
HISTOIRE DES VERTEBRES 6
3.2.6 La classe des Oiseaux.
3.2.6.1 Introduction
Les Oiseaux peuvent presque être définis comme étant des Reptiles spécialisés car leur architecture est similaire à celle des Reptiles et les modifications peuvent être reliées au vol. Les oiseaux sont des homéothermes endothermes qui ont des plumes. Leurs membres antérieurs sont modifiés pour le vol, et leurs membres postérieurs pour la marche, la nage, ou pour se percher.
Modifications associées au vol
Le vol exige une très grande dépense métabolique, et les forces exercées par les muscles alaires sont également importantes. On retrouve donc chez les Oiseaux des adaptations pour augmenter le taux métabolique qui peut être soutenu, pour réduire les dépenses métaboliques inutiles (réduction du poids), pour augmenter l’efficacité du système cardio-vasculaire (pour maintenir un métabolisme élevé), et pour solidifier le squelette.
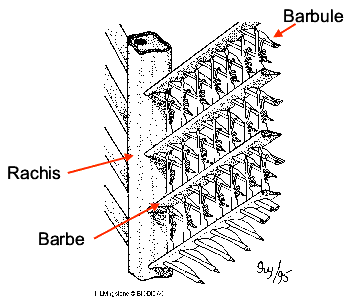
Figure Parties d’une plume d’oiseau: rachis, barbe, barbule. © BIODIDAC
La température influence énormément les réactions métaboliques. L’activité des enzymes qui catalysent les réactions augmente avec un accroissement de la température, jusqu’à un maximum, puis diminue. Pour maintenir les hauts taux métaboliques requis pour le vol, les oiseaux maintiennent leur température corporelle à l’intérieur de limites précises (ils sont homéothermes).
On croit que les plumes étaient à l’origine une adaptation pour conserver la chaleur corporelle (réduction de la convexion), et qu’elles ont été modifiées pour le vol par la suite .
Il est important pour les oiseaux qui volent de minimiser la masse qu’ils ont à transporter. Les os des oiseaux sont généralement creux et renforcés par des cloisons. Chez certains oiseaux, les sacs aériens se prolongent dans les plus gros os.
Figure Coupe d’un os d’oiseau. © BIODIDAC
Chez les oiseaux, l’excrétion des déchets azotés se fait sous forme d’acide urique. Comme les cristaux d’acide urique peuvent être éliminés avec une quantité minime d’eau, l’oiseau n’a donc pas à transporter une masse inutile de liquide.
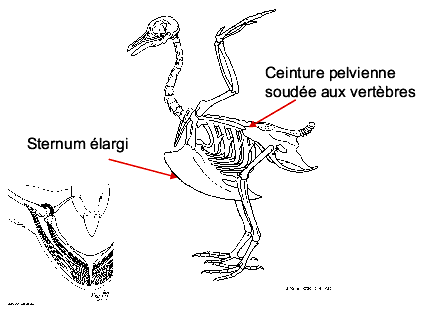
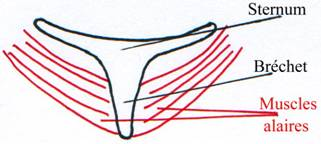
Pour résister aux forces qui entrent en jeu lors du vol, la colonne vertébrale des oiseaux est renforcée par la fusion de plusieurs vertèbres, particulièrement dans la queue et au niveau de la ceinture pelvienne. La cage thoracique doit également être renforcée pour éviter d’être écrasée par les muscles alaires.
Les côtes sont donc fusionnées et le sternum est hypertrophié pour permettre un solide point d’ancrage pour les muscles alaires.
La ceinture pelvienne est également fusionnée pour solidifier les points d’ancrage des muscles de la queue.
Figure Modifications du squelette associées au vol chez les oiseaux. © BIODIDAC
Respiration et circulation
La présence de plumes réduit la ventilation de l’épiderme et empêche les oiseaux d’avoir une respiration cutanée.
