
Anatomie comparée des Vertébrés 3
I.4 Les glandes cutanées
1.4.1 Origine et évolution
L'épiderme non kératinisé des Vertébrés inférieurs aquatiques ( Cyclostomes, Poissons, ... ), et des larves d'Amphibiens contiennent de nombreuses cellules glandulaires, isolées, différenciées à partir de l'assise génératrice. L'activité germinative de cette dernière les déplace progressivement vers la surface de l'épiderme où elles crèvent, déversent leur sécrétion et meurent. la plupart élaborent du mucus qui forme un film protecteur continuellement renouvelé, jouant vis-à-vis des micro-organismes parasites ( Champignons,Bactéries... ) . un rôle comparable à celui de la couche cornée des Tétrapodes. Certaines, chez les Téléostéens Cypriniformes, sécrètent une substance d'alarme qui est détectée par l'organe olfactif d'autres individus de la même espèce.
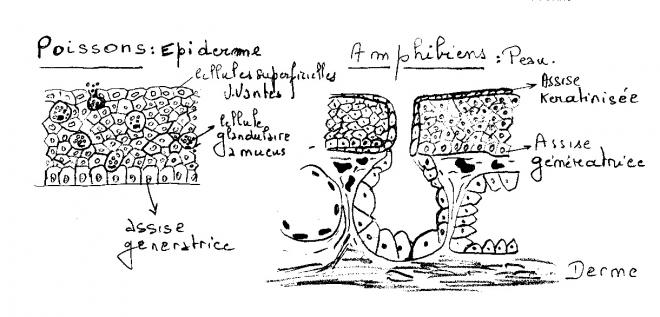
Les cellules glandulaires vont disparaître de l’épiderme kératinisé des Tétrapodes pour se grouper en glandes, invaginations épidermiques de formes variées ( acineuses , tubuleuses ou mixtes) qui s’enfoncent dans le derme et s’ouvrent à l’extérieur par un canal excréteur traversant l’épiderme.
1.4.2 Cas des Amphibiens adultes
Glandes très développées chez les Amphibiens adultes :
- Acini simples muqueux répartis sur toute la surface du corps , ils jouent un rôle essentiel dans l’adaptation aérienne par la sécrétion d’un mucus qui limite l’évaporation, maintient la peau humide et permet la respiration cutanée.
- Acini granuleux à sécrétion venimeuse jouant un rôle défensif vis à vis de la prédation (Amines biogènes irritantes, oligopeptides hypotenseurs,alcaloïdes jouant sur les canaux Na et K de la membrane plasmique des cellules musculaires et nerveuses…)
1.4.3 Sauropsidés
Peu développées et très localisées chez les oiseaux et les Reptiles :
1.4.3.1 Reptiles
- Glandes anales des Serpents,
- glandes cloacales de Sauriens et Crocodiliens ( rapprochement des sexes et marquage des territoires),
- glandes fémorales de nombreux Sauriens ( Iguanes, Lézards, Scinques ) servant à marquer le territoire,
- Glandes mandibulaires des Chéloniens
- Glandes cervicales des Crocodiles
1.4.3.2 Oiseaux
1.4.3.2.1 Préalable
- Glandes mérocrines
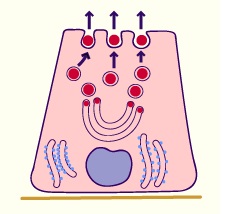
C'est le mode le plus courant. Le produit de sécrétion est relaché par exocytose.
La membrane des vésicules fusionne avec la membrane plasmatique apicale et ainsi les vésicules contenant le produit de sécrétion s'ouvrent à la surface.
La membrane fusionnée retourne dans le cytoplame par endocytose. Elle est recyclée et réutilisée pour d'autres vésicules.
Cette modalité de secrétion est observée par exemple dans les glandes salivaires, le pancréas exocrine (granules de zymogène) et au niveau de la glande mammaire pour la sécrétion de protéine (caséine).
