niveau de celle d’un oiseau (Stenonychosaurus - Crétacé supérieur - Amérique du Nord). Ceci révèle sans doute de fortes différences de capacité cérébrale et de souplesse comportementale chez les dinosaures. Rien de bien surprenant dans tout cela si l’on considère la diversification des genres et des espèces réalisée au cours de près de 150 millions d’années d’existence des dinosaures.
Date de dernière mise à jour : 30/03/2018
Ajouter un commentaire