

Insectes 3
1.4.4 Le système nerveux
1.4.4.1 Généralités
Comme chez les Vertébrés, l’élément de base du système nerveux de l’Insecte est le neurone avec dendrites et axone.
L’influx nerveux est transmis également par un médiateur chimique, l’acétylcholine.
La cholinestérase, enzyme inhibitrice de cet influx est également présente.
1.4.4.2 Données morphologiques
- système nerveux typique d’un Arthropode.
- Plan annélidien, primitivement autant de paires de ganglions que de métamères.
- Courte commissure transversale entre les deux ganglions d’une paire
- Ganglions de métamères voisins reliés par des connectifs longitudinaux.
Cette disposition primitive est généralement altérée et peu reconnaissable :
- Raccourcissement des commissures voire coalescence de ganglions d’une paire.
- Accolement , voire fusion de connectifs longitudinaux.
- Condensation en de volumineux amas nerveux de ganglions appartenant à des métamères associés dans un même tagme : tête, thorax, région ano-génitale.
Le système nerveux des insectes est très développé, il peut être divisé en trois grandes parties :
- Le système nerveux central
- Le système nerveux sympathique
- Le système nerveux périphérique
1.4.4.3 Le système nerveux central
Le système nerveux central est formé de deux ganglions par segment, reliés entre eux par des commissures transversales et aux autres paires de ganglions par des connectifs longitudinaux. Il s’est différencié sur ce type de base et comporte :
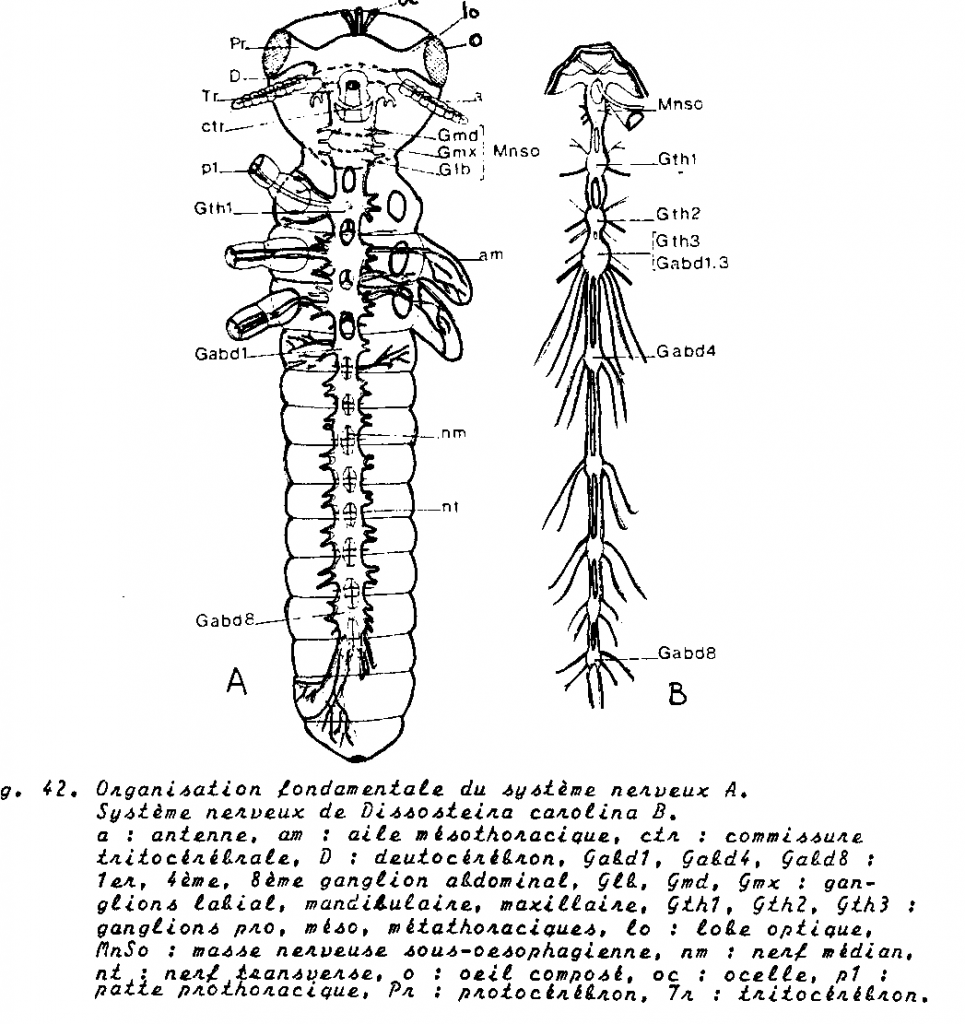
1.4.4.3.1 Le cerveau
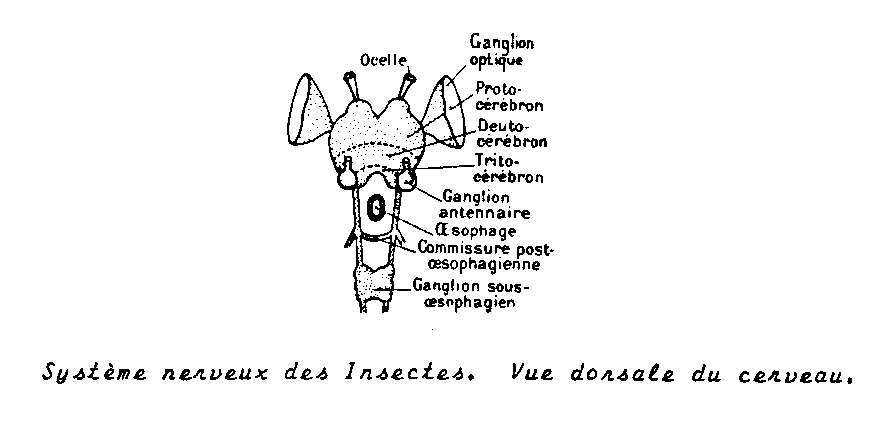
ganglions cérébroïdes, structure typique des arthropodes à savoir
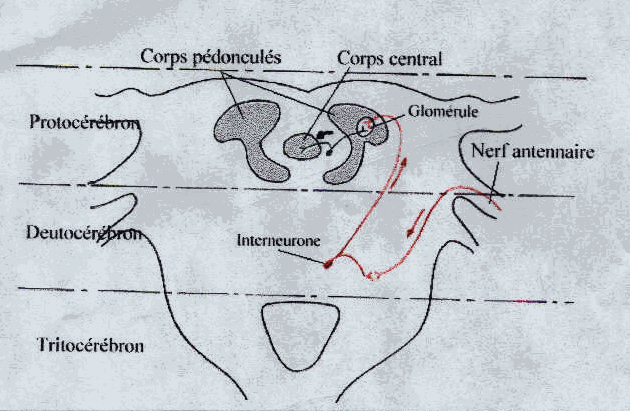
- Protocérébron ou cerveau antérieur avec corps pédonculés qui sont des centres d’association . En rapport avec les centres optiques innervant les yeux composés.
- Deutocérébron ou cerveau moyen : comporte dux lobes antennaires d’où se détachent deux volumineux nerfs antennaires symétriques
- Tritocérébron ou cerveau postérieur constitué par deux masses nerveuses coniques , séparées par l’œsophage , en relation avec le deutocérébron.
1.4.4.3.2 Amas sous oesophagien
constitué par la condensation de trois paires de ganglions qui innervent mandibules, maxilles et labium, ainsi que la musculature et les téguments post céphaliques.
1.4.4.3.3 Chaîne nerveuse ventrale métamérisée.
Trois paires de ganglions thoraciques innervent les pattes et les ailes.
Onze paires de ganglions abdominaux ( en principe), mais on en dénombre souvent que 8. Le dernier ganglion visible dit ganglion génital résulte de la coalescence des 8,9,10 et onzième paires et innerve les gonopodes, les pièces anales, les cerques, le rectum et les conduits génitaux.
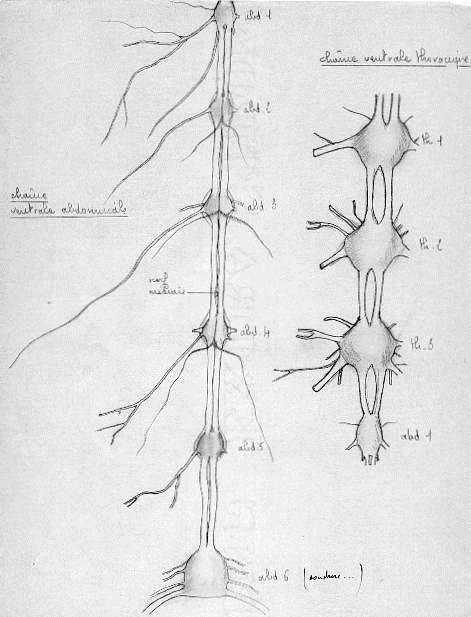
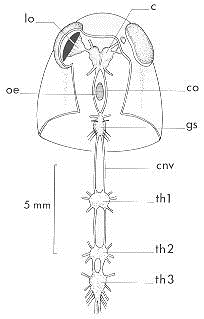
Chaque ganglion constitue le centre moteur et sensoriel du segment correspondant
Schéma du système nerveux central de Locusta migratoria migratorioides en vue dorsale jusqu'au 3e gangilon thoracique (d'après E.T. BURTT & W.T. CATTON, 1959)
c : cerveau, cnv : chaîne nerveuse ventrale, co : connection péri-oesophagienne, gs : ganglion sous-oesophagien, lo : lobe optique, oe : oesophage, th1-2-3 : 1er, 2e et 3e ganglions thoraciques
1.4.4.4 Le système nerveux symphatique
Le système nerveux central est en relation avec le système nerveux sympathique qui innerve les principaux viscères. Il préside à la vie végétative et peut se subdiviser en :
- Système stomatogastrique en relation avec le tritocérébron, il se trouve en position dorsale sur l’intestin antérieur. Il innerve l’intestin antérieur et moyen et aussi le cœur.
- Système sympathique ventral , ce sont les nerfs ventraux associés avec les ganglions de la chaîne ventrale, ils innervent les stigmates.
- Système sympathique caudal , innerve les organes reproducteurs et l’intestin postérieur. Ces nerfs sont connectés avec les derniers ganglions de la chaîne ventrale.
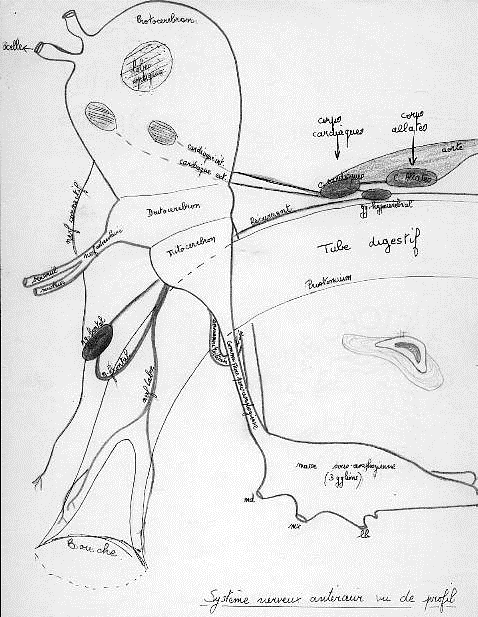
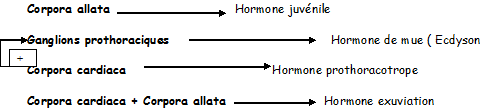
Au système nerveux symphatique sont annexés des organes du système endocrinien : c’est le système endocrine rétrocérébral , comportant :
Les corpora cardiaca et corpora allata , c’est le système endocrine rétrocérébral
1) Les corpora cardiaca : organes symétriques annexés aux parois latérales de l’aorte. Leurs rôles sont les suivants :
Schéma du système neuroendocrinien de la tete de Locusta migratoria en vue latérale (d'après G.B. STAAL, 1961 in B.P. UVAROV, 1956)
ao : aorte, b : bouche, ca : corpora allata, cc : corpora cardiaca, cn : cellules neurosécrétrices du cerveau, gc : ganglion hypocérébral, gf : ganglion frontal, gp : glande prothoracique, gs : ganglion sous-oesophagien, m : membrane du cou, nc : nerf des corpora cardiaca, nca : nerf des corpora allata, no : nerf oesophagien externe, nr : nerf récurrent, o : ocelle, oc: oeil composé, oe : oesophage

a) Assurent le rejet dans l’hémolymphe des neurosécrétions élaborées par les cellules neurosécrétrices réparties dans les différents lobes du cerveau.
b) Organes endocrines qui contrôlent le rythme cardiaque ,…péristatilsme intestinal et rectal, lipémie, protéinémie et croissance.