Leurs grands besoins métaboliques (de 6 à 10 fois ceux d’un reptile de taille similaire) ont entraîné des modifications au niveau des systèmes respiratoire et circulatoire.
Figure Poumon et sacs aériens chez un oiseau. © BIODIDAC
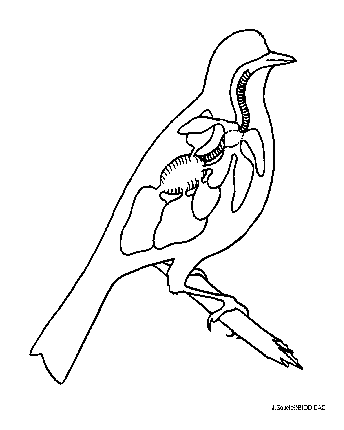
Les poumons des oiseaux ne sont pas des sacs aveugles (culs-de-sac), mais plutôt des conduits dans lesquels l’air circule à sens unique. Il n’y a pas d’alvéoles, mais plutôt des parabronches. La ventilation de ces parabronches est assurée par l’entremise de sacs aériens qui sont situé à l’avant et à l’arrière des poumons, et qui agissent à la manière de pompes. L’air passe de la bouche directement aux sacs aériens postérieurs, puis aux parabronches, puis aux sacs aériens antérieurs, pour être finalement exhalé. Il n’y a donc pas de mélange entre l’air frais et l’air résiduel. La présence de valves empêche l’air de revenir vers l’arrière. Dans les poumons, la circulation du sang des capillaires des parabronches est à contre-courant de celle de l’air, ce qui maximise les échanges gazeux. On considère que le système respiratoire des oiseaux est le plus perfectionné et le plus efficace du règne animal. La ventilation des poumons peut également servir à éliminer les surplus de chaleur (la sudation est impossible à cause de la présence des plumes).
Le coeur des oiseaux a quatre chambres séparées (comme chez les crocodiliens), ce qui prévient le mélange entre de sang veineux et artériel. Chez les oiseaux nordiques, la circulation est à contre-courant dans les pattes, de manière à réduire les pertes de chaleur.
Alimentation et digestion
Les oiseaux ont des besoins métaboliques élevés et doivent donc se nourrir d’aliments riches. Les premiers oiseaux étaient sans doute des insectivores, mais le mode alimentaire des oiseaux actuels est très varié (granivores, piscivores, carnivores, nécrophages, frugivores). La spécialisation alimentaire s’accompagne de modifications au niveau du bec et du tube digestif. Par exemple, les oiseaux granivores, comme la poule, ont un bec robuste pour briser les graines, un jabot pour les entreposer, et un gésier qui contient typiquement des cailloux pour les broyer (les oiseaux n’ont pas de dents).
Excrétion et osmorégulation
Les oiseaux marins ont des glandes à sel semblables à celles des iguanes pour excréter les surplus d’ions.
Système nerveux
Le vol requiert une très bonne vision et une bonne coordination; les zones du cerveau responsables de ces aspects sont donc particulièrement développées chez les oiseaux.
Les yeux sont très développés, et leur position est reliée au régime alimentaire. Chez les oiseaux herbivores, les yeux sont disposés de chaque coté de la tête, ce qui leur permet d’avoir un très large champ de vision et de voir venir les prédateurs. Chez les rapaces, les yeux sont situés à l’avant, ce qui permet d’avoir une meilleure vision binoculaire et de mieux juger des distances et les vitesses.
3.2.6.2 Origine et évolution
Ils sont apparus il y a 180 millions d’années à partir des dinosaures. Ils sont amniotes. Leurs écailles se sont mobilisées pour donner les plumes. Leur température interne est élevée (40 à 41°C) avec un fort métabolisme.
Les oiseaux fossiles avaient des dents, mais ils n’en ont plus : ils ont des mâchoires à bec corné. Le membre antérieur s’est transformé en aile : il ne touche plus le sol. Les oiseaux sont toujours des tétrapodes.
