

Insectes 6
5.5 Mue
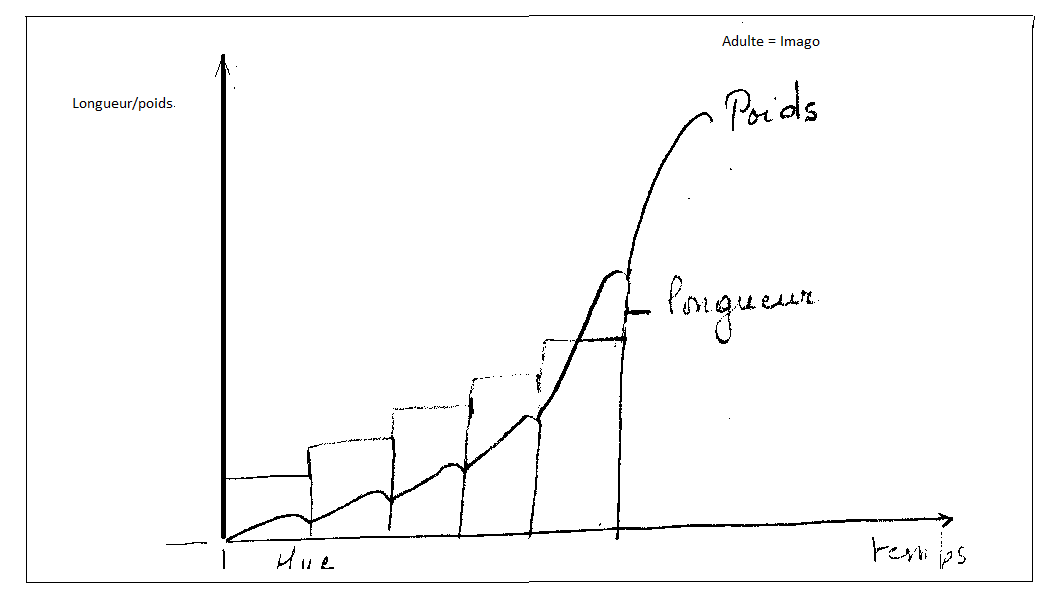
Une fois sorti de l’œuf, le jeune insecte va se développer, mais, du fait de l’existence d’un tégument chitineux inextensible, la croissance est discontinue : elle se fait par des mues successives .
Pour chaque espèce, le nombre de mue est fixe ou presque fixe.
Chaque stade est séparé du suivant par une mue , la mue après laquelle l’insecte est sexuellement mûr s’appelle mue imaginale, cette dernière mue fait apparaître l’insecte adulte ou imago.
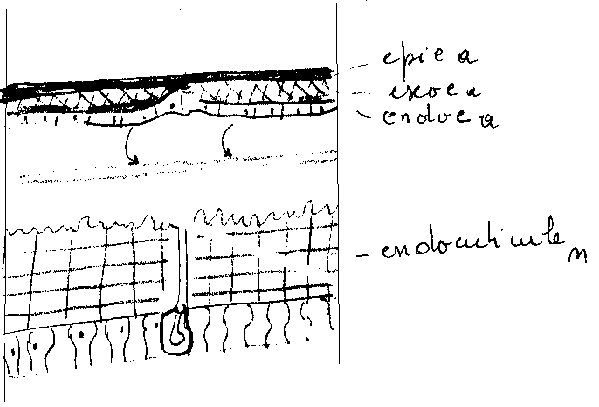
Le renouvellement périodique de la cuticule ou exuviation, est l’aboutissement d’un processus physiologique complexe qui affecte l’ensemble des formations ectodermiques , où les cellules épidermiques jouent un rôle essentiel.
En dehors des périodes de mue, les cellules épidermiques inactives ont une forme approximativement cubique . Elles sont étroitement imbriquées et collées. Les organites cellulaires ne présentent aucune répartition particulière.
Le déroulement de la mue peut être scindé en quatre étapes que nous examinerons plus loin.