2) Les corpora allata sont deux glandes parenchymateuses ovoïdes situées de part et d’autre de l’œsophage. Elles élaborent l’hormone juvénile qui assure , au cours du développement postembryonnaire , le maintien des caractères larvaires, et, au cours de la vie imaginale, la vitellogénèse et l’activité des glandes annexées à l’appareil génital.


1.4.4.5 Le système nerveux périphérique
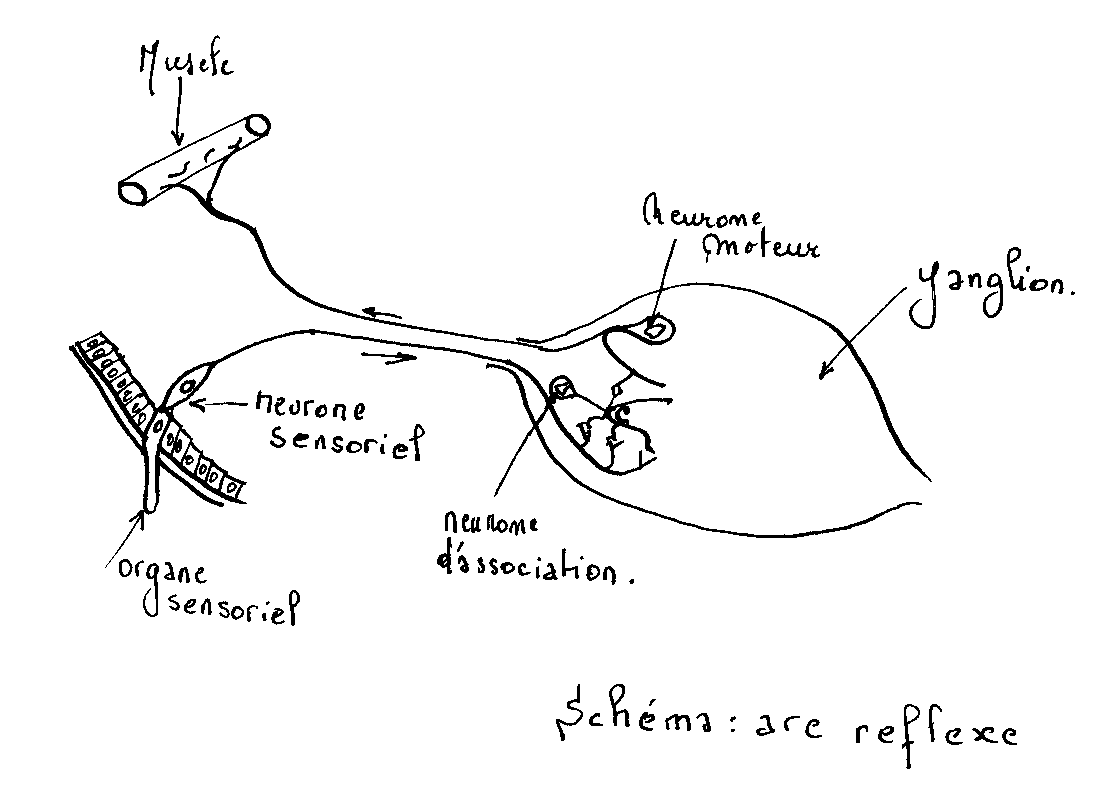
Ce système comprend tous les nerfs irradiant les ganglions des systèmes nerveux central et sympathique . Chaque ganglion ventral possède des neurones périphériques soit moteurs et en position dorsale, soit d’association et en position ventrale
Le type normal de réflexe fait intervenir au moins trois neurones : Un neurone sensitif périphérique, un neurone d’association, et finalement, un neurone moteur.
Remarque : le système nerveux tel que nous venons de le décrire peut subir des modifications dans le sens de la condensation des ganglions.
Dans un premier stade, les trois paires de ganglions lymphatiques se sont réunies en une seule masse dans le mésothorax. Dans un deuxième stade, toute la chaîne ventrale s’est condensée en une seule masse, toujours dans le mésothorax. C’est le cas de la mouche domestique.
Notons encore, qu’avec les Mollusques Céphalopodes, les Insectes sont les Invertébrés qui comportent le système nerveux le plus compliqué. Chez eux, le rôle des réflexe est très important, il réduit celui des ganglions cérébroïdes. Chaque ganglion de la chaîne ventrale comporte à la fois des nerfs sensitifs et moteurs et offre de ce fait une autonomie relative. Il existe bien sûr un coordination contrôlée par le cerveau.
Les réflexes suffisent à assurer la plupart des mouvements nécessaires à la vie de l’Insecte.
Chez les Insectes, pas de neurones moteurs dans le "cerveau ( ils sont dans la chaîne ventrale) et pas de neurones sensoriels dans le système nerveux central
Le nombre de neurones est faible dans le système nerveux central (Oeil d'Odonate : 210 000 neurones; Homme:400 000 000)
1.4.5 . le sens chez l’Insecte
1.4.5.1 le sens tactile
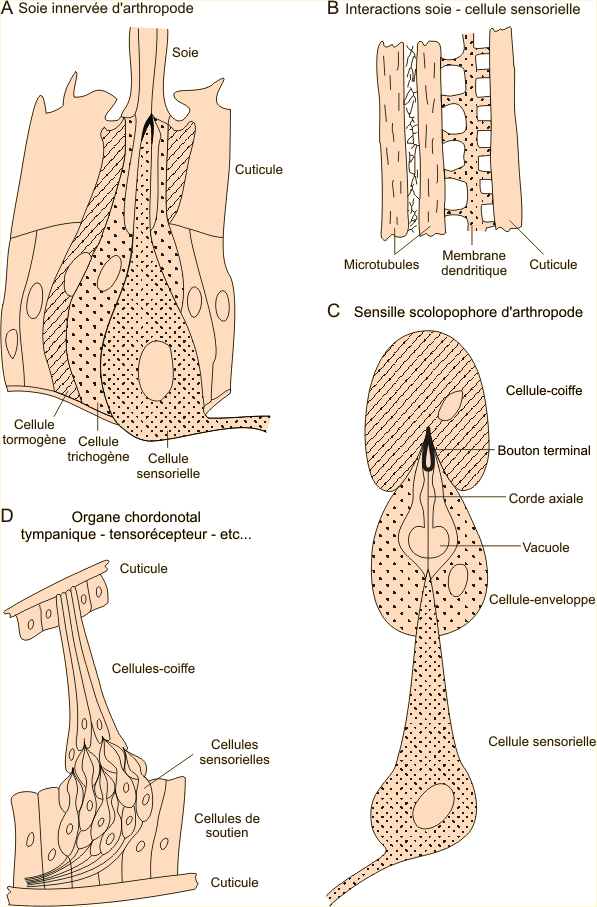
Nombreux sont les organes des sens chez l’insecte, le type le plus simple est le poil sensoriel , Il constitue d’ailleurs la structure de base de beaucoup d’autres organes des sens.
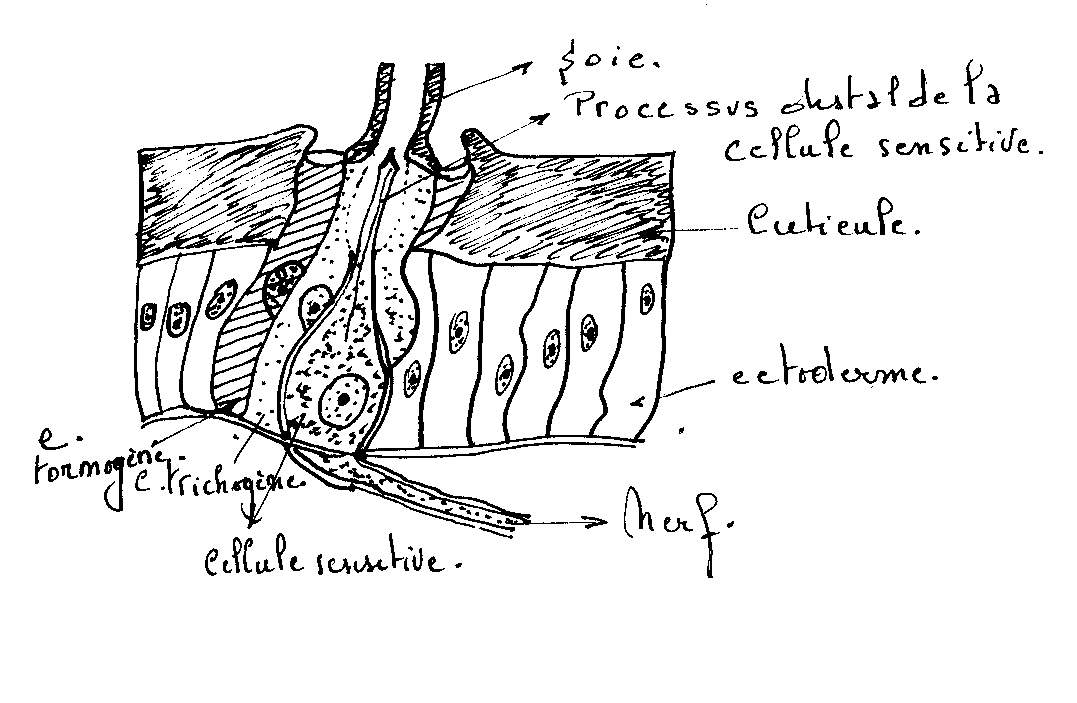
Le poil sensoriel comporte une structure sensorielle ( une ou plusieurs cellules) dont l’extrémité dendritique pénètre dans le canal cuticulaire de la soie sécrétée par une cellule annexe ou cellule trichogène et relié au tégument par une membrane articulaire élaborée par une autre cellule annexe ou cellule tormogène.
détails et localisation d’organes sensoriels tactiles
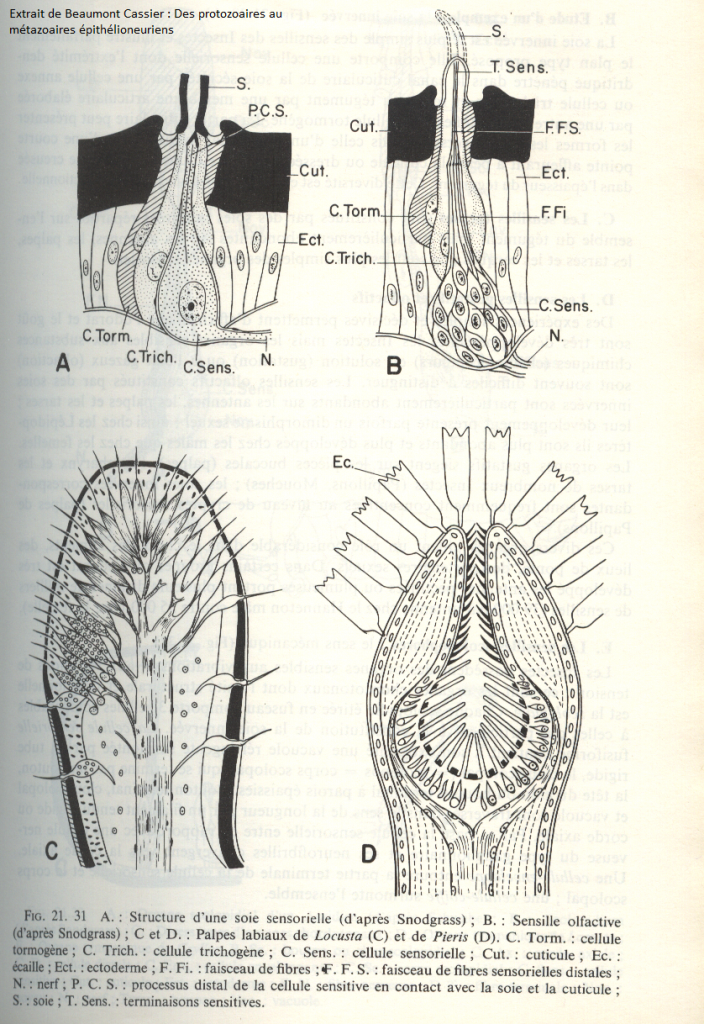
Les extrémités dendritiques se terminent dans le canal cuticulaire au voisinage immédiat de la base de la soie par une zone riche en microtubules associés à la membrane par des invaginations coniques de celle-ci (B) La membrane est par ailleurs étroitement associée à la base de la soie de sorte que tout mouvement de celle-ci provoque une distension dans les invaginations coniques fixées au réseau de microtubules peu mobile. Ce serait ces mouvements qui induiraient l'ouverture des canaux cationiques responsables de la formation d'un potentiel générateur.Le toucher est exercé par les poils sensoriels qui sont localisés en divers endroits du corps et aussi sur les antennes , d’autres poils sensoriels , plus complexes fonctionnent comme des récepteurs de tension . Ils forment alors des organes chordotonaux distribués sur les antennes, les palpes, les ailes, les pattes, le thorax… L’unité structurale fonctionnelle est est la scolopidie .