Ils sont dits pseudobipèdes (pseudo car leur colonne vertébrale est restée horizontale).
Les œufs ont une membrane imprégnée de sels calcaires ; les jeunes sont élevés dans un nid.
Un texte de André Holbecq Dernière Mise à jour : 30 août 2004
Quand vous verrez un vol d’oies du Canada passer au dessus de vos têtes, dites-vous: les dinosaures migrent", ainsi écrivait déjà en 1986, Bob Bakker, le paléontologue américain, dans son livre "Dinosaurs hérésies" ( traduit et publié en France en 1990 sous le titre: le ptérodactyle rose et autres dinosaures chez Armand Colin).
Trop longtemps (et encore maintenant) l’archéoptéryx a été montré comme le chaînon manquant, l’intermédiaire entre les dinosaures et les oiseaux, qu’en est-il maintenant ?
Ce premier "dinoiseau", "dinobird" en anglais, et selon la paléontologue Angela Milner présentait des ailes et des plumes, comme les oiseaux actuels mais aussi des caractères reptiliens comme les dents, la queue, etc… "Vieille aile" ou archéoptéryx pour les savants, fut déjà pressenti comme l’archétype du chaînon manquant par Thomas Huxley mais cette thèse fut combattue violemment.
Cependant dans les années 90 des découvertes chinoises ont permis de déduire que les oiseaux sont des petits dinosaures à plumes et à queue courte.
Huxley avait déjà remarqué 35 traits communs entre les deux groupes, mais à son époque ses détracteurs avaient prétendu qu’il ne s’agissait que d’une convergence de formes : deux créatures différentes évoluent indépendamment et finissent par avoir la même apparence (comme par exemple ichtyosaures, requins et dauphins). De plus on pensait que les dinosaures , à sang froid, n’avaient pas le métabolisme adéquat pour se permettre de voler, ce qui consomme une énorme quantité d’énergie. Plus grave, le bréchet n’existait pas chez les dinosaures alors qu’il est indispensable pour le vol avien; et ce qui disparaît dans l’évolution ne peut pas réapparaître! Et puis archéoptéryx , du jurassique supérieur, est trop vieux pour jouer le rôle d’ancêtre des oiseaux puisque les ancêtres potentiels des oiseaux sont les dromaeosaures datant du crétacé moyen. Dans les années 60, John Ostrom et son Deinonychus, reprenant les arguments de Huxley, réalisa une étude systématique comparant les caractères communs aux dromaeosauriens et aux oiseaux .
Dans les années 70, l’idée de dinosaures à sang chaud faisait son chemin, car à défaut de bréchet dinosaurien on avait trouvé des "furcula" (l’os du bonheur) en forme de boomerang ressemblant à celles en forme de V chez les oiseaux. Mais la controverse ne s’en arrêta pas pour autant à cause des embryologistes en 1990, car ceux-ci pensaient que les doigts des oiseaux sont les 2è,3è et 4ème, alors que chez les dinosaures ce sont les 1er, 2è et 3è.
Et toujours au cœur du problèmes: LES PLUMES. Ce problème ne pouvait pas être résolu tant que l’idée que "plumes implique vol", n’était pas remplacée par d’autres utilités indépendantes du vol. Après tout, les chauve-souris les ptérosaures, les papillons volent bien sans plumes ! Encore fallait-il y penser ! Et si l’on avait mieux observé le comportement des oiseaux actuels on se serait rendu compte des rôles fondamentaux des plumes, à savoir :
- caractères spécifiques
- caractères sexuels secondaires
- isolant thermique qui sont, eux, toujours utilisés même chez les oiseaux qui ne volent pas comme les autruches, casoars, émeus..
Enfin et heureusement les dernières découvertes chinoises permettent de dire qu’il y a autant de preuves de l’existence de dinosaures à plumes que de ptérosaures poilus; ce sont des faits maintenant reconnus.