Remarque : Les insectes ont une croissance définie, ce qui les différencie des Crustacés qui ont une croissance indéfinie , ces animaux grandissant et muant quasiment pendant toute leur vie.
5.5.1 Le mécanisme de la mue
Le renouvellement de la cuticule est assuré par l’épiderme dont l’activité est alors remarquable.
5.5.1.1 Changements au niveau de l’épiderme

Intense activité mitotique ( Ex : chez Rhodnius, le nombre de cellules épidermiques passe de 2,5 à 9 mm2 en trois jours ). Ces cellules épidermiques acquièrent des caractéristiques ultra structurales liées à leur activité synthétique ( appareil de Golgi très développé, mitochondries plus nombreuses…). L’épiderme prend un aspect plissé aidant le décollement , il correspond ainsi, par ses dimensions , au stade suivant.
Les cellules épidermiques sont remaniées, leur hauteur augmente , avec élargissement des espaces intercellulaires . Les cellules acquièrent dans leur partie apicale, des villosités.
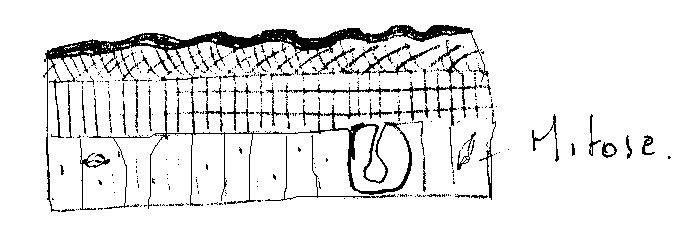
5.5.1.2 Décollement de la cuticule ou apolyse
Le changement de forme des cellules épidermiques induit leur décollement de la cuticule . Leurs liaisons avec la cuticule se rompent au niveau des canalicules,des pores , des canaux des glandes dermiques, des dendrites et des cellules sensorielles.
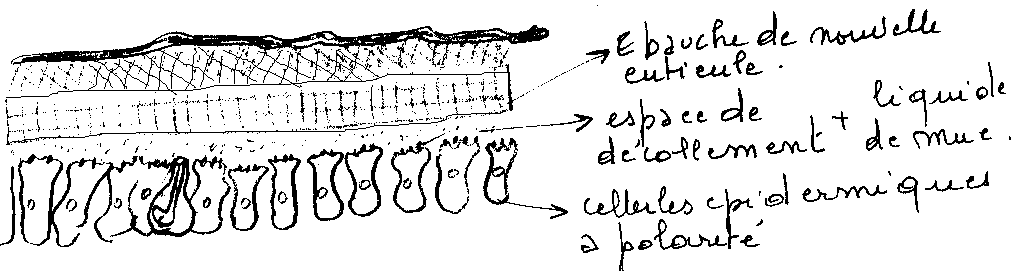
Dès l’apolyse, un liquide de mue apparaît à l’état de gel dans l’espace ainsi créé. Ce liquide exuvial contient divers enzymes ( chitinase , chitibiase, protéase…). Mais inactifs à ce stade.
5.5.1.3 Sécrétion des couches cuticulaires préexuviales
Les cellules épidermiques ne restent à nu que peu de temps , car rapidement, la cuticuline est élaborée. L’épicuticuline est secrétée immédiatement après, d’une manière uniforme , la procuticuline ( chitine et protéine ) l’est ensuite.