Chaque scolopidie comporte trois cellules comparables à celles qui participent à la constitution de la soie innervée :
- Une cellule sensorielle fusiforme contient à son extrémité une vacuole réfringente surmontée par un tube rigide : le clou scolopal se terminant par un bouton ( la tête du clou ou bouton terminal ). La tête du clou, le clou scolopal et la vacuole réfringente sont traversé dans le sens de la longueur par un filament tendu : la corde axiale. Par sa base, la cellule sensorielle entre en rapport avec une cellule nerveuse de type ganglionnaire. Une cellule enveloppe entoure la partie terminale de la cellule sensorielle et le corps scolopal ; une cellule coiffe surmonte l’ensemble.

complexes fonctionnent comme des récepteurs de tension . Ils forment alors des organes chordotonaux distribués sur les antennes, les palpes, les ailes, les pattes, le thorax… L’unité structurale fonctionnelle est est la scolopidie .
Chaque scolopidie comporte trois cellules comparables à celles qui participent à la constitution de la soie innervée :
- Une cellule sensorielle fusiforme contient à son extrémité une vacuole réfringente surmontée par un tube rigide : le clou scolopal se terminant par un bouton ( la tête du clou ou bouton terminal ). La tête du clou, le clou scolopal et la vacuole réfringente sont traversé dans le sens de la longueur par un filament tendu : la corde axiale. Par sa base, la cellule sensorielle entre en rapport avec une cellule nerveuse de type ganglionnaire. Une cellule enveloppe entoure la partie terminale de la cellule sensorielle et le corps scolopal ; une cellule coiffe surmonte l’ensemble.
Physiologie animale- Ulg : http://www2.ulg.ac.be/physioan/chapitre/
Plusieurs scolopidies associées forment un organe chordotonal.
- Le clou scolopal transmet au système nerveux toutes sortes de vibrations , chocs ou sons, l’on peut parler dans ce cas « d’ouïe » de l’Insecte
Les organes chordotonaux peuvent être associés à des organes tympaniques ( tympan de la base du tibias de la patte antérieure du Grillon) Ceux-ci peuvent être ouverts sur l'extérieur mais peuvent aussi être fermés par une membrane tégumentaire ; dans ce cas, les cellules-coiffe s'insèrent dans la membrane qui sert alors de système récepteur, sélecteur et amplificateur de vibrations.On retrouve par exemple cette organisation dans les organes tympaniques (D) qui représentent chez les insectes le système auditif le plus évolué, capable selon la disposition et la structuration des scolopidies constituantes, de percevoir des sons allant de 5 kHz à 800 kHz, soit largement plus que l'oreille humaine (voir figure 8-15, ci-après). Typiquement, ces organes, généralement situés sur l’abdomen ou les pattes, comportent une fine membrane, le tympan, tendue sur un anneau sclérotinisé. Le tympan coiffe un sac trachéen faisant office de caisse de résonance. Les scolopidies forment un organe acoustique complexe soit directement fixé à la membrane tympanique soit, lorsqu’il est plus interne, en communication avec celle-ci et la cavité tympanique par un canal sanguin. On retrouve également des organes chordonotaux internes servant de propriocepteurs. Dans ceux-ci, des ligaments prolongeant les cellules-coiffe solidarisent les scolopidies à une paroi tégumentaire opposée. Ils peuvent alors faire office de tensorécepteurs, capables d'enregistrer les variations de pression interne du corps (D). Ils peuvent également intervenir dans l'équilibration et l'orientation spatiale de différents appendices renseignant alors sur le positionnement de pièces anatomiques par rapport à d'autres.
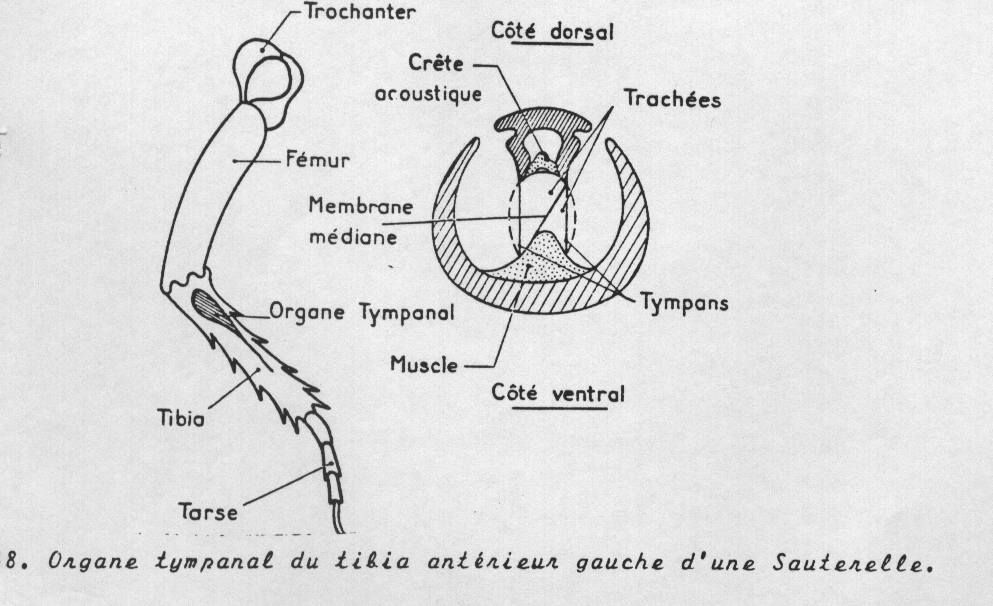
Fonctionnement de l’organe tympanal de la Sauterelle
Les ondes sonores font vibrer les membranes tympaniques qui sont fortement tendue et derrière lesquelles se trouve un espace rempli d’air . Les cellules sensorielles ( organes scolopaux) qui se trouvent dans so voisinage immédiat ( dans les crètes acoustiques sont alors excitées par les vibrations sonores.
1.4.5.2 Le sens chimique
a) les récepteurs du goût sont situés soit :
- Sur les antennes ( Hyménoptères )
- Sur les palpes ( Coléoptères)
- Sur les tarses ( Diptéres , Lépidoptères)
- Sur la trompe ( idem)
Ces récepteurs permettent de déceler les quatres saveur de base : salée, sucrée acide, amère.
b) Les récepteurs de l’odorat :
- Sont situés principalement près des antennes , très peu sur les palpes. Bien souvent, un Insecte ayant perdu ses antennes, mais gardé ses yeux intacts, sera perdu et ne trouvera plus son chemin.
La structure de ces deux groupes de récepteurs est cependant moins bien connue , de plus les substances chimiques entrant en jeux sont infiniment variées : cas des phéromones émises dans un but sexuel.
1.4.4 Le système glandulaire
Les insectes possèdent de nombreuses glandes, l’énumération qui suit ne saurait être exhaustive ; nous en reprendrons quelques types importants.
1.4.6.1 Les glandes exocrines
a) Les glandes tégumentaires
- Glandes cirières chez les abeilles, certain Homoptères.
Les ouvrières de l’Abeille domestique possèdent huit glandes cirières situées sur la partie interne des sternites des 4, 5, 6 et 7ème segments de l'abdomen. L'importance des glandes cirières varie suivant l'âge de l'ouvrière.
Elles atteignent leur maximum de développement vers le 12ème jour après la naissance et commencent à décroître à partir du 18 ou 19ème jour jusqu'à la mort de l'abeille.
Pendant cette période, les écailles de cire produites par ces glandes sont malaxées par les mandibules. L'abeille y incorpore, à ce moment, un solvant d'origine salivaire qui rend son malaxage plus aisé.
Ainsi trituré, l'écaille de cire entre dans la construction du rayon ou bien sert à l'operculation des alvéoles remplies de miel.
- Glandes laquifères : chez certains Coccides
- Glandes exuviales : Elles deviennent actives au moment de la mue. Elles sécrètent un liquide qui dissous la cuticule interne et externe , et de cette façon les métamorphoses larvaires sont possibles. Sans cette action, il serait impossible aux insectes d’augmenter de taille d’un stade larvaire à l’autre.
Les glandes exocrines sont constituées d'un ensemble de cellules que l'on nomme cellules glandulaires. Chez les insectes, il en existe trois types distincts :
- les cellules de type I
Elles ont une activité sécrétrice importante et n'ont pas de débouché direct sur l'extérieur de l'organisme. Le corps de l'insecte étant recouvert de cuticule imperméable, la sécrétion doit traverser cette barrière. Il existe donc de très fins canaux au contenu lipidique la traversant. Par exocytose, la sécrétion passe de la cellule au canal cuticulaire puis se mêle aux cires recouvrant la cuticule.
- les cellules de type II
Ce sont de grosse cellules situées à la base de l'épithélium et intercalées entre des cellules de type I dans lesquelles elles déversent leur sécrétion.
- les cellules de type III
Ces cellules peuvent elles rejeter leurs sécrétions à l'extérieur du corps grâce à un canal rejoignant cette fois directement les cellules aux canaux poraires de la cuticule.
Ci-dessous, un dessin de A. Quennedey illustrant les différents types de cellules (1, 2 et 3)
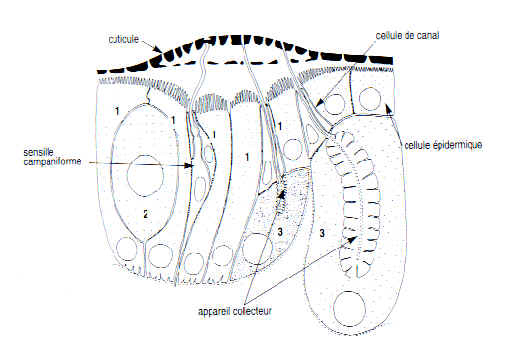
NB : la sensille campaniforme mesure la pression exercée sur la cuticule lorsque l'insecte frotte son abdomen contre le sol.
l'appareil collecteur collecte les sécrétions des cellules de type III. Il est connecté avec un canalicule qui traverse la cuticule.
Dans les glandes exocrines ont peut donc trouver des cellules de type I seules ou couplées à des cellules de type II, ou des cellules de type III, ainsi que l'association des trois types (cas général). Chez la blatte Nauphoeta cinerea mâle, par exemple, la phéromone est produite par des cellules de type III uniquement. Pour les femelles de Lepidoptères, les glandes ne sont constituées que de cellules de type I. Enfin chez les termites c'est l'association des trois types qui est rencontré.