Donc "la plume ne fait plus l’oiseau" comme l’annonçait le paléontologue Philippe Taquet à Valenciennes lors de l’exposition de l’iguanodon bernissartensis au musée des beaux arts, le 14 octobre 1998. Conséquence inattendue : LES DINOSAURES N’ONT PAS DISPARU, ILS SONT PARMI NOUS !
Et il va falloir revoir la classification animale et certaines définitions, en effet qu’est-ce qu’un oiseau maintenant ?
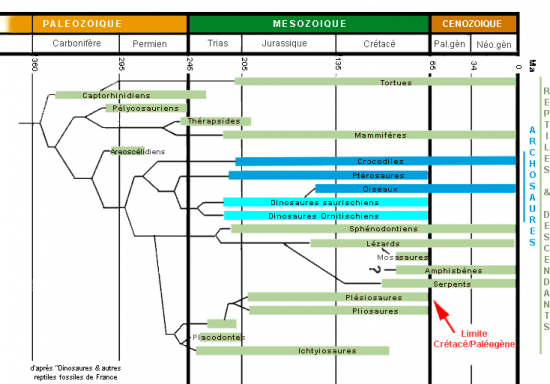
ARCHEOPTERYX LITHOGRAPHICA ou "vieille aile du calcaire lithographique"
Le premier, trouvé en 1861 à Solnhofen en Allemagne, date du jurassique supérieur (150 000 000 d’années) C’est un magnifique squelette avec des empreintes de plumes, parfaitement conservé dans ce sédiment à grain très fin préservant tous les détails. C’est l’exemplaire londonien, sans tête, mais avec furcula (os du bonheur). Son "inventeur", Karl Häberlein, l’a vendu au MNHN de Londres à Richard Owen 750 Livres.
En 1877 le 2ème exemplaire, avec sa tête, fut découvert par le fils d’Häberlein et vendu au musée de Berlin où il est toujours exposé. C’est le plus beau et le plus souvent présenté dans les manuels (avec tête mais sans furcula).
Un 3ème exemplaire a été trouvé en 1956: exemplaire du Maxberg malheureusement déjà décomposé lors de la fossilisation, sans crâne sans queue, mais bonnes empreintes des plumes des ailes. Il est en collection privée. Un 4ème exemplaire découvert en 1855 celui de Haarlem, fut tardivement identifié (en 1970) car il n’existe que les parties extrêmes des membres, et il fut à tort considéré comme ptérosaure.
Un 5ème exemplaire, un juvénile, a été trouvé en 1951 à Eichstätt, pris à tort pour un dinosaure compsognathus, mais identifié correctement en 1973; j’y ai personnellement observé un détail typiquement dinosaurien : une homologie, une astragale avec processus ascendant développé. Des gastralies ou cotes ventrales typiquement dinosauriennes sont bien visibles. Son crâne est remarquablement bien conservé. Les empreintes de plumes sont difficilement visibles (lumière rasante nécessaire).
Le 6ème et dernier a été trouvé en 1987 dans une collection privée à Eichstätt mais il est actuellement au musée de Solnhofen : les membres sont très bien conservés. De puissantes griffes très incurvées aux ailes, en font un grimpeur comme le poussin d’hoatzin actuel.
La plume ne faisant plus l’oiseau, que révèle l’étude du squelette des archéoptéryx ? Archéoptéryx est-il un oiseau ou un dinosaure à plumes ?
-Son museau long et étroit n’est pas un bec, on l’observe aussi chez gallimimus et les ornithomimidés.
-Il a des dents recourbées de type dinosaure, comme on le voit bien sur l’exemplaire d’Eichstätt et les oiseaux n’ont plus de dents.
-La fenêtre antéorbitaire existe distinctement, elle n’est pas du tout fusionnée avec l’orbite oculaire.
-Grandes orbites et gros cerveau mais certains dinos théropodes aussi.
-La zone de fixation du basin sur la colonne vertébrale est bien moins étendue que chez les oiseaux.
-La queue vertébrée est très longue comme chez les dinos.
-Il n’existe pas de pygostyle (=croupion) quelques vertèbres soudées en bout de queue (pourtant certains dromaeosaures en ont un !).