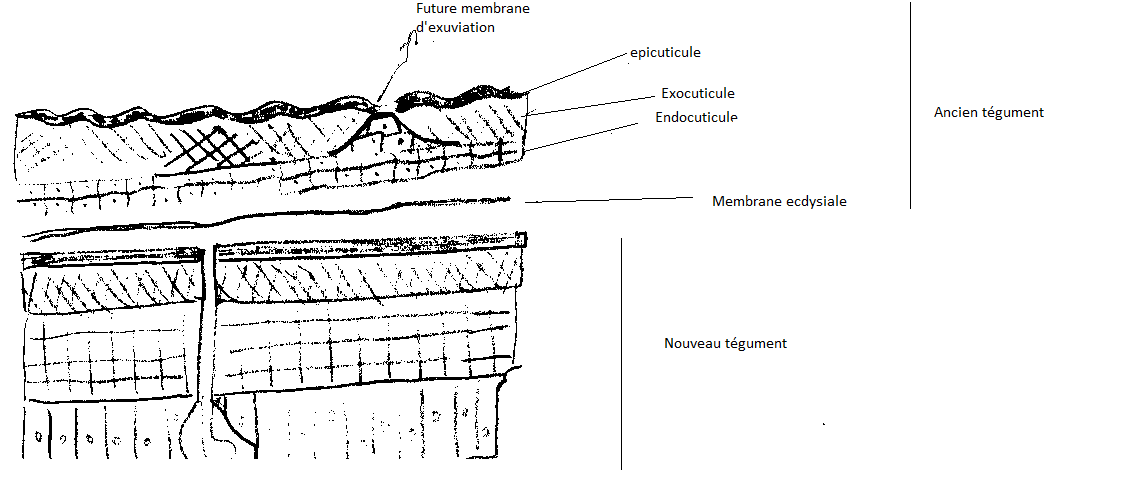
5.5.1.4 Résorption de l’ancienne cuticule
L’activation du liquide de mue coïncide avec le dépôt de l’épicuticule interne , ce liquide détruit toute l’endocuticule à l’exception de sa couche la plus profonde ( vers l’extérieur) qui se modifie et persiste comme mebrane exuviale L’exocuticule n’est pas affectée.
Les produits de l’histolyse de l’endocuticule sont récupéré par l’insecte.
5.5.2 Exuviation
Après action du liquide de mue, l’ancienne cuticule n’est plus composée que d’épicuticule et d’exocuticule . Des lignes exuviales sont présentes sur la tête de la larve et représentent des lignes de moindre résistance le long desquelles la cuticule peut se rompre facilement sous l’action d’une pression.
Au moment du rejet de l’ancienne cuticule, l’insecte avale de l’air ou de l’eau s’il est aquatique , de sorte que la pression de l’hémolymphe augmente après gonflement du tube digestif.
Une fois l’exuvie fendue, l’insecte s’en extrait, la tête et le thorax en premier suivi par l’abdomen et les appendices .
L’exuvie est formée non seulement par les restes de l’ancienne cuticule mais aussi par l’intima des trachées, de l’intestin postérieur et intérieur et des canaux évacuateurs des diverses glandes.
5.5.3 Evolution post exuviale
Au moment de l’exuviation, la nouvelle cuticule est molle et souple : elle le reste quelque instants . L’insecte avalant de l’air augmente la pression de l’hémolymphe et ainsi provoque le déplissement et l’extension du tégument. Très rapidement, le tannage de la cuticule entraîne l’inextensibilité et le durcissement tégumentaire.
Remarque : La mue n’est pas un phénomène d’excrétion, puisqu’une partie de l’ancien tégument est utilisée à nouveau.
5.6 La nymphose
La larve des insectes holométaboles arrivée au terme de sa croissance ne s’alimente plus, vide son intestin et s’immobilise. Généralement elle changera d’habitat et réagira différemment aux facteurs externes.
On parlera alors de nymphe , celle-ci acquiert les caractéristiques externes de l’imago ; elle possède des yeux composés , des antennes , des appendices et des fourreaux alaires ; les organes génitaux sont présents, mais en général réduits.