- Glandes holocrines
Lors du cycle sécrétoire, le cytoplasme de la cellule se charge d'une quantité considérable de produit de sécrétion et ensuite se désintègre. En d'autres termes la cellule en mourant devient elle même le produit de sécrétion.
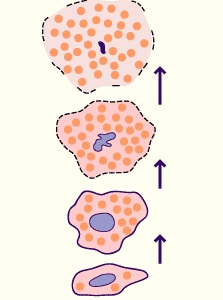
Cette modalité de secrétion est observée par exemple au niveau des glandes sébacées.
- Glandes apocrines
Le produit de sécrétion accumulé au pôle apical est éliminé par apocytose.
La membrane apicale se détache lors de l'extrusion et entoure le produit de sécrétion. La cellule glandulaire conserve cependant son noyau et ses organites. Elle peut ainsi reprendre un cycle sécrétoire.
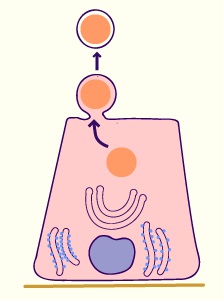
Cette modalité de sécrétion est observée par exemple dans certaines glandes sudoripares et au niveau de la glande mammaire pour la sécrétion de produit lipidique.
1.4.3.2.2 Les glandes cutanées des Oiseaux
Les oiseaux possèdent trois types de glandes à sécrétion holocrine huileuse :
- Les glandes uropygienne paires : à la face dorsale du croupion
- Les glandes du conduit auditif externe
- Les glandes de la face ventrale de l’abdomen
Le peu de glandes cutanées s’explique par le fait que les cellules banales de l’épiderme ont également une fonction exocrine, elles élaborent des lipides au cours de leur kératinisation. Ceux-ci sont libérés par la rupture de la paroi épaisse des Kératinocytes. Cette dernière activité est toujours plus importante dans les régions glabres.
L’épiderme d’un Oiseau fonctionne donc comme une vaste glande holocrine dont la sécrétion huileuse semblable à celle des glandes sébacées des Mammifères protège la peau et offre une barrière efficace à l’eau.
1.4.4 Les Mammifères
Nombreuses et différenciées en trois grandes catégories très spécialisées :
- Glande sébacées.
- Glandes sudoripares.
- Glandes mammaires.
1.4.4.1 Les glandes sébacées
Caractéristiques
- Glandes acineuses simples ou composées
- S’ouvrent à la surface du tégument par l’intermédiaire de l’espace pilo-sébacé qui entoure la racine du poil
- Proviennent du bourgeonnement de la gaine épithéliale externe qui s’enfonce dans le derme et prend la forme d’une poire ou d’une massue.
- La sécrétion est composée de triglycérides, de cholestérol, de cires et de débris cellulaires, c’est le sébum.
- Sécrétion holocrine
Rôle de la sécrétion
- Régularisation des pertes d’eau par évaporation.
- Imperméabilisation.
Glandes sébacées absentes chez les Cétacés.
Assouplissement de la toison du Mouton
Chez l’homme, particulièrement développées autour du nez, de la bouche, au niveau des pommettes, du front, du cou, de la partie antérieure du tronc, sur les organes génitaux.
1.4.4.2 Les glandes sudoripares.
Ce sont des glandes exocrines, tubuleuses simples pelotonnées, sécrétant la sueur. Leur portion sécrétrice (épithélium cubique simple) entourée de cellules myo-épithéliales, siège dans le derme profond. Leur canal excréteur (épithélium cubique bistratifié) gagne la surface de l'épiderme par un trajet hélicoïdal. L'innervation des glandes sudoripares est sympathique, segmentaire.
Les glandes sudoripares se subdivisent en deux groupes :
- Les glandes sudoripares apocrines :
Quasi totalité des glandes sudoripares.
Développement
- Prennent naissance à partir d’un bourgeon de la gaine épithéliale externe des follicules pileux, situé au dessus de l’ébauche de la glande sébacée.