-Les pseudo "ailes" sont différentes de celles d’oiseaux: trois longs doigts griffus libres au lieu des doigts réduits et soudés, non griffus des oiseaux; de plus l’embryologie dit que ce sont les doigts 1,2,3 comme chez les dinos et non pas les 2,3,4 comme chez les oiseaux !
-La ceinture scapulaire montre une fourchette ou " furcula " ou " os du bonheur " en forme de boomerang et sans hypocléide, non pas en V avec hypocléide comme chez les oiseaux.
-La ceinture pelvienne a un ilion court et étroit de dinosaure, non pas long et large d’oiseau.
-L’ensemble pubis + ischion sans soudure, avec pré pubis , est de type théropode.
-Le hallux ou pouce inversé longtemps prétendu être uniquement avien existe pourtant chez les dinos théropodes et il est bien plus court que chez les oiseaux.
-Les os ne sont pas pneumatisés, pourtant ils le sont chez les oiseaux et les ptérosaures, et donc le squelette est relativement trop lourd pour voler.
Tout cela fait d’archéoptéryx un dinosaure emplumé ! (cqfd)
Une découverte dans le Middle West des USA implique une diversité déconcertante chez les dinosaures et amène Gerhard Heilmann à écrire déjà en 1926 sur l’origine des oiseaux en se basant sur l’anatomie et l’embryologie des reptiles, oiseaux, archéoptéryx et autres reptiles fossiles… Il pense que les archosaures et parmi eux les dinosaures saurischiens, et parmi ces derniers les coelurosaures seraient à l’origine des oiseaux. Mais il y a un problème ...
Le problème c’est qu’il n’y a pas de clavicules chez les coelurosaures, donc pas de futures fourchettes possibles !
S’agirait-il donc d’une convergence évolutive ?
Faut-il chercher plus avant chez les archosaures plus primitifs du Trias ? Mais alors il s’agirait d’animaux arboricoles " parachutistes " adeptes du vol plané. Mais cette hypothèse est remise en question avec les trois hypothèses suivantes :
1°/ selon Peter Galton (1970) les ornitischiens seraient proches des oiseaux mais ces dinosaures sont trop spécialisés pour devenir des oiseaux, d’où il faut chercher au Trias un archosaure ornitischien. Mais il n’a jamais été trouvé !
2°/ selon Alick Walker, Sphénosuchus, un archosaure à allure de crocodile ferait l’affaire; il se base sur les os du crâne, la structure de la ceinture scapulaire, sur les membres antérieurs et sur la cheville; mais il oublie le cas: archéoptéryx; il se rétracte et opte pour la 3ème hypothèse.
3°/ John Ostrom prétend que l’archéoptéryx a plus de 20 caractères communs avec les coelurosaures. Heureusement, on découvre des clavicules chez des théropodes, de plus la fourchette des oiseaux n’est pas l’équivalent des clavicules.
Sa théorie implique que :
- le bassin d’archéoptéryx n’est pas du type ornitischien mais saurischien théropode ;
- les membres antérieurs sont très semblables à ceux de théropodes même dans les plus petits détails ;
- les membres postérieurs sont très proches de ceux des théropodes.
Alors s’agirait-il d’une convergence évolutive de formes ?
Non ! Plutôt un lien de descendance !
Une 4ème hypothèse ( Hecht et Tarsitano) reprend les arguments d’Heilmann, et prétend qu’Ostrom se tromperait sur certains points.
L’embryologie permet de penser que chez l’oiseau ce sont les doigts 2,3,4, alors que chez les théropodes, ce sont les doigts 1,2,3, et archéoptéryx possède les doigts 1,2,3. Donc une main de dinosaure théropode.
D’où l’archéoptéryx serait alors un dinosaure à plumes, un "cul de sac de l’évolution".
Actuellement un consensus majoritaire opte pour l’hypothèse de l’ascendance coelurosaurienne.