5.6.1 Les nymphes.
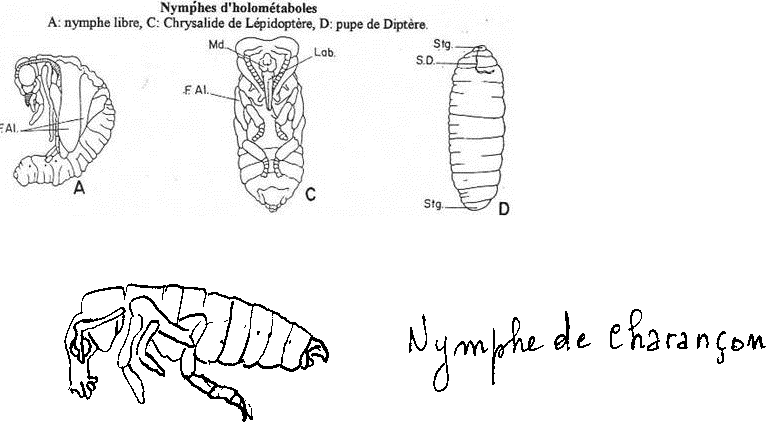
La nymphe est partiellement ou totalement immobile. L’organisation de la forme de la larve est fortement remaniée en vue de la transition vers la phase adulte.
a\ La nymphe libre (nue).
Les nymphes de ce type sont blanchâtres, immobiles et possèdent des appendices libres mais immobiles. On les trouve chez les coléoptères et typiquement chez Ténébrio.
b\ La chrysalide ou nymphes momies.
Ces nymphes sont enfermées dans un cocon de soie. C’est le cas des papillons (lépidoptères). La nymphe possède des appendices qui sont collés au corps.
c\ La pupe des Diptères.
Les pupes sont totalement enfermées dans la dernière exuvie de la dernière larve vermiforme. Cette larve est totalement immobile, de couleur foncée.
Dans tous les cas, la nymphe a comme l’adulte des yeux composés, des antennes, des appendices et des fourreaux alaires (replis de tégument enfermant les ailes).
La dernière mue va libérer l’adulte. Cette métamorphose débute avec la mue nymphale et se poursuit pendant tout le stade nymphal et se termine avec la mue imaginale. Durant cette période, l’insecte subit des phénomènes d’histogenèse, d’histolyse et de remaniements.
5.7 Le mécanisme de la métamorphose
Pendant le développement embryonnaire, des amas de cellules non différenciées forment des disques imaginaux et des éléments diffus (histoblastes). Au cours de la vie larvaire, les disques sont stationnaires (amas blanchâtres) et seront à l’origine des organes spécifiques à l’adulte. Ces disques s’accroissent et poursuivent leur évolution après la mue de la nymphe.
Les mécanismes de la métamorphose débutent avec la mue nymphale, se poursuivent durant toute la phase nymphale et se terminent par la mue imaginale.
La métamorphose est caractérisée par des phénomènes d’histogenèse ( développement des organes propres) et d’histolyse ( destruction des organes larvaires) et de phénomènes de remaniement qui affectent les organes communs à la larve et à l’imago.
Lorsque la métamorphose extérieure est considérable, elle s'accompagne de remarquables phénomènes internes. Dans la Chrysalide, les tissus sont réduits en une bouillie où se résolvent tous les organes larvaires; ce retour à l'état embryonnaire est nommé histolyse. Lorsque l'histolyse est particulièrement rapide (Diptères), les leucocytes ou phagocytes semblent dévorer les tissus et en accumuler la substance pour refaire des tissus nouveaux (Canin, 1876; Viallanes, 1882; Kowalesky, 1887). L'histolyse du système musculaire peut avoir lieu :
1° par prolifération des noyaux qui deviennent sphériques, s'entourent d'une aire protoplasmique et enrichissent la masse contractile, s'en nourrissent puis se répandent dans la cavité, analogues à des granules vitellins (Viallanes). Kowalesky, d'autre part, sur des pupes de Muscides, a vu les muscles pénétrés par les leucocytes, découpés en îlots, et finalement englobés sous forme de boules à noyaux vingt-quatre heures ont suffi à cela;
2° par dégénérescence et mort du noyau musculaire qui se vide, et reste semblable à une petite coque vide, tandis que la substance contractile disparaît.
Le corps adipeux rentre en histolyse par formation dans ses cellules de granules nombreux, que la rupture de la membrane met en liberté. Les grandes boules à noyaux seraient les cellules du corps adipeux hypertrophiés, contenant de nombreux granules, dont chacun représenterait une cellule embryonnaire.