Ces cellules glandulaires permettent la production de mélanges chimiques très complexes.
b) Les glandes associées à des appendices
Sur les ailes de beaucoup de papillons mâles, on trouve des écailles de forme spéciale, disséminées dans les autres ou réunies en bandes . On les appelle androconies. A la base de ces écailles, se trouvent des cellules glandulaires secrétant une substance excitant la femelle et favorisant l’accouplement. Ces produits portent le nom de phéromones et sont très volatiles.
Il existe également des cellules glandulaires sur les tibias postérieurs de certains Lépidoptères ( Hépialus).
Les phéromones
La notion de phéromone représente donc un aspect de la transmission de l’information entre individus. Certaines phéromones agissent par voie orale, mais la majorité d’entre elles agissent par voie olfactive.
Les cellules sécrétrices de phéromones peuvent :
- Etre isolées, dispersées sur tout le corps ( Criquets )
- Former des glandes à structure plus ou moins complexe et être localisées dans une région précise du corps ( glandes de Nassanof chez les ouvrières d’abeilles appartenant au septième tergite abdominal).
La glande de Nasanoff est située sur la face dorsale abdominale des abeilles, elle produit une phéromone aux fonctions multiples. Elle sert à marquer l'entrée de la ruche, un lieu intéressant comme une source de nectar, une source d'eau ou un lieu d'arrêt provisoire lors de l'essaimage. Pour diffuser la phéromone, les abeilles exposent leur abdomen et ventilent en battant des ailes. L'odeur de la phéromone guide les autres ouvrières.
Selon leur mode d’action, on distingue parmi les phéromones, les deux catégories suivantes :
a) Les phéromones modificatrices
Entraînent des modifications physiologiques des Insectes récepteurs, par exemple chez les Insectes sociaux, elles interviennent dans la détermination des castes, le bloquage ovarien…
Elles agissent sur la physiologie de l’animal, et ont donc un différé. La fonction principale de ces phéromones est de coordonner le développement physiologique et comportemental d’un ensemble d’individus. Du fait de leur action à plus long terme, elles sont difficiles à identifier.
b) Les phéromones incitatrices ou « de déclenchement ».
Produisent une réaction instantanée et réversible, suivant la nature du comportement qu’elles induisent , on les classe en :
- Phéromones de piste : Chez les Insectes sociaux, le chemin entre la source de nourriture et le nid , par exemple, est marqué par le dépôt de gouttelettes de substance
Il a été montré que les termites les plus archaïques et les termites dont les populations restent confinées dans leur site de nidification sécrètent une phéromone de piste. Ce terme recouvre toutefois une certaine hétérogénéité fonctionnelle. Chez Mastotermes darwiniensis, la phéromone permet simplement semble-t-il de marquer l’environnement. Chez les Termopsidae et les Kalotermitidae, elle oriente et chez la plupart des Rhinotermitidae et des Termitidae, non seulement elle oriente, mais elle recrute aussi les termites sur les pistes. Dans certains cas, une seule molécule suffit à rendre compte des deux effets, mais dans d’autres cas, l’intervention de composés annexes n’est pas exclue.
- Phéromones d’agrégation : Les agrégations peuvent être temporaires : essaims, rassemblement de Scolytes lors d’attaques massives d’arbres.
- Phéromones d’alarme : déclenchent un comportement de défense ou d’attaque.
Les espèce du genre Formica, par exemple, lorsqu'un ennemi les menace, recourbent leur abdomen entre leurs pattes postérieures et arrosent l'ennemi d'un jet d'acide formique qui agit comme un produit d'alarme pour les autres fourmis et comme un produit de défense contre l'intrus.
Ce double rôle alarme et défense peut aussi être dévolu aux glandes mandibulaires comme chez la fourmi australienne Calomyrmex. Les ouvrières de cette espèce, lorsqu'elles sont en danger, sécrètent des gouttelettes originaires des glandes mandibulaires, ce qui provoque une accélération de la démarche des ouvrières du voisinage et des vibration de leurs antennes. Elles sont ainsi en état d'alerte. De plus, la sécrétion des glandes mandibulaires est très gênante pour l'ennemi sur qui elle est projetée. En effet, elle devient visqueuse à l'air libre, l'ennemi est alors obligé de se nettoyer et ne peut pas attaquer. On a aussi pu constater qu'elle avait un rôle répulsif. En effet, sa présence dans une source de nourriture dissuade les fourmis à viande de s'en approcher.
- Phéromones sexuelles : produites généralement par l’un des sexes seulement et agissent sur le sexe opposé. Les attractifs sexuels sont le plus souvent produits par les femelles , ils permettent la rencontre des sexes ; chez de nombreux insectes, des aphrodisiaques souvent produits par le mâle ( par les androconies) agissent ensuite à courte distance , interviennent à un moment précis de la séquence comportementale et favorisent l’accouplement.
a) Les glandes associées à des segments
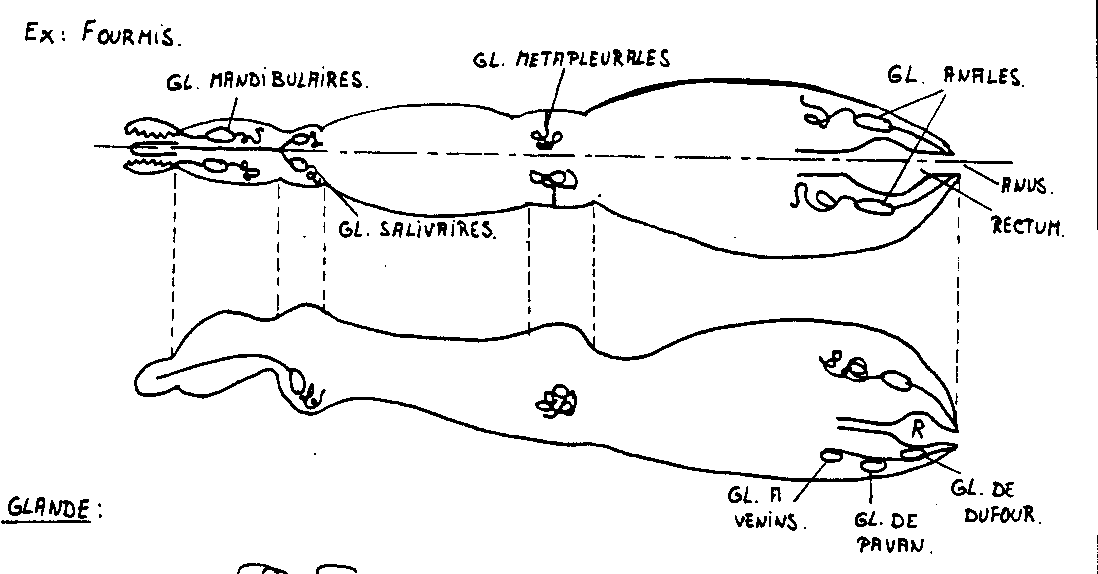
- Les glandes mandibulaires
Elles émettent des phéromones ( géraniol, etc…) qui , selon la concentration, auront un effet attractif ou répulsif. Une substance pour deux effets : économie de moyens.
Ce sont les glandes responsables, par excellence, de l'alarme, chez beaucoup de fourmis.
La Formicinae Lasius umbratus, par exemple, en présence de têtes écrasées, présente un comportement typique d'alarme : Sa vitesse de déplacement augmente, ses mandibules sont écartées, parfois, elle sautille sur place puis ferme ses mandibules sur un congénère trop proche. Les sécrétions mandibulaires contiennent surtout du citronellol qui a un rôle défensif et un peu de citronellal qui est un déclencheur d'alarme.
Schéma : glandes exocrines d’une fourmi
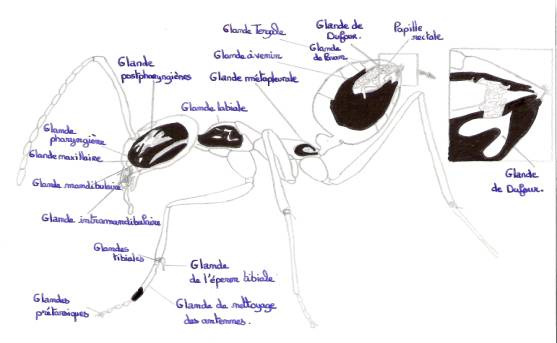
- Les glandes labiales ou salivaires
Glandes très volumineuses, s'étendant souvent dans le thorax.
La salive peut remplir plusieurs rôles :
- Humidification des aliments et lubrification des pièces buccales.
- Transformer, grâce à un enzyme, l’amidon en glucose.
- Posséder des propriétés irritantes et être anticoagulantes
- Posséder des propriétés toxiques vis à vis des tissus végétaux, par exemple.
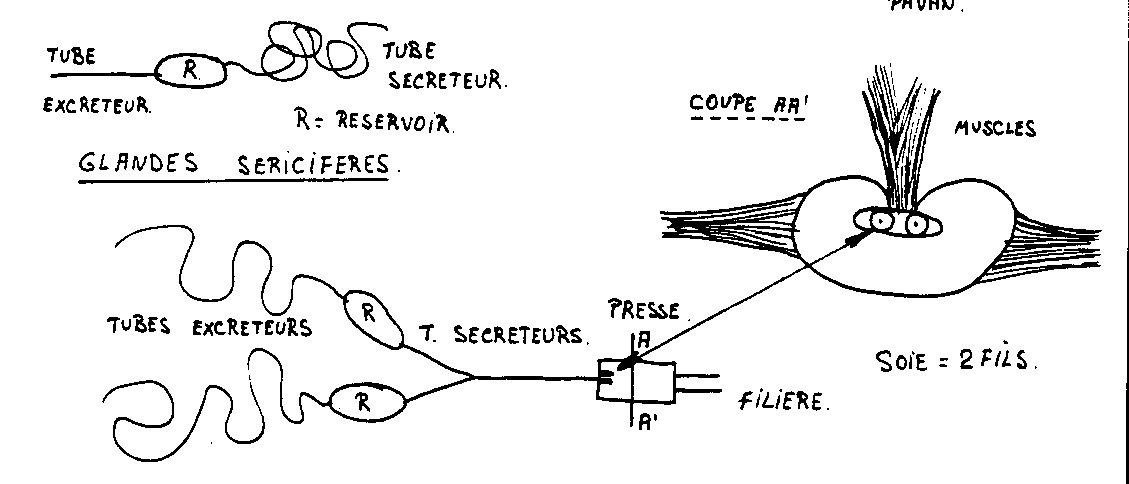
- Les glandes séricigènes
Ce sont des glandes salivaires modifiées qui produisent de la soie. Voir schéma. Elles peuvent être céphaliques, rectales ou tarsales . Sauf quelques exceptions chez les insectes, la soie est toujours sécrétée par les larves ( Bombyx mori). Le fil de soie se compose de deux brins et chaque brin est formé de trois substances : Au centre, la fibrine entourée par le grès ou séricine et la mucoïdine.
- Les glandes métapleurales
Elles émettent des phéromones : donnant par exemple leur odeur à la fourmilière son odeur spécifique, perceptible par les antennes, propriétés antiseptiques.
- Glandes anales et glandes répugnatoires
Elles émettent des liquides malodorants ou nocif à fonction défensive . Glandes abdominales des Hétéroptères, des Blattes. Glandes pygidiales des Coléoptères. Osmétérium des Lépidoptères
- Glandes de Dufour
Aussi appelées glandes alcalines, elles produisent des alcalis qui réagissent sur les sécrétions acides des glandes venimeuses inoculées par aiguillon chez les Hyménoptères. Ce liquide sert aussi, chez les fourmis, à marquer les pistes.
Elles ont un rôle actif chez les Formicinae esclavagistes qui ont leurs glandes mandibulaires réduites. Elle ont un double rôle dans les raids de pillage : Elles excitent et recrutent les congénères esclavagistes mais répulsent et dispersent, dans un comportement de panique les ouvrières de l'espèce pillée.

- Les glandes à venin
Le venin des abeilles et des guêpes contient de l’acide formique et un hydrocarbure.

Le système endocrinien assure avec le système nerveux la coordination entre les activités des différents organes ou tissus, il permet à l’organisme d’adapter se physiologie aux modifications du milieu. Le système endocrinien utilise les liquides qui circulent dans le corps, porte ses messages chimiques ou hormones aux organes cibles. Cette coordination plus lente que celle exercée par le système nerveux , porte ses effets sur des processus généralement de plus longue durée : la croissance, le maintien de concentrations appropriées en métabolites dans l’hémolymphe…
1.4..6.2.1 Corpora cardiaca, corpora allata, ganglions prothoraciques.
Croissance, mues et métamorphoses
Ces processus ont été le mieux étudiés chez les insectes et les crustacés, essentiellement pour des raisons économiques.
Le développement postembryonnaire des insectes peut s'effectuer selon quatre modalités ( voir plus loin) différentes suivant l'importance plus ou moins grande des changements métamorphiques qui les font passer d'une larve sans organes sexuels fonctionnels à un adulte capable de se reproduire. On distinguera ainsi des insectes amétaboles, paurométaboles, hémimétaboles et holométaboles.
Chez les amétaboles, insectes primitifs aptérygotes (collemboles, protoures, thysanoures), les jeunes sont semblables aux adultes mis à part la taille et l'absence d'organes sexuels fonctionnels. Ils ont également le même mode de vie que les adultes. La croissance résulte d'une suite de mues. Au cours de l'une d'entre elles, que l'on qualifie de mue imaginale, l'insecte acquiert des gonades fonctionnelles le transformant en adulte capable de se reproduire (imago). L'insecte adulte continuera de grandir au cours de mues postimaginales.
Certains insectes ptérygotes dits paurométaboles (dictyoptères, hémiptères, orthoptères, phasmoptères) ont un mode de croissance assez semblable. La différence essentielle est l'apparition des ailes au cours de l'avant dernière mue, qualifiée de nymphale. La mue suivante, la dernière, est la mue imaginale au cours de laquelle les organes sexuels deviennent fonctionnels.
Chez les hémimétaboles (éphéméroptères, odonatoptères), la larve a un habitat et un mode de vie différent de l'adulte (aquatique contre terrestre). La mue imaginale entraîne donc des transformations plus importantes. Comme chez les paurométaboles, les ébauches des ailes apparaissent à la mue nymphale. La nymphe reste libre, mobile et capable de se nourrir.
Chez les holométaboles (coléoptères, diptères, lépidoptères, névroptères), la mue nymphale donne naissance à une pupe caractérisée par son immobilité extérieure quasiment totale. C'est au cours de ce stade pupal que la nymphe va se métamorphoser en adulte.
Bien que la métamorphose soit pour les morphologistes caractéristique des holométaboles, des changements importants de type métamorphique s'effectuent également dans les autres groupes lors des mues nymphales et imaginales. La croissance présente par ailleurs dans tous les groupes un même caractère discontinu, étant réglée par des "sauts" (mues) successifs. Le contrôle endocrinien du développement, tel qu'il a été étudié jusqu'à présent, présente d'ailleurs des caractéristiques communes dans les différents groupes. Il ne semble pas y avoir de différences essentielles en fonction de l'importance plus ou moins grande des changements s'opérant lors des mues nymphales ou imaginales ou de la pupaison.
Il paraît clair à l'heure actuelle que l'entrée en mue, quel que soit le type de celle-ci, dépende essentiellement de la production, au niveau de cellules neurosecrétrices cérébrales, d'une hormone prothoracotrope (PTTH, ecdysiotropine, prothoracotropine). Cette hormone passe par transport axonal dans les corpora cardiaca, organes neurohumoraux associés au cerveau, où elle est stockée. Sa sécrétion, suite à une série de stimuli externes et internes appropriés, va induire la production d'a -ecdysone par la glande prothoracique. L'a -ecdysone est rapidement hydroxylée en C20 dans les tissus adipeux pour donner la b -ecdysone (HM, hormone de mue), beaucoup plus active que la forme a. Cette dernière doit donc être considérée plus comme un précurseur que comme l'hormone de mue elle-même. La b -ecdysone paraît à la base de l'ensemble des modifications préparant et accompagnant les différents types de mue.
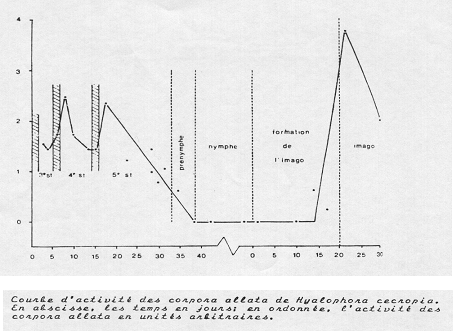
Le caractère larvaire ou non d'une mue paraît essentiellement déterminé par la sécrétion plus ou moins abondante d'une hormone juvénile (HJ )issue des corpora allata. Ces structures neurohumorales sont voisines des corpora cardiaca et sont sous contrôle direct d'allatostatines et d'allatolibérines produites par le cerveau. C’est en fait la présence de HJ en concentration plus ou moins importante par rapport à l'hormone de mue qui paraît déterminer le caractère larvaire, nymphal ou imaginal de la mue. Au cours de la mue imaginale donc, l'activité des corpora allata et la production de HJ sont pratiquement nulles. Elles reprennent après cette mue alors que l'activité des corpora cardiaca et de la glande prothoracique régresse. L'hormone juvénile produite par l'imago va intervenir dans le contrôle du comportement sexuel, l'activité des glandes génitales et la gamétogenèse
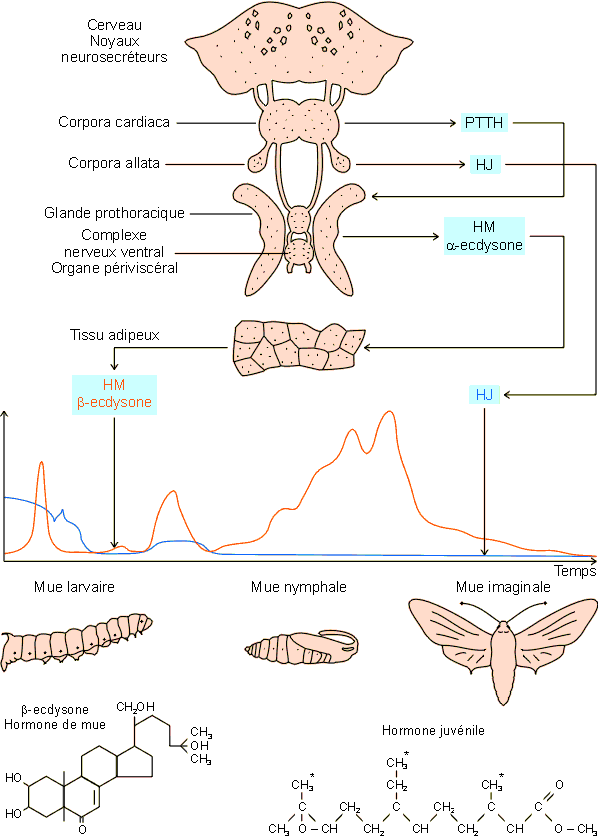
Physiologie animale : Ulg :http://www2.ulg.ac.be/physioan/chapitre/
Contrôle hormonal des différents types de mue chez les insectes. Détails dans le texte. L'hormone juvénile existe sous différentes formes, variables suivant le type de radicaux, méthyle ou diméthyle, se trouvant aux positions *.
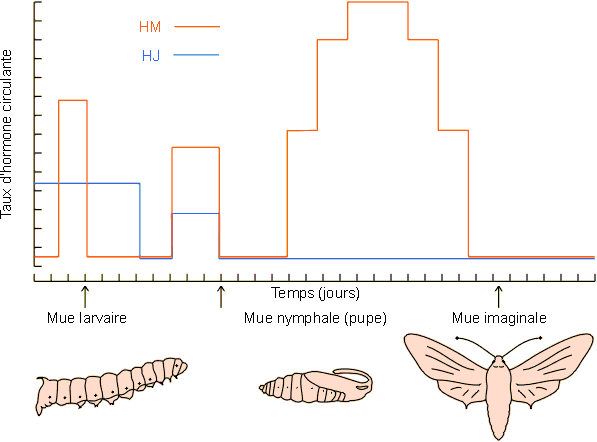
Evolution schématique des taux d'hormone de mue (HM) et juvénile (HJ) aux cours de différentes mues chez Manduca sexta. D'après des données reprises de Landau, Biggers et Laufer, dans Danzler 1997, modifié.
Les mues s'accompagnent d'une exuviation suivie d'un tannage et d'un durcissement de la nouvelle exocuticule. Ces processus sont sous contrôle hormonal. Un facteur neurohumoral, l'hormone d'exuviation (EH : éclosion hormone), ayant des effets directs sur différents phénomènes impliqués dans ce processus (assouplissement des structures alaires, contractions musculaires, etc.…) est en effet produit par des cellules spécialisées du complexe corpora cardiaca - corpora allata. L'hormone d'exuviation induit par ailleurs la production de bursicon, par le complexe nerveux périviscéral. Cette hormone est directement impliquée dans l'expansion des ailes, la sclérotinisation de l'exocuticule de même que dans sa mélanisation éventuelle.
Bon nombre d'insectes peuvent suspendre l'essentiel de leurs activités biologiques pendant des temps plus ou moins longs à différents moments de leur développement ou de leur vie adulte. Ces périodes, dites de diapause, coïncident généralement avec des moments où les conditions de vie deviennent défavorables (hibernation, estivation, déshydratation, etc.…). Les diapauses larvaires, nymphales et imaginales semblent essentiellement en rapport avec la mise en repos du système endocrine. On observe ainsi une diminution importante de la production de PTTH et du taux d'ecdysone circulante. Il en va de même pour la HJ dans différents cas. Chez certaines espèces cependant, le taux de HJ est élevé au cours des diapauses larvaires et pupales. L'hormone paraît ici exercer un contrôle inhibiteur sur la production de PTTH.
Les facteurs déclenchant les différents types de mues et de diapauses restent en général très mal connus. Dans la plupart des cas, il semble qu'il faille faire intervenir le jeu de facteurs internes et externes. L'ingestion de nourriture est chez beaucoup d'espèces un facteur externe important. Ainsi, chez la tique Rhodnus prolixus, la mue ne se produit qu'après la prise d'un important repas de sang qui provoque une distension abdominale servant de facteur déclenchant. Une larve de 50 mg peut ainsi absorber jusqu'à 300 mg de sang. Chez le criquet Locusta migratoria, la propagation de messages sensoriels provoqués par la déglutition au niveau pharyngien paraît indispensable à la sécrétion des hormones de mue. Chez le papillon Bombyx mori, la diapause embryonnaire paraît déterminée par les conditions photopériodiques subies par la mère. Ainsi les reproductrices élevées en jours longs produisent des œufs à diapause tout à fait caractéristiques, riches en hydroxykynurénine et de teinte violette. En jours courts, la diapause embryonnaire n’apparaît pas.
L’Hormone de mue ou ecdysone
Le cholestérol est le précurseur de cette hormone . Alors que les Vertébrés sont capables de réaliser la synthèse du cholestérol, les arthropodes ne le peuvent pas, donc ils doivent trouver dans leur alimentation les stérols nécessaires.
En dehors de la synthèse de cette hormone, les stérols remplissent encore d’autres rôles importants dans la physiologie de l’insecte :
- Ils permettent le tannage et le durcissement des protéines ( le tégument externe rigide est constitué de protéines tannées).
- Ils jouent un rôle phagostimulant, c’est à dire que leur présence incite les insectes à mordre les feuilles. Explication de l’inféodation d’un insecte à un type végétal.
Cette hormone agissant seule provoque , comme nous l’avons vu plus haut, la mue imaginale c’est à dire l’apparition d’un insecte adulte.
L’hormone juvénile
Les terpènes en sont les précurseurs, produits largement représentés dans le monde végétal. Si on examine chez les Mammifères la synthèse du cholestérol à partir des groupements acétyl , on constate que les produits intermédiaires formés sont justement les terpènes. L’accumulation dans le foie d’acides terpéniques bloque l’enzyme responsable de la synthèse du cholestérol.
Il en est de même chez l’insecte, la présence de terpènes bloque cette synthèse cholestérique .
Conclusion : la consommation de terpènes lors de la synthèse de l’hormone juvénile va permettre , à terme , le redémarrage de la production de cholestérol et par là permettre la synthèse de l’hormone de mue.
Contrairement à l’hormone de mue qui mettait seule un point final au développement de l’insecte par l’occurrence de la mue imaginale, l’hormone juvénile sera responsable des mues larvaires, l’insecte reste sous forme de larve et continue à se développer. Deux hormones de juvéniles très connues sont la néotinine et la juvabione . La néotinine est très peu spécifique, la juvabione beaucoup plus.
Ces hormones peuvent avoir une portée importante dans la lutte insecticide ( utilisation de mimétiques de l’hormone juvénile).

L’hormone sexuelle
Responsable des caractères sexuels secondaires
L’hormone gonadotrope
Responsable de la croissance des ovocytes.
L’hormone des changements de coloration ou contractine
Cette hormone déversée dans l’hémolymphe a pour effet de concentrer les granules pigmentaires dans les chromatophores.
Les neuro-hormones
- Les hormones antidiurétiques et diurétiques agissent sur les cellules des tubes de Malpighi, du rectum et des glandes rectales responsables du niveau hydrique des tissus.
- Hormone plastifiante qui changent les propriétés de la cuticule abdominale , lui permettant de se distendre pour s’accommoder au volume des repas de sang pris par certains insectes.
- Hormone hyperglycémiante d’action analogue à celle du glucagon chez les Vertébrés.
Hormone de diapause
Présente dans le ganglion sous oesophagien, elle stimule la diapause embryonnaire.
Le Bursicon ou facteur de tannage
Contrôle le tannage du puparium
L'hormone d'exuviation induit la production de bursicon, par le complexe nerveux périviscéral. Cette hormone est directement impliquée dans l'expansion des ailes, la sclérotinisation de l'exocuticule de même que dans sa mélanisation éventuelle.
1.4.4 Les organes génitaux
Chez les insectes, les sexes sont séparés, les cas d’hermaphrodisme sont rares. Ce gonochorisme strict a en général pour conséquence un dimorphisme sexuel accentué.
1.4.4.1 L’appareil reproducteur femelle
Deux ovaires ,deux oviductes, vagin.
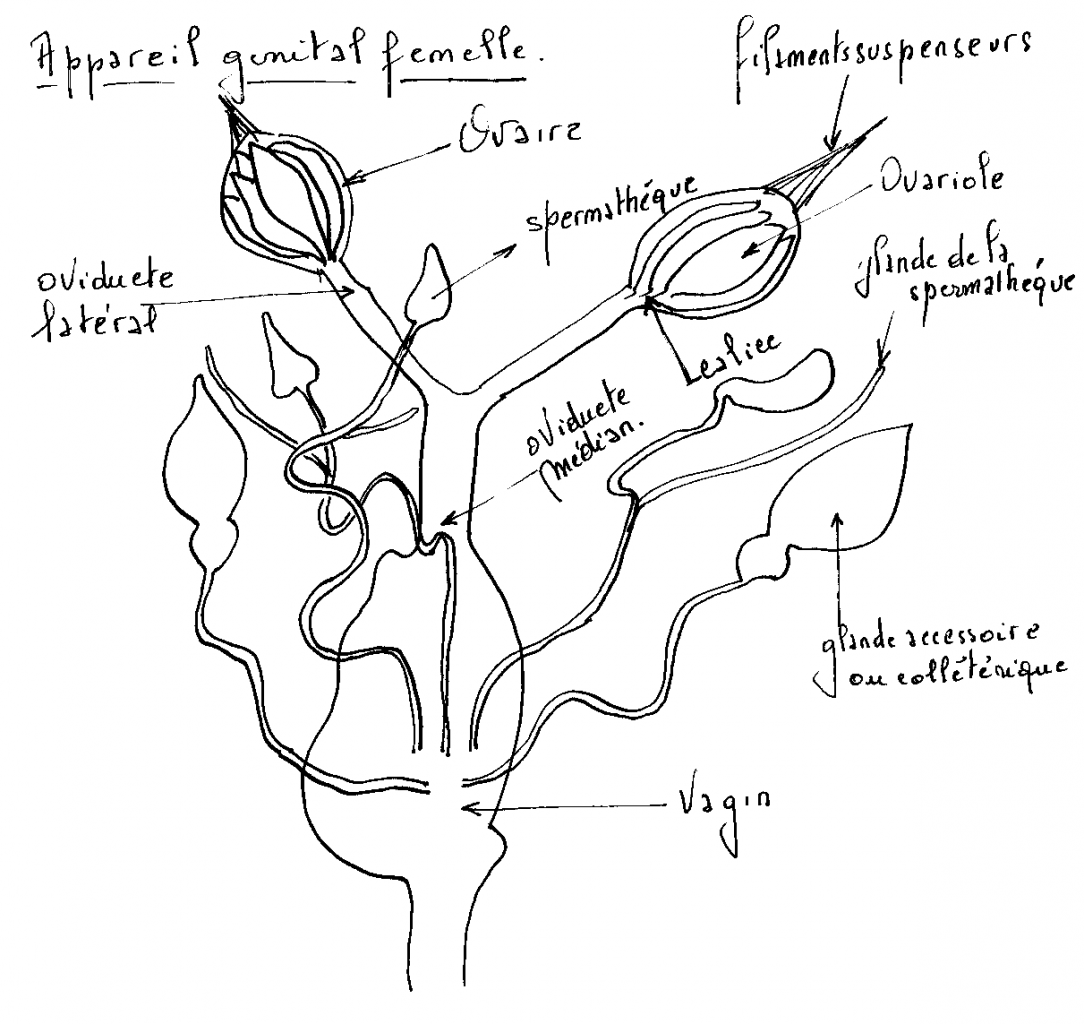
Réceptacle séminal ou spermathèque et glandes annexes.
L’orifice copulateur peut être distinct de l’orifice de ponte. Les glandes annexes aussi appelée glandes collétériques sécrètent la matière visqueuse servant à cimenter les œufs entre eux ( formation oothèque) ou à les fixer sur un support. Chaque ovaire est constitué par un nombre variable d’ovarioles ( certains Aphidiens ne possède qu’une seule avariole par ovaire, certains termites supérieurs en comportent jusque 2400 ).
Chaque ovariole comprend une série d’ovocytes de taille régulièrement croissante, disposés en chapelet et sur lesquels se soudent les parois ovigères. Il aboutit par sa base élargie dans sa partie proximale ou calice de l’oviducte
a) Organisation des ovarioles
Chaque ovariole peut être divisé en quatre régions :
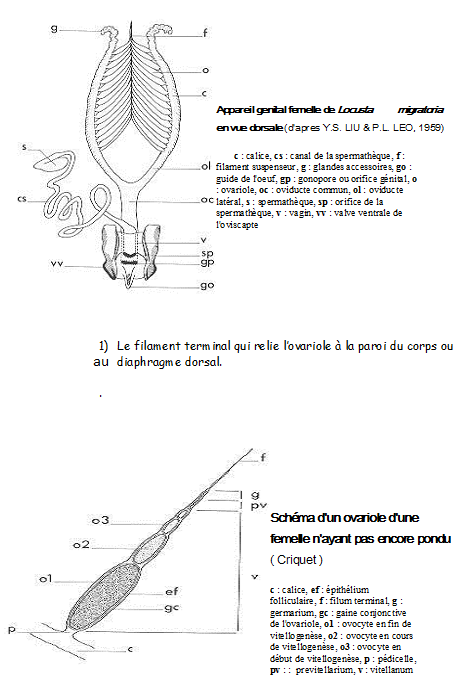
2) Le germanium qui contient les cellules germinales primordiales dont l’évolution aboutit exclusivement à la formation d’ovogonies puis d’ovocytes (ovaires panoïstiques) ou à la formation d’ovogonies et de cellules nourricières ( ovaires méroïstiques)
3) Le vitellarium ou zone de croissance qui représente la majeure partie de l’ovariole . Contient une série lminéaire d’ovocytes en voie de vitellogénèse . Chaque ovocyte y est entouré par un sac épithélial : le follicule.
4) Le pédicelle , tube à parois minces qui aboutit au calice. Obturé par un bouchon épithélial compact qui est détruit lors de chaque ovulation.
B. Les principaux types d’ovarioles
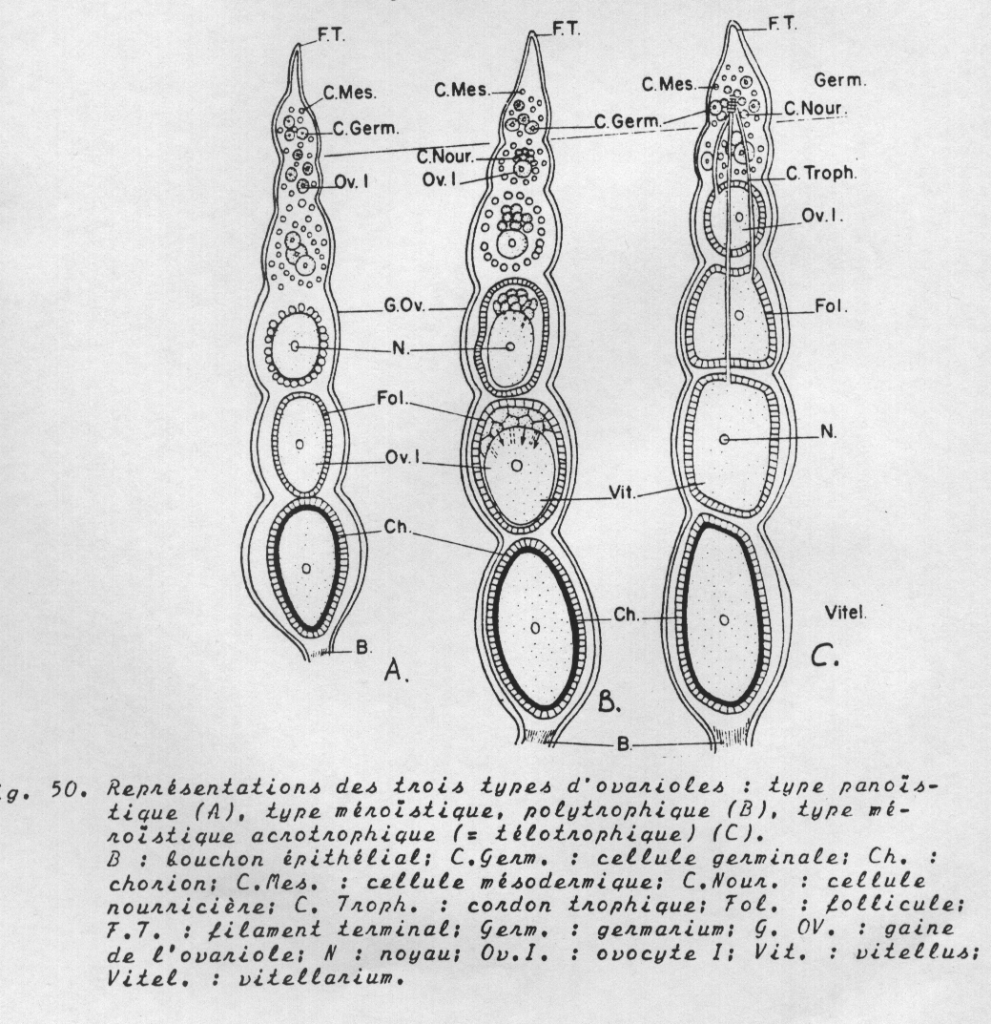
- Le type panoïstique : toutes les cellules germinales évoluent en ovocytes qui seront pondus. Il n’existe pas de cellules nourricières et les éléments nutritifs sont empruntés à l’hémolymphe directement par les ovocytes ou par les cellules folliculeuses qui en assurent le transfert dans les ovocytes.
- Le type méroïstique : dans ce type, les cellules germinales engendrent par division deux catégories de cellules sœurs, des ovocytes qui évolueront normalement et qui seront pondus et des cellules nourricières qui assurent l’approvisionnement des ovocytes au début de la vitellogénèse puis dégénèrent
Dans ce type, on distingue :
- Des ovaires polytrophiques : chaque ovocyte est accompagné par un nombre spécifique de cellules nourricières.
- Des ovaires acrotrophiques ( = télotrophiques) : Les cellules nourricières restent accrochées à la partie supérieure du germanium où elles ont pris naissance et assurent l’approvisionnement de l’ovocyte correspondant par l’intermédiaire de long cordons trophiques. Après la rupture des cordons et la dégénérescence des cellules nourricières, la nutrition des ovocytes est assurée par le follicule.
1.4.4.1 L’appareil reproducteur mâle
Deux gros testicules, canaux déférents et canal éjaculateur. Ce dernier traverse à son extrémité un organe servant à la copulation ou aedeagus.
Les testicules sont constitués par un faisceau de tubes séminifères ou follicules testiculaires dont le nombre varie selon les espèces ( Poux : 2, Orthoptères : très grand nombre. )
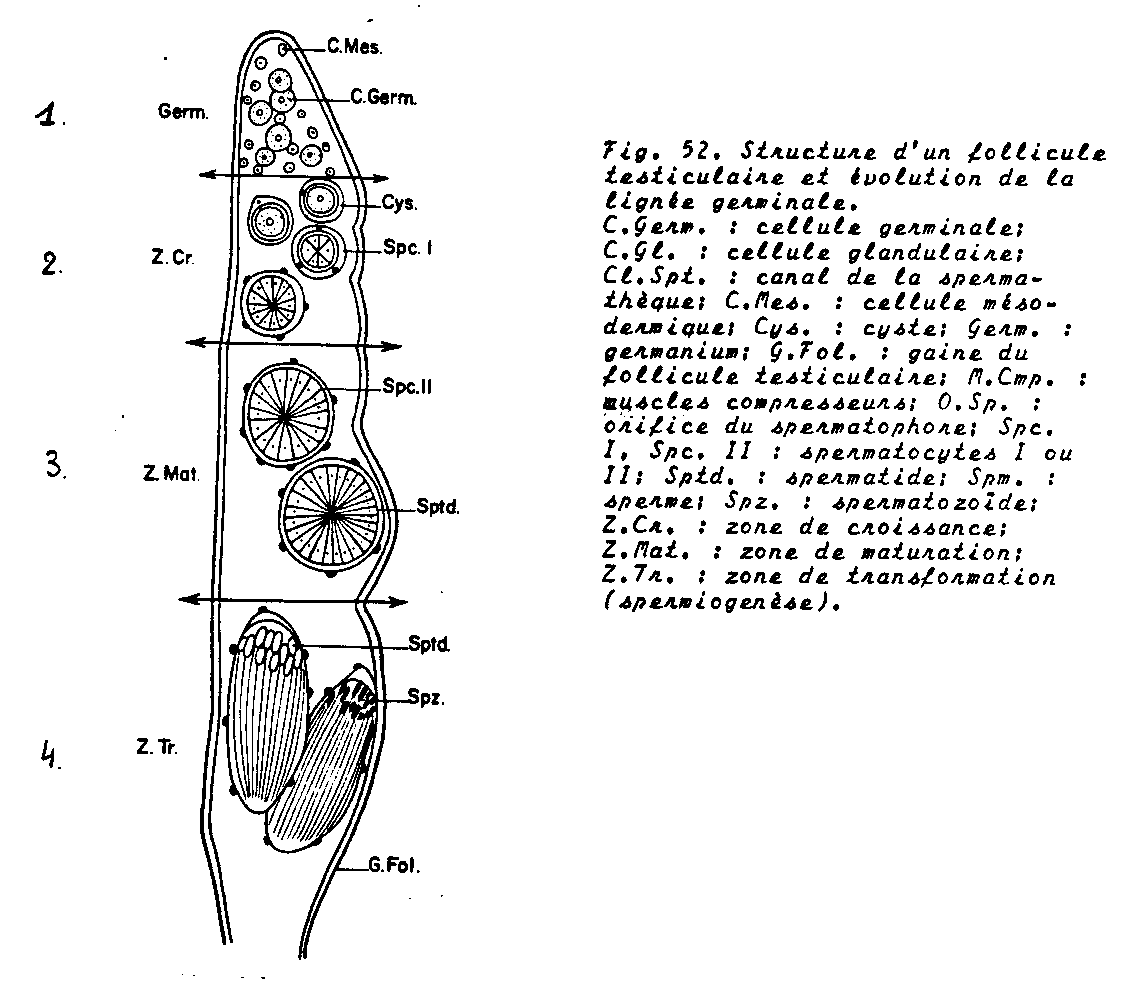
Tubes séminifères et spermatogénèse
- Le germanium ou zone des spermatogonies.
- La zone de croissance des spermatocytes I , chaque spermatogonie qui quitte le germanium est immédiatement entourée par une assise de cellules mésodermiques . A l’intérieur de ce cyste, elle se divise activement et engendre des spermatocytes de premier ordre . Chaque cyste contient selon les espèces , 64 à 256 spermatocytes I. Il n’y a pas de cellule nourricière.
- La zone de maturation =zone de la méiose. A l’intérieur de chaque cyste, la division hétérotypique et ensuite la division homéotypique , assurent respectivement la formation des spermatocytes II puis des spermatides. A l’issue de ces phénomènes, le nombre de cytes est multiplié par 4 ( voir méiose)
- La zone de transformation = zone de spermiogénèse.
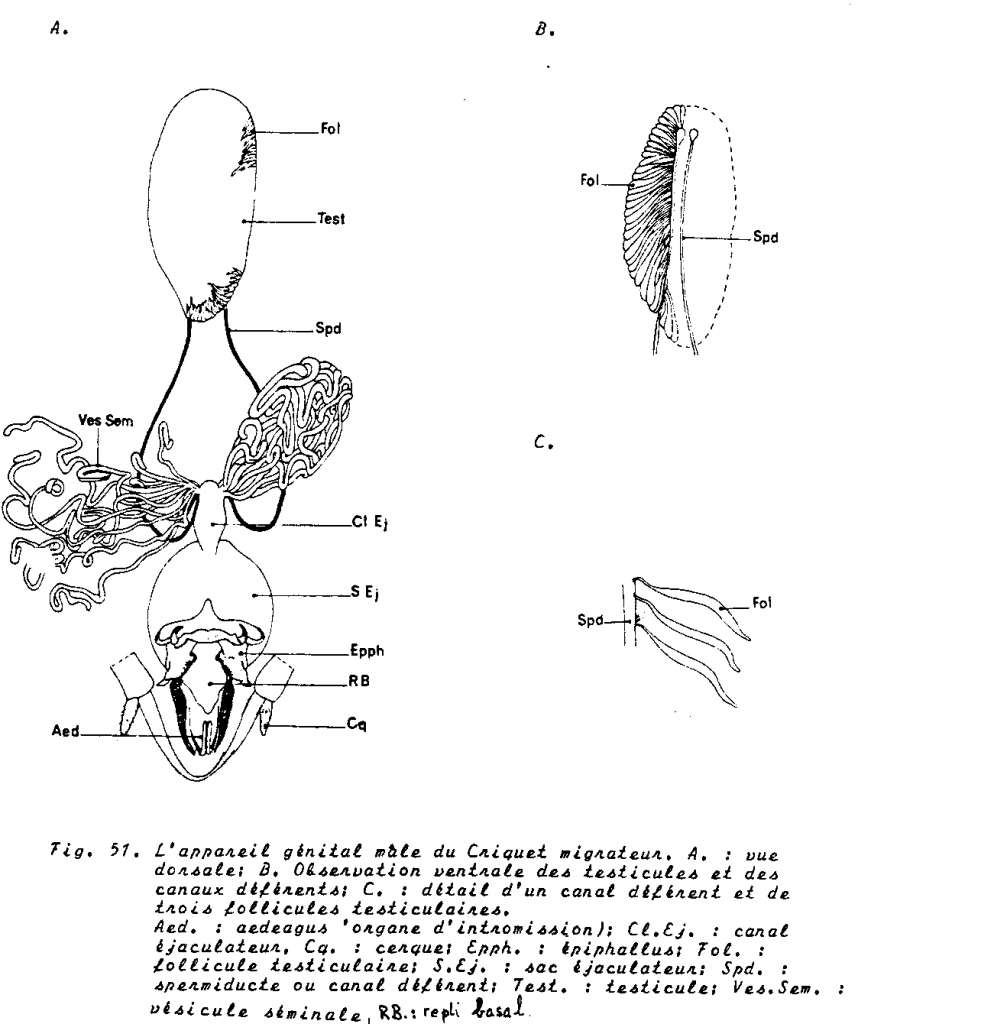
Les spermatozoïdes quittent finalement leur cyste d’origine et gagnent les spermiductes, ils se dissocient dans les vésicules séminales. Les spermatozoïdes-ci font un séjour dans les glandes annexes qui permettent de les diluer et de les activer.
1.4.8 La vision chez les insectes
Les insectes peuvent posséder deux types d’organes de la vision :
- Les yeux simples ( ocelles ), latéraux ( stemmates) ou dorsaux
- Les yeux composés.
1.4.8.1 Les yeux simples
a) Les ocelles dorsaux
Les ocelles dorsaux existent en même temps que les yeux composés chez les imagos et les larves d’hétérométaboles.
Ils sont situés sur le vertex et sont typiquement au nombre de trois , disposés en triangle sur la tête.
Chaque ocelle comprend :
- Une cornée unique au niveau de laquelle la cuticule apparente se renfle habituellement en une lentille biconvexe.
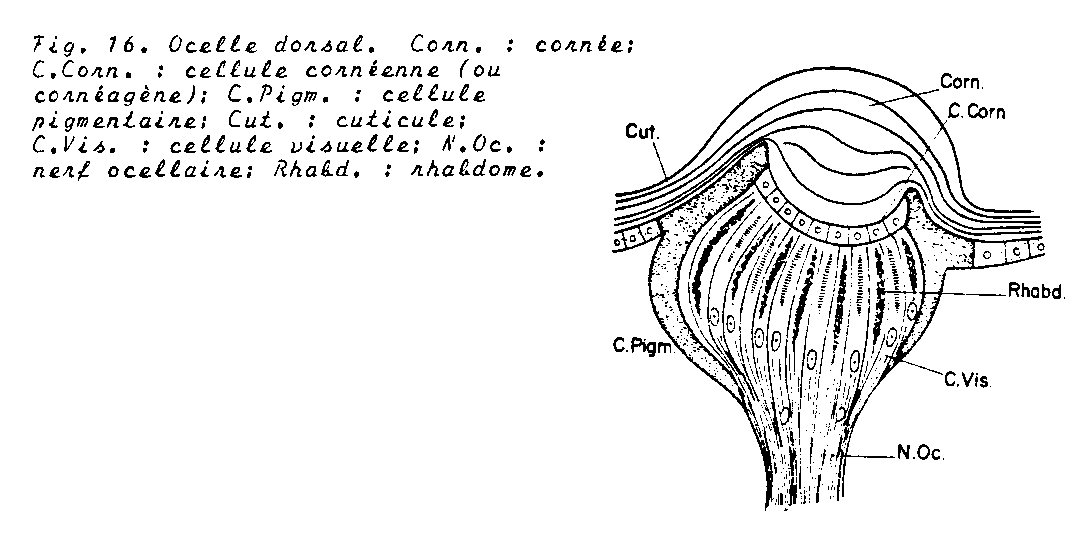
- Les cellules épidermique sous jacentes ou cellules cornéagènes peuvent s’allonger et se regrouper en un corps vitré qui s’adjoint au point de vue fonctionnel à la cornée.