Dans ce cas deux hypothèses concernant le mode de vol sont proposées :
1°/ soit voler à partir du sol, l’aile donnant de la vitesse pour la chasse (selon Nopsca) Mais cette proto-aile en réalité freine plus qu’elle n’accélère la vitesse de déplacement, alors Ostrom invente l’aile capteur-piège à insectes, avec en conséquence la première étame du vol ramé. Courir ailes déployées implique un effet ascensionnel diminuant la force d’appui au sol pourtant nécessaire pour la course ! Je note au passage que si cela avait été efficace, il existerait encore aujourd’hui quelques oiseaux utilisant cette méthode de chasse, or ils n’existent pas ! Le seul est un héron qui pèche en statique, en déployant ses ailes en ombrelle pour attirer les poissons dans une zone d’ombre.
2°/ soit planer à partir des arbres selon Heilmann, qui voit un proto-oiseau arboricole qui amortit sa chute et plane d’arbre en arbre pour vite changer de milieu, compte tenu des griffes très incurvées aux ailes comme aux pieds.
A l’ouest il y a du nouveau !
À 60 km de Lisbonne, Octavio Mathéus, un amateur découvre des œufs de dinos avec des embryons qui sont étudiés par Philippe Taquet et Armand de Riclès (publication en février 2001). L’embryon in ovo de LOURIHANOSAURUS âgé de 150 MA est le plus vieux des embryons de théropodes. L’étude démontre une croissance très rapide, supérieure à celle des tortues ou des crocodiles et comparable à celle des oiseaux.
Que nous apportent les récentes découvertes chinoises exposées au Musée de Londres? (en 2002/2003)
Les dinosaures à plumes de Liaoning en Chine
- PROTARCHEOPTERYX ROBUSTA, proche parent des DROMAEOSAURES. Un exemplaire unique et incomplet décrit en 1997, théropode coureur à longues pattes, à plumes de queue symétriques, avec des mains et poignets agiles, des griffes acérées. - CAUDIPTERYX ZOUI (avec des gastrolites dans l’estomac, et la queue en éventail) proche d’ - ARCHEOPTERYX et SINOSAUROPTERYX (un adulte de 150cm et un juvénile de 55cm au duvet dorsal). Ce sont deux dinosaures emplumés âgés de 145MA, avec des plumes symétriques ne permettant pas le vol mais ils sont postérieurs de 5MA à l’archéoptéryx et ils n’ont pas de plumes modernes, et selon Eric Buffetaut il faudrait trouver un dinosaure à plumes modernes…
C’est fait ! Avec SINORNITHOSAURUS (124 à 147MA) dit " le raptor frisé ", plus un "non nommé" appartenant aux dromaeosauridés. Ces nouveaux dromaeosaures ont des plumes à filament central à leur base rappelant les arêtes semblables aux plumes d’oiseaux actuels avec un rachis central et des barbes en série. Découvert en 2000 ce dromaeosaure long de 1,25m pesant 6Kg pouvait replier le bras comme une aile grâce à un os en demie lune.
CONSÉQUENCE :
Les plumes ont précédé l’origine des oiseaux et du vol !
Quel est donc le rôle des plumes ?
Parade sexuelle, reconnaissance sociale ou spécifique, isolation thermique, les trois certainement.
Coelurosaures ou dromaeosaures, de toutes façons, certains dinosaures sont à l’origine des oiseaux. Certains dinosaures n’ont pas disparu et selon Philippe Curie du Tyrel Muséum dans l’Alberta (Canada) : "cela montre que les dinosaures ne sont pas disparus mais sont bien représentés par 10 000 espèces d’oiseaux".
Et si vous n’êtes pas encore convaincu, voyez donc ces curieux oiseaux chinois possédant encore quelques archaïsmes, bien qu’étant étonnamment modernes, comme :
- Confuciusornis (1994), à bec édenté dont on distingue le mâle par les immenses plumes caudales tel un coq faisan vieux de 120 MA, possédant un pygostyle et un bréchet, d’une envergure de 36cm pour une longueur de 20cm, par centaines d’exemplaires.