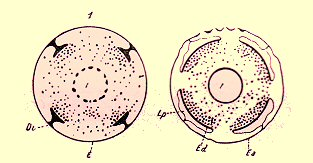
Disques imaginaux de nymphes d'insectes. 1. Naissance des disques Di au dépens de l'ectoderme E; I. intestin larvaire en histolyse. - 2. Ed, nouvel ectoderme des disques, remplaçant l'ectoderme ancien Ea; I, intestin reconstitué; lp, lame provisoire.
Les éléments cellulaires des trachées et des glandes salivaires, lors de la transformation, prennent un surcroît d'activité qui donne à ces organes l'aspect d'un tube à parois épaisses formées de cellules embryonnaires; à ce moment les cellules se séparent, l'organe larvaire disparaît; il y a donc mise en circulation dans la cavité générale de la nymphe de très nombreux éléments embryonnaires. Le système nerveux est, lui aussi, profondément remanié. L'histolyse est suivie d'une reconstruction des tissus, ou histogenèse.
Sur chaque anneau en quatre points (deux supérieurs et deux inférieurs), l'hypoderme s'épaissit, par production de nombreuses cellules embryonnaires. Ces épaississements sont les disques imaginaux. Par une sorte de clivage, leur zone externe, formée en définitive de cellules hypoderdermiques larvaires n'ayant pas pris part à leur formation, et situées en dehors, leur zone externe fort mince (lame provisoire) dégénère et disparaît.
Le disque proprement dit constitué par dés éléments ectodermiques et mésodermiques (globules du sang) se sépare en deux couches : l'exoderme ou hypoderme de l'Imago, et le mésoderme des tissus profonds. Enfin, les cellules hypodermiques larvaires non transformées ayant disparu entre les disques, ceux-ci s'accroissent par leurs bords, se soudent, limitent la cavité du corps et constituent l'hypoderme de l'Imago.
Les éléments cellulaires des trachées et des glandes salivaires, lors de la transformation, prennent un surcroît d'activité qui donne à ces organes l'aspect d'un tube à parois épaisses formées de cellules embryonnaires; à ce moment les cellules se séparent, l'organe larvaire disparaît; il y a donc mise en circulation dans la cavité générale de la nymphe de très nombreux éléments embryonnaires. Le système nerveux est, lui aussi, profondément remanié. L'histolyse est suivie d'une reconstruction des tissus, ou histogenèse.
Sur chaque anneau en quatre points (deux supérieurs et deux inférieurs), l'hypoderme s'épaissit, par production de nombreuses cellules embryonnaires. Ces épaississements sont les disques imaginaux. Par une sorte de clivage, leur zone externe, formée en définitive de cellules hypoderdermiques larvaires n'ayant pas pris part à leur formation, et situées en dehors, leur zone externe fort mince (lame provisoire) dégénère et disparaît.
Le disque proprement dit constitué par dés éléments ectodermiques et mésodermiques (globules du sang) se sépare en deux couches : l'exoderme ou hypoderme de l'Imago, et le mésoderme des tissus profonds. Enfin, les cellules hypodermiques larvaires non transformées ayant disparu entre les disques, ceux-ci s'accroissent par leurs bords, se soudent, limitent la cavité du corps et constituent l'hypoderme de l'Imago.
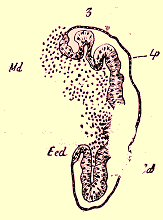
Disque imaginal de l'aile d'une Tipule. Ecd, Ectoderme du disque; Md, son mésoderme; Cd, cavité du disque; lp lame provisoire.
Dans la tête et le thorax, les disques sont formés bien avant la métamorphose; dans l'abdomen, c'est à ce moment même. C'est donc par un mécanisme analogue à celui du renouvellement des dents chez les Mammifères que l'hypoderme larvaire est remplacé chez les Holométaboliens par un hypoderme à structure définitive (Viallanes).