- Le bourgeon s’allonge en un long tube et la partie glomérulaire s’enfonce dans la pannicule adipeux.
Sécrétion
- Sécrétion laiteuse, jaunâtre et parfois rougeâtre ( Hippopotames), cette sécrétion devient rapidement odorante sous l’effet des bactéries.
Chez l’homme adulte, elles subsistent au niveau des téguments, des aisselles, du nombril, des mamelons, de la région ano- génitale et des conduits auditifs externes.
- Les glandes sudoripares mérocrines ou eccrines
- Indépendantes des follicules pileux
- Les glandes sudoripares eccrines, à peu d'exceptions près, sont réparties sur la surface entière du corps
- glandes simples, tubuleuses et enroulées sur elles-mêmes Leur conduit débouche habituellement à la surface de la peau par un pore.
- Plus petites que les précédentes, leur glomérule ne pénètre jamais dans le panicule adipeux sous-jacent
Leur principale fonction consiste à maintenir au même niveau la température de l'organisme (thermorégulation) et a véhiculer la sécrétion des glandes sudoripares apocrines.
Sécrétion
Sécrètent une solution salée contenant de faibles quantités d'autres solutés.
1.4.4.3 Glandes cutanées spécialisées
La plupart des Mammifères possèdent dans des territoires cutanés très localisés des glandes sudoripares ou sébacées ( ou mélange des deux).
Ces glandes sécrétent des substances grasses à odeur souvent très forte, voire infecte.
Leurs rôles se situeraient à divers niveaux :
- Marquage du territoire
- Phéromones
- Reconnaissance individuelle
- Rapports sociaux
- Appétence sexuelle
Exemples :
- Glandes anales des Mustélidés ( odeur fétide )
- Glandes périnéales des Viverridés
- Glandes préputiales des Rongeurs, celles-ci sont de la taille d’une petite orange chez le Chevrotain porte musc et dont la sécrétion désséchée constitue le musc.
Et bien d’autres encore.
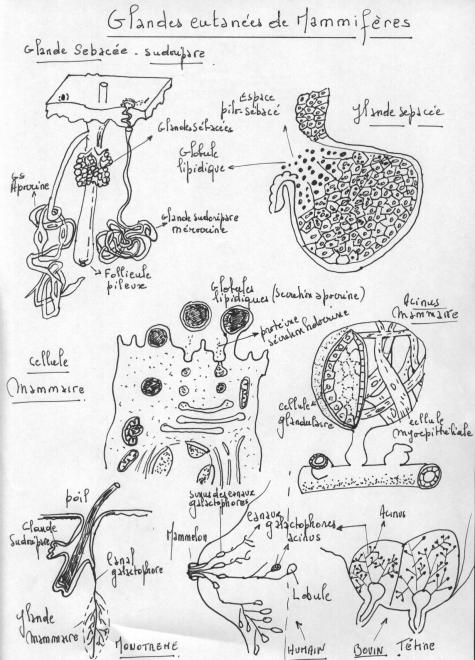
1.4.4.4 Les glandes mammaires
Glandes tubulo-acineuses composées, responsables de la production de lait. Elles sont toujours présentes chez les Mammifères.
1.4.4.4.1 Anatomie
Son architecture est schématiquement superposable à celle des autres glandes exocrines composées.
On y trouve :
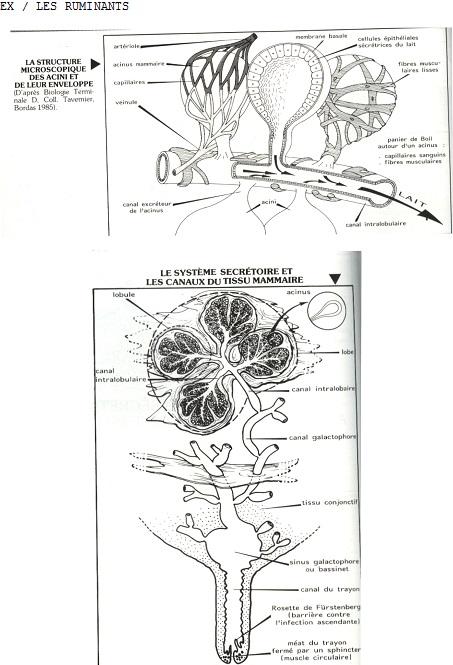
- un système ramifié de canaux excréteurs terminés en cul-de-sac par des portions sécrétrices tubulo-alvéolaires ( acini).