- Chaoyangia (137 à 142 MA), de la taille d’un gros moineau, avec bréchet développé, pygostyle, et furcula en V, du début du crétacé inférieur, qui n’était pas exposé ni mentionné à Londres en 2002/2003 à l’exposition "Dinobirds".
- Changchengornis, muni d’une corne sur le bec comme un calao.
- Liaoxiornis, 122MA est le plus petit oiseau fossile connu dans cette formation de Liaoning.
Les deux dernières nouveautés :
- Jeholornis, un oiseau à corps de dromaeosaure de 120MA, volant et grimpant, avec 50 graines dans l’estomac ;
- Shenzhouraptor sinensis, qui serait le chaînon entre théropodes et oiseaux daté de 140 MA seulement (mais pas encore de publication donc méfiance…).
La prudence s’impose, témoin cette chimère made in China et vendue fort cher aux USA: l’archéoraptor liaoningensis, fabriqué à partir de deux fossiles distincts : une aile de Ianornis martini et une queue de dromaeosaure.
Enfin il est fait mention d’un véritable oiseau espagnol : Eoalulavis hoyasi, vieux de 135MA, découvert en juin 1996, son aile est munie d’alules lui permettant de ne pas décrocher en vol, indispensable pour le vol lent et pour manœuvrer, très moderne par son aile sa fourchette, avec hypocléide, son pygostyle son sternum ossifié, mais primitif par son bec denté.
Même dans le livre la classification phylogénétique du vivant, on n’arrive plus à distinguer les dinosaures des oiseaux, ces derniers en étant !
Cependant j’ai cherché et trouvé des distinctions propres aux oiseaux :
- fenêtre antéorbitaire fusionnée dans l’orbite
- tête globuleuse et gros cerveau
- des os pneumatiques
- un ilion plus long et fixé sur de nombreuses vertèbres
- un pubis orienté nettement vers l’arrière, parallèlement à l’ischion et soudé à celui-ci
- une queue courte avec pygostyle (croupion - vertèbres soudées)
- un bréchet très développé
- des os coracoïdes longs
- une fourchette en V avec hypocléide
- une aile faite pour le vol battu
- des doigts 2/3/4 réduits et soudés
Mais dites-vous bien comme Bob Baker que "dans votre canari, il y a une parcelle de T-Rex qui sommeille" et comme Marc Norell du musée de New York : "les mots dinosaure et oiseau sont interchangeables et synonymes. Ils caquettent, roucoulent, sont dans nos poulaillers, sur nos places bref ils sont encore parmi nous et partout !".
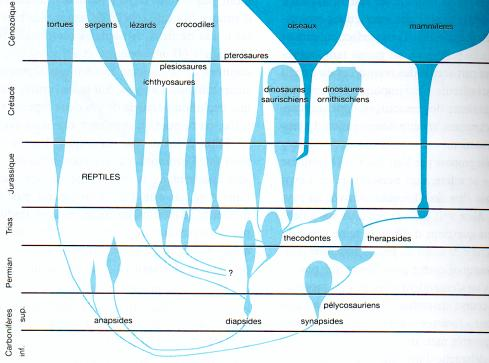
Phylogénie "classique" des reptiles, des oiseaux et des mammifères,
montrant l’extinction des ichtyosaures, des plésiosaures, des ptérosaures
et des dinosaures ( plus soudaine mais déjà bien amorcée ).
3.2.6.3 Classification
a. Les Ratites.
Ils n’ont pas de capacité au vol. Leur bréchet est absent : pas de muscles pour le vol. Ce sont des oiseaux coureurs (Autruches, Namdoux, Emeux, Kiwi).
b. Les Carinates.
Ils ont l’aptitude au vol et un bréchet bien développé
c . Les Impennés
Oiseaux aquatiques ( ailes nageoires ), plumes non différenciées, quatre orteils palmés, ce sont les Manchots.
Date de dernière mise à jour : 30/03/2018
Ajouter un commentaire