Tous les organes (tube digestif, trachées) vont être édifiés à nouveau par des processus semblables. Les yeux, dits composés, de l'Imago s'organisent au dépens des cellules optogènes et des fibres postrétiniennes de la nymphe (la larve possédait de nombreux ocelles).
En résumé, le développement des Insectes peut se faire : 1° sans histolyse ni histogenèse (Amétaboliens); 2° avec histogenèse sans histolyse (Corethra, Diptère Némocère) (Weissmann); 3° enfin avec histogenèse précédée d'histolyse. Tels sont les Insectes à métamorphoses, au sens le plus restreint; encore cette histolyse peut-elle être simple, ou accompagnée de phagocytose, au moins pour le tissu musculaire (Muscides, d'après Kowalesky). (Maurice Maindron).
Des remaniements affectent les organes communs : le cœur, certains muscles, les tubes de Malpighi (à fonction excrétrice). Les cellules subissent une dé-différenciation puis acquièrent les caractères imaginaux sans changer de destinée.
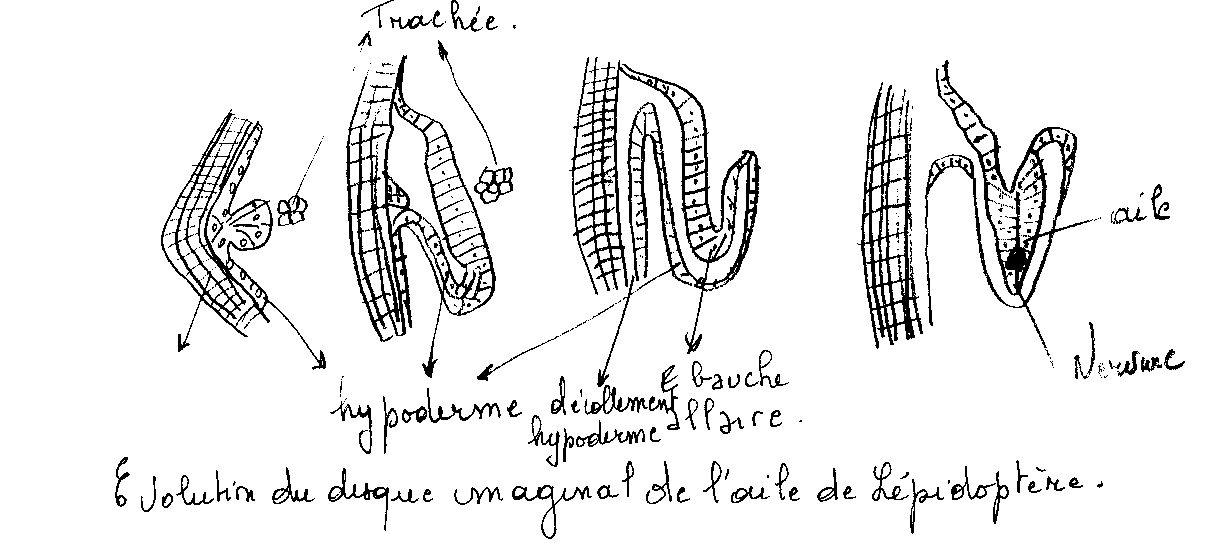
Ex : les ailes
L’hypoderme se décolle du tégument au voisinage d’une trachée . Il se forme une anse qui s’invagine au sommet . Dans cette invagination pénètrent des éléments mésenchymateux et une ramification de la trachée qui elle même se divisera .
A la mue nymphale, du fait de la multiplication des cellules de l’ébauche , une évagination déjà commencée antérieurement s’accentue et il en résulte la saillie à l’extérieur du disque , devenu ébauche alaire.
5.8 Le déterminisme de la métamorphose
- Les mues larvaires sont conditionnées par l’activité du cerveau et d’une glande prothoracique ; en même temps, les corpora allata secrètent une hormone qui s’oppose au développement des ébauches d’imago : l’hormone juvénile ou néoténine .
- La mue nymphale et la mue imaginale dépendent toujours de l’action combinée du cerveau et d’une glande prothoracique , mais sont rendue possible du fait de l’arrêt de la sécrétion de la néoténine. Cet arrêt doit être commandé par le cerveau.
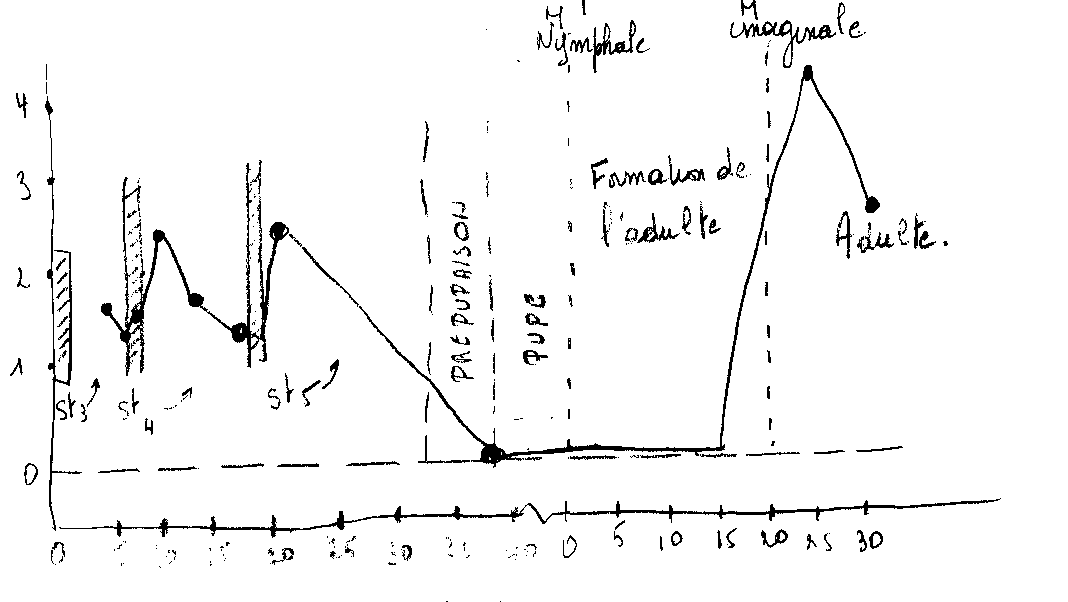
Evolution du taux de J.H au cours des mues
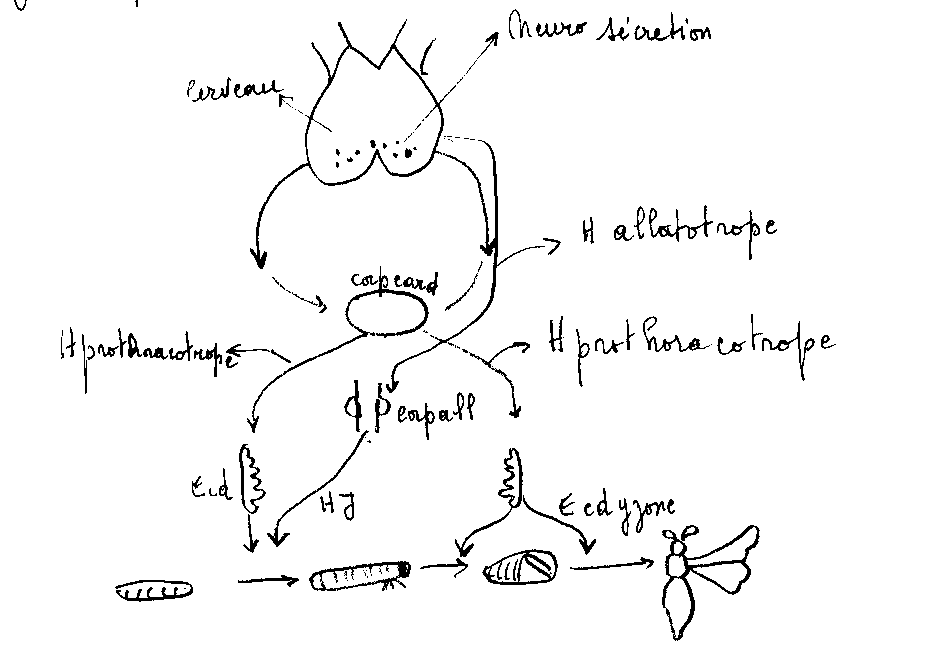
La neurosécrétion du protocérébron ou hormone prothoracotrope agirait, par l’intermédiaire des corps cardiaques sur la glande prothoracique qui libère l’hormone de mue ou ecdysone.
5.9 Analyse fonctionnelle des cycles de développement.
Principe : On peut reconnaître, pendant le même cycle développement, plusieurs fonctions remplies par divers stades de développement (stades pré-adultes et adultes). On distingue différentes fonctions : une fonction de développement et de différenciation (larves) ; une fonction d’acquisition de la nourriture ; une fonction de dispersion ; une fonction d’accouplement ; une fonction de distribution de la nourriture à la progéniture ; une fonction de sélection des sites de croissance ; une fonction de ponte.
On reconnaît cinq cycles fondamentaux chez les arthropodes.
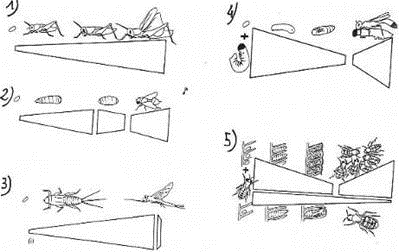
La largeur de la bande représente la quantité de ressources disponibles. Une augmentation de cet largeur montre une acquisition de cette nourriture alors qu’une diminution de cette même bande indique une utilisation de ces ressources.
En 1, on voit une transition graduelle vers l’adulte. La larve et l’adulte sont impliqués dans l’exploitation des mêmes ressources. Les adultes ont en plus les fonctions d’accouplement, de ponte et de dissémination.
En 2, on voit que les larves et les adultes se nourrissent en exploitant des ressources différentes. La larve se nourrit avec des cadavres d’animaux morts : c’est une fonction de développement et d’acquisition des ressources. On obtiendra alors une pupe qui va utiliser les ressources acquises par la larve pour son développement et sa différenciation. L’adulte va continuer à se nourrir mais à partir d’une autre source de nourriture. Ce dernier possède les fonctions d’accouplement et de sélection du site de développement des larves, de ponte.
En 3, on observe les insectes amphibiontiques (à larves aquatiques et à adultes aériens). Il y a séparation du rôle d’acquisition et de dispersion d’une manière très marquée. L’adulte ne se nourrit pas ou seulement très peu. C’est le cas des éphémères, des trichoptères et des plécoptères.
La larve est herbivore ou carnivore. Sa durée de vie varie entre quelques mois et trois ans. Elle possède la fonction de nutrition. L’adulte ne vit que quelques heures pendant lesquelles il doit s’accoupler puis pondre dans un site convenable aux larves. C’est ensuite sa mort.
En 4 et 5, on voit des adultes de certaines espèces qui remplissent les fonctions de dispersion, de ponte mais aussi de stockage et de mise à disposition des ressources sous formes provisions pour les larves.
En 4, c’est le cas des guêpes solitaires où l’adulte pond un œuf sur une proie paralysée. Cet adulte a alors remplie son rôle de dispersion, d’accouplement de sélection du cycle de ponte et de ponte. La larve doit acquérir la nourriture et la fonction de développement.
En 5, on observe les hyménoptères sociaux. Il y a un partage des fonctions assuré par une différenciation des individus en castes. Les larves utilisent les ressources mises à leur disposition pour leur croissance. Les ouvrières sont stériles et ont la fonction d’acquisition des ressources. La reine est un adulte fertile regroupant les fonctions d’accouplement, de ponte et de dispersion. Les males sont des adultes fertiles avec la fonction d’accouplement mais n’ont aucun rôle dans l’acquisition.
Date de dernière mise à jour : 12/01/2019
Ajouter un commentaire