- Les cellules réceptrice avec une partie finement striée, le rhabdomère, semblable à celui des yeux composés . Ces cellules peuvent associer leur rhabdomère pour former le rhabdome
Les neurones des cellules visuelles quittent l’ocelle et établissent dans le nerf ocellaire des contact synaptiques avec des neurones d’association.
Leur rôle : Ils agiraient comme des organes sensoriels stimulateurs en relation avec la fonction du vol et l’activité générale ( ex : si on masque les yeux composés d’une abeille, elle se comporte comme si elle était aveugle bien que les ocelles soient découverts : au contraire, avec les ocelles masqués l’Abeille voit, mais les changements qui interviennent dans les variations de l’intensité de la lumière ne lui parviennent que lentement ; les ocelles de ce types accroîtraientla sensibilité phototropique des centres nerveux)
Elles donnent des informations sur les variations de l’éclairement.
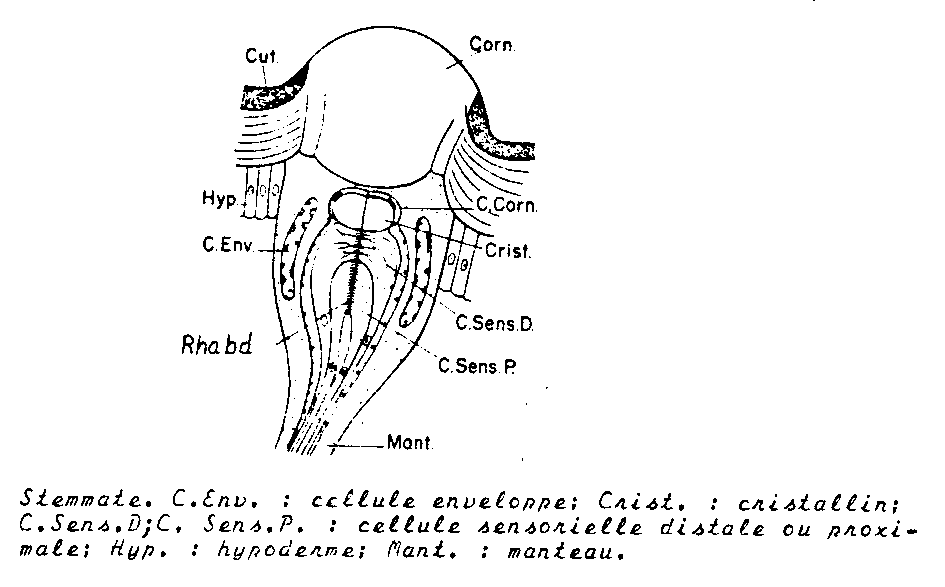
b) Les ocelles latéraux ou stemmates
Les ocelles latéraux sont les seuls organes visuels des larves d’holométaboles
Ils occupent une position voisine de celle qu’occupent les yeux composés imaginaux , ce ne sont pas les précurseurs de ces derniers, ils dégénèrent à la métamorphose.
De 1 à 7 de chaque côté de la tête, ils sont reliés aux lobes optiques du protocérébron, comme le seront les yeux composés.
Leur rôle : les stemmates permettent une vision des formes, bien qu’elle soit certainement imparfaite
1.4.8.2 Les yeux composés
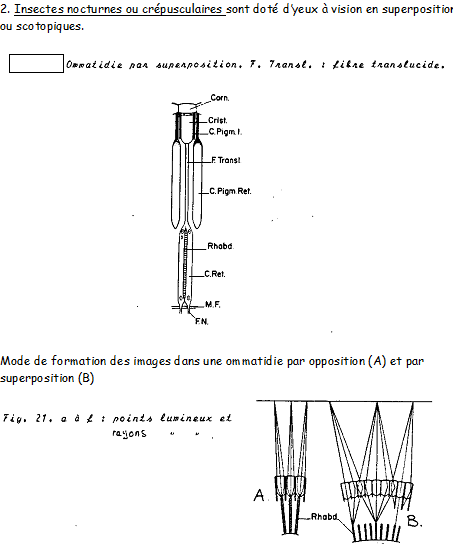
Présents chez l’adulte, ainsi que chez les larves d’hétérométaboles . Toujours au nombre de deux, caractéristique des Arthropodes.
Yeux composés = somme d’un nombre plus ou moins grand d’unités : les ommatidies.
Remarque : ce nombre est en rapport avec le mode de vie de l’Insecte. Elevé chez les espèces à vol rapide. Ex : les yeux des Libellules en contiennent plus de 10.000, nous n’en trouvons que 6 chez certaines fourmis souterraines.
Structure : Schéma semblable dans ses grandes lignes à celui d’une ocelle, à savoir :
- Une cornée, un cristallin variable, des cellules rétiniennes se prolongeant par des fibres nerveuses, des rhabdomères, des cellules pigmentaires.

La vue d’un insecte : ils ne voient bien qu’à un mètre de distance environ . L’abeille qui a pourtant une bonne vue parmi les insectes a une vision qui correspond à peine à un centième de celle de l’homme . Par contre, les yeux composés semblent particulièrement aptes à suivre des mouvements étant donnés qu’un objet mobile est enregistré successivement par les différentes facettes de l’œil. Donc d’une importance pour l’insecte puisque l’agresseur à éviter comme la proie à capturer sont des objets en mouvement. Les papillons perçoivent les couleurs comme les mammifères.
Généralement, les Insectes évoluent dans un univers riche en couleurs , mais dont la gamme perçue est différente de la nôtre. Exemple : les abeilles ne distinguent pas le rouge, mais bien l’ultra violet , le bleu, le jaune, le blanc, le noir . Il est à remarquer que les fleurs pollinisées par ces insectes ont ces couleurs. De plus, beaucoup de fleurs qui nous semblent blanches, sont en réalité ornées de dessins dans la gamme de l’ultra violet ( Hélianthus rigidus, Oenothera biennis ) .
Ils sont capables de percevoir la lumière polarisée.
1.4.8.3 Le cas de l’abeille
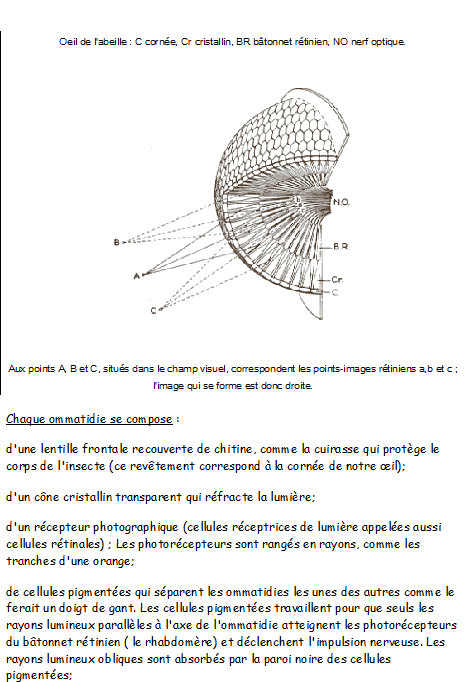
Les abeilles possèdent nos cinq sens et d'autres encore, mais l'organisation et l'anatomie de leurs organes sensoriels sont très différentes des nôtres. Leurs yeux ne sont en rien comparables aux nôtres qui se présentent comme des appareils photographiques miniatures: la lumière pénètre dans l'œil au travers d'une grande lentille, le cristallin, qui focalise l'image sur un film photosensible, la rétine. C'est par son intermédiaire que nous avons conscience des images qui surgissent et s'évanouissent au rythme des mouvements de nos yeux, accompagnées de leurs jeux d'ombres et de lumières.
Les minuscules cellules nerveuses de la rétine agissent pour accentuer certains éléments et structurer l'image, avant de la transmettre au cerveau en réunissant tous ces filets nerveux dans le nerf optique.
Chez presque tous les invertébrés dont l'abeille, les yeux sont à facettes: c'est-à-dire composés de nombreuses petites lentilles ou ommatidies. L'ouvrière possède 4500 facettes par œil; la reine, pour qui la vision est de peu d'utilité n'en a que 3500. Quant au mâle, il est primordial qu'il soit capable de repérer une reine à grande distance, s'il veut être le premier à la féconder: ses yeux ont 7500 facettes chacun.
Chaque ommatidie se présente et fonctionne comme un récepteur visuel indépendant qui capte la partie du champ visuel situé juste devant lui, mais aucune image ne s'y forme: celle-ci est recomposée à partir des informations transmises par l'ensemble des facettes.
Un objet situé dans le champ visuel de l'œil de l'abeille émet des rayons dans toutes les directions, intéresse l'œil dans sa globalité, mais seul le rayon dirigé exactement dans l'axe du rabdomère ou bâtonnet rétinien sera enregistré.
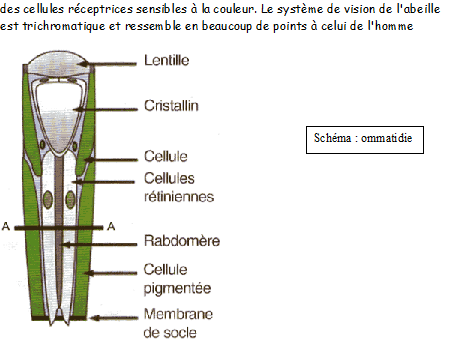
capable de situer l'emplacement et le mouvement d'un objet. Elles ne donnent aucune image, mais permettent à l'insecte volant de stabiliser sa ligne de vol par rapport à l'horizon. On peut dire que le corps en vol est bien stabilisé, quand les deux ocelles supérieures sont bien éclairées et que l'ocelle inférieure qui est dirigée sur l'horizon absorbe moins de lumière. Dans la ruche, les ocelles permettent à l'abeille à s'orienter vers la sortie.
I.4.9 Le système musculaire
Les muscles des insectes sont généralement translucides ou de couleur grise ; à l’exception des muscles des ailes qui présentent une teinte jaune orangée.
Une grande différence avec les muscles des Vertébrés : les fibres des muscles des mouvements volontaires comme celles des mouvement involontaires sont striées.
I.4.9.1 Types de muscles
- Les muscles viscéraux qui recouvrent les organes à l’intérieur de la cavité générale . Ils peuvent former une double couche régulière de fibres longitudinales et circulaires autour du tube digestif avec, à certains niveau, un développement de la couche circulaire en sphincter.

I.4.10 Les organes diffus
Ce sont des éléments dispersés, non rassemblés en un organe : les « organes diffus ».
Les cellules péricardiales et les néphrocytes
Ceux-ci jouent probablement un rôle important dans l’excrétion. Ils sont disposés en amas sur le diaphragme dorsal et au niveau de divers organes dont les glandes salivaires. Doués de capacités d’absorption par pinocytose, ils pourraient participer à la détoxification des substances étrangères introduites dans l’hémocoele.
Les oenocytes
Cellules volumineuses polyploïdes, disposées en amas métamériques au niveau des stigmates , dispersées sous l’épiderme dont elles dérivent et dans le corps gras , leurs fonctions sont mal connues, elles sont riches en réticulum endoplasmique .
Le corps gras ou tissus adipeux
Très développé chez les insectes , il forme une gaine à peu près continue autour du tube digestif ( corps gras interne ). Il comporte des adipocytes , cellules chargées de sphérocristaux , voire de cellules hébergeant des organismes symbiotiques ( levures, bactéries, mycoplasmes).
Date de dernière mise à jour : 05/07/2021
Commentaires
-
 This paragraph is truly a pleasant one it assists new net people, who are wishing for blogging.
This paragraph is truly a pleasant one it assists new net people, who are wishing for blogging. -
 c trés bien d'avoir cette quantité d'information merci à vous
c trés bien d'avoir cette quantité d'information merci à vous
Ajouter un commentaire