- Chaque glande mammaire est constituée de un (ruminants) plusieurs lobes drainés ( femme ) par des canaux galactophores collecteurs « lactifères » s'ouvrant individuellement à la peau au niveau du mamelon.
- Chaque lobe est lui-même constitué de lobules, chaque lobule étant constitué de tubulo-alvéoles correspondant à la partie sécrétrice de la glande.
- Les lobes sont séparés par du tissu conjonctif dense et entourés par du tissu adipeux abondant, de même les lobules sont entourés par du tissu conjonctif dense alors que le tissu conjonctif intralobulaire (tissu conjonctif palléal) est lâche.
- Les canaux excréteurs (ou canaux galactophores) sont d'abord intralobulaires, puis interlobulaires (épithélium cubique avec présence de cellules myoépithéliales) et enfin interlobaires à épithélium pavimenteux stratifié.
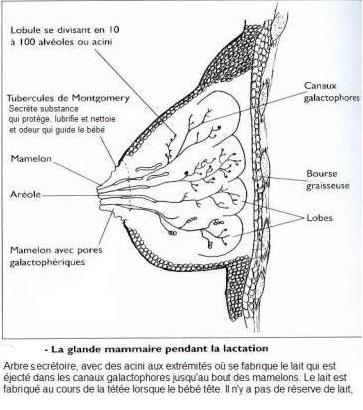
Il est à noter, que chez les ruminant, il n’y a qu’un lobe par mamelle
1.4.4.4.2 Développement
- Développement très lent : ébauché dans la vie fœtale, il ne s’achève qu’avec la première gestation. Il peut se partager en trois étapes :
Etape prénatale
Elle aboutit dans les deux sexes, à la mise en place de glandes mammaires réduites à un court système de tubules, par prolifération dans le mésenchyme de bourgeons plein à partir de l’épaississement localisés de la ligne mammaire.
Etape pré pubertaire
Après la naissance, les glandes du mâle ne subissent qu’un développement très limité. Chez la femelle , elles s’accroissent d’abord lentement puis plus rapidement à l’approche de la puberté suite à l’action des hormones ovariennes.
Gestation
Après la gestation, les glandes atteignent leur plein développement : elles s’hypertrophient et les acini se différencient aux extrémités des tubules . Une activité sécrétrice se manifeste en fin de gestation sous l’action de la prolactine hypophysaire. Après la gestation, le maintien de cette sécrétion dépend du stimulus de la têtée qui entretient la sécrétion de prolactine par un réflexe neuro-hormonal.
1.5 Formations squelettiques dermiques
1.5.1 Généralités
Le derme des Vertébrés peut se minéraliser de diverses façons et donner naissance a des formations squelettiques souvent lamellaires constituant un exosquelette superficiel plus ou moins développé.Un exosquelette à peu près complet protégeait le corps des premiers Vertébrés du Silurien et du Dévonien :
- Ostéostracés
- Hétérostracés
- Placodermes
Il a subit une importante évolution régressive et ne subsiste plus, chez la plupart des Tétrapodes actuels, que sous la forme d’un petit nombre d’éléments incorporés dans le squelette crânien et zonal.Indépendamment de cette régression, un développement secondaire d’éléments exosquelettiques est apparu chez divers Reptiles et Mammifères, allant même dans certains cas jusqu’à reconstituer, par une curieuse convergence, une carapace presque aussi complète que celle de leurs ancêtres du primaire.
1.5.2 Les différents tissus squelettiques d’origine dermique
Le degré de minéralisation de la substance fondamentale conjonctive, la forme des cellules conjonctives et leur disposition par rapport à cette substance fondamentale permettent de définir trois grands types de tissus squelettiques d’origine dermique :
- Les tissus osseux.
- Les dentines.
- Les émailoïdes.
1.5.2.1 Le tissus osseux
Le plus classique et le moins minéralisé des tissus exosquelettiques ( 60 à 70 % éléments minéraux ). Ses cellules, étoilées, les ostéocytes, sont incluses dans la substance fondamentale qu’elles ont élaborée.
1.5.2.2 Les dentines
Très diverses, elles sont aussi plus riches en sels minéraux ( 70 à 75%), et leurs cellules, les odontoblastes sont extérieures à la substance fondamentale dans laquelle elles envoient un long prolongement apical.
1.5.2.3 Les émailoïdes
- Hyperminéralisés ( 90 à 95 % minéraux ).
- Croissance centripède avant le dépôt de dentine
- Matrice de collagène avec cellules épidermiques suggérant une origine mixte dermo-épidermique.
- Ne pas confondre avec l’émail d’origine épidermique et à croissance centrifuge après le dépôt de dentine.
La ganoïne à structure lamellaire classiquement considérée comme émailoïde pourrait être un émail vrai.
1.5.3 L’exosquelette des Ancêtres des Vertébrés et sa régression
Les premiers Vertébrés pisciformes du Dévonien et de Silurien ( Ostéostracés, Hétérostracés, Placodermes) étaient à peu près complètement recouverts d’une armure exosquelettique , véritable cuirasse faite d’écailles articulées, fusionnées en l’avant du corps en un vaste bouclier céphalothoracique . Le développement de cet exosquelette leur a valu le nom de « Poissons cuirassés ». L’armature exosquelettique de ces Poissons cuirassés est généralement composée de trois couches superposées :
- Une couche profonde d’os lamelleux
- Une couche moyenne d’os spongieux
- Une couche externe faite de tubercules de dentine formant l’ornementation des écailles et des plaques.
- Très souvent, ces tubercules sont recouvert d’une mince couche d’émailoïde.
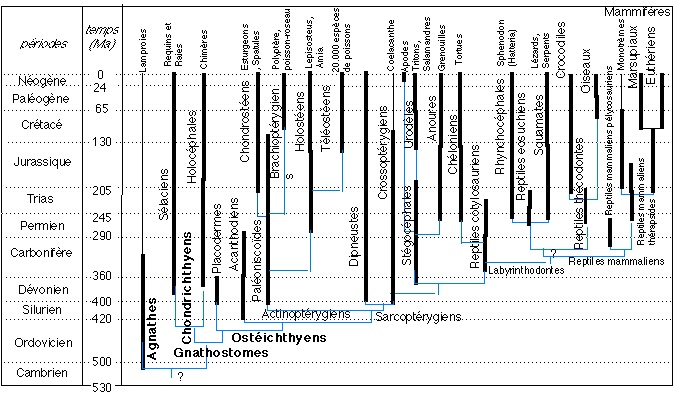
Au cours de l’évolution, cet exosquelette s’est considérablement allégé par régression de son épaisseur et de son extension, tandis qu’inversement l’endosquelette se développait.
Chez les Vertébrés actuel, il n’est plus représenté que par les écailles des Poissons et par des os dermiques qui participent à la constitution du squelette céphalique et de la ceinture pectorale des Gnathostomes.
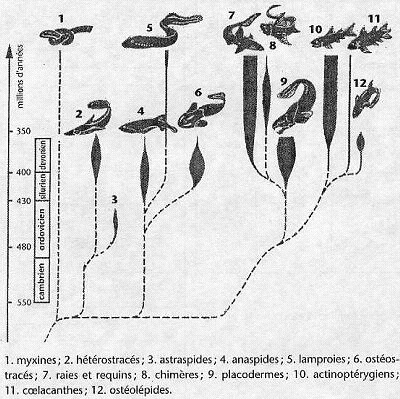
1.5.4 Les écailles des Ostéichthyens
1.5.4.1 Caractéristiques
- Plaques exosquelettiques permanentes, articulées ou imbriquées à croissance continue.
- Elles sont constituées par des tissus exosquelettiques variés parmi lesquels le tissus osseux est toujours présent.
- Leur évolution s’est faite dans le sens d’une régression à partir des plaques dermiques des Placodermes du Dévonien.
1.5.4.2 Types
Deux grands types d’écailles caractérisent les deux grandes lignées d’Ostéichthyens :
1) Les écailles cosmoïdes propres à la lignée sarcoptérygienne
2) Les écailles ganoïdes propres à la lignée actinoptérygienne
1.5.4.3 Les écailles cosmoïdes et leurs dérivés
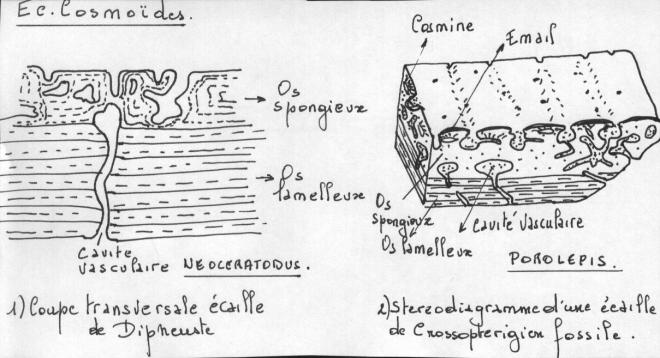
Une écaille cosmoïde se compose de quatre couches superposées :
1) Une épaisse plaque basale d’os lamelleux : l’isopédine
2) Une couche d’os spongieux pénétré par de nombreuses cavités vasculaires en réseau.
3) Une couche de dentine de structure particulière ou cosmine, faite d’une série de tubercules. Ces tubercules sont séparés par une cavité pulpaire d’où s’irradient de nombreux tubules très fins contenant les prolongements cytoplasmiques des Odontoblastes La cavité pulpaire communique avec les cavités vasculaires de l’os spongieux.
4) Une très mince couche superficielle très dure d’émail recouvrant les tubercules.
Les écailles cosmoïdes s’accroissent en profondeur par l’addition de nouvelles couches d’isopédine à la face profonde et en surface par la formation de nouveaux tubercules sur le bord antérieur.Les grosses écailles du coelacanthe ( seul Crossoptérygien actuel )recouvrent la totalité du corps et conservent encore quelques tubercules de dentine recouverts d’émail.Cest tubercules sont complètement disparus chez les dipneustes actuels.
1.5.4.4 Les écailles ganoïdes et leurs dérivés
( Lignée actinoptérygienne )
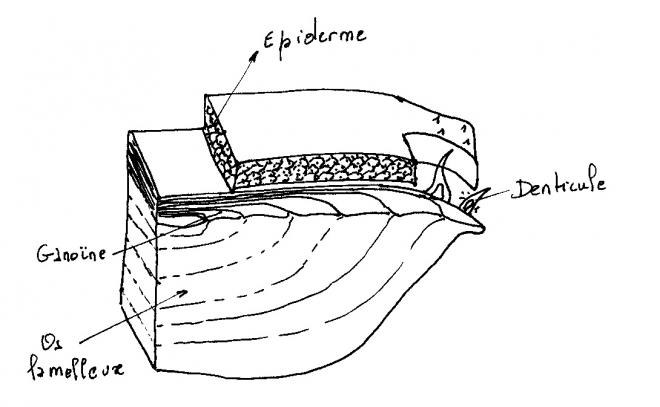
Sont trouvées sous leur forme typique chez les premiers Actinoptérygiens du Primaire, les paléoniscoïdes, ce sont de grandes écailles épaisses, rhombiques assez semblables aux écailles cosmoïdes. Elles en diffèrent toutefois par l’absence d’os spongieux et les remplacement de la fine couche superficielle d’émaïl par plusieurs couches superficielles de ganoïne.De véritables écailles ganoïdes ne persistent actuellement que chez les Polyptères ( Brachioptérygiens ) et Lepisosteus ( Holostéens ).
Les Polyptères sont complètement recouverts d’épaisses écailles articulées ayant conservé une structure primitive.
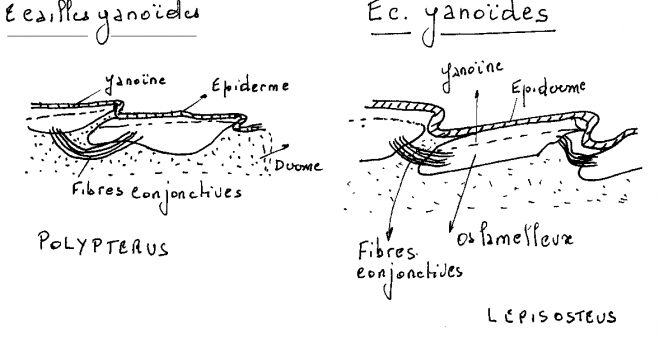
La ganoïne , épaisse d’une cinquantaine de microns est présente uniquement sur la partie découverte des écailles. Elle est percée de nombreux pores par lesquels s’ouvre le réseau de canaux de la dentine. Ce réseau communique également avec les couches profondes du derme par quelques lacunes et canaux creusés dans la couche d’isopédine.
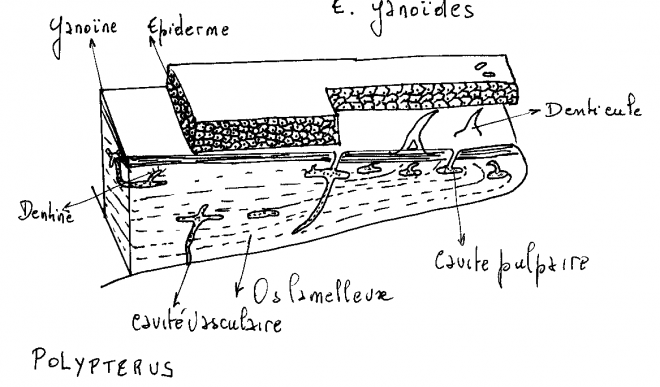
La ganoïne , épaisse d’une cinquantaine de microns est présente uniquement sur la partie découverte des écailles.
 LEPIDOSTEUS
LEPIDOSTEUS
Chez Lepisosteus, l’os lamelleux est recouvert sur ses parties exposées d’une couche de plus ou moins cent microns de ganoïne. La ganoïne porte en surface, de petits tubercules.La dentine et la ganöine disparaissent chez tous les autres Actinoptérygiens. Les écailles réduites à l’os lamelleux s’amincissent.Chez les Téléostéens, elles deviennent des lamelles transparentes ( écailles élasmoïdes ) recouvertes d’une couche superficielle ornementée faite le plus souvent d’un tissus osseux acellulaire.
1.5.4.5 Les écailles placoïdes des Chondrichtyens
Contrairement au Ostéichtyens, remplacement continu et limité.
- Eruption par percement de l’épiderme
- Cavité pulpaire et soudure à une socle acellulaire.
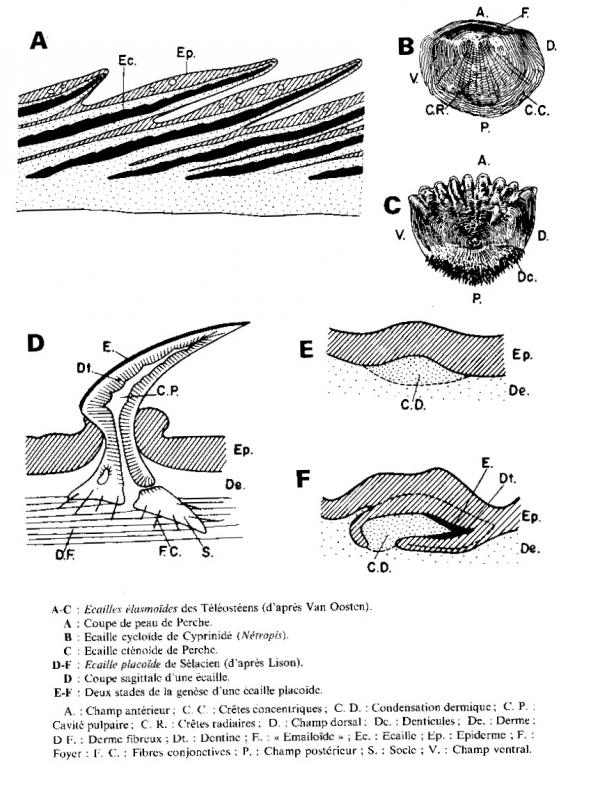
Ces ont des dents cutanées constituées essentiellement constituées par un cône de dentine recouvert par un mince vernis d’émailoïde .
1.5.4.6 Les écailles osseuses des Amphibiens
Le corps des premiers Amphibiens du primaire, les Stégocéphales , est souvent recouvert sur la face ventrale , entre les ceintures de rangées d’écailles osseuses disposées en V. Parfois ces écailles fusionnent dans une même rangée et forment des gastralia.Ces gastralia sont assez fréquentes chez les Reptiles fossiles et sont encore présentes chez l’Archéoptéryx
Chez les Reptiles actuels, ces gastralia ne persistent que chez les Rhynchocéphales ( 24 à 26 gastralia ) et chez les Crocodiliens ( 7 à 8 ) ou elles sont improprement appelées côtes abdominales ou parasternum.
Les petites écailles dermiques de certains Amphibiens apodes ( Ichtyophys de l’Inde, Coecilie de Guyanne, Hypogéophis des îles Seychelles ) sont généralement considérées comme homologues des écailles de Stégocéphales.
1.5.4.7 Les os dermiques du crâne et la ceinture pectorale
Une partie de la cuirasse exosquelettique des premiers Vertébrés s’est incorporée dans le squelette du crâne ( neurocrâne et splanchnocrâne). Et de la ceinture pectorale.
1.5.4.8 Développement secondaire d’un exosquelette dermique
Bien que l’exosquelette primitif des ancêtres des Vertébrés ait considérablement régressé au cours de l’évolution, le derme a cependant conservé sa potentialité à s’ossifier, et chez divers Vertébrés actuels ou fossiles est apparu un développement secondaire d’os dermiques.
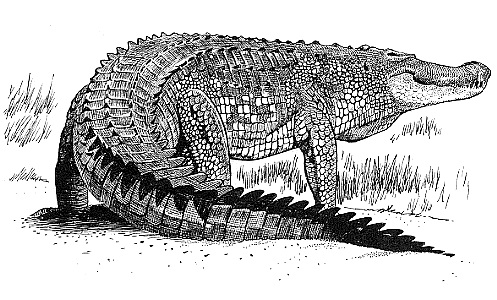
- Les Ostéodermes des Reptiles
Les ostéodermes des Reptiles actuels ( à l’exception des Ophidiens et des Chéloniens ) sont généralement de petites formations osseuses lamellaires distribuées en certaines régions du tégument : dessus de la tête, du tronc et de la queue. Chez les Crocodiliens, ils sont mieux développés et deviennent de grandes plaques épaisses, plus ou moins quadrangulaires à bord dentelé, dont la surface peut se soulever en une carène médiane.
Les Stégosaures, reptiles Dinosauriens du Jurassique , portaient deux rangées dorsales parasagittales de grandes plaques ou de longue épines osseuses dressées au dessus des apophyses neurales des Vertébres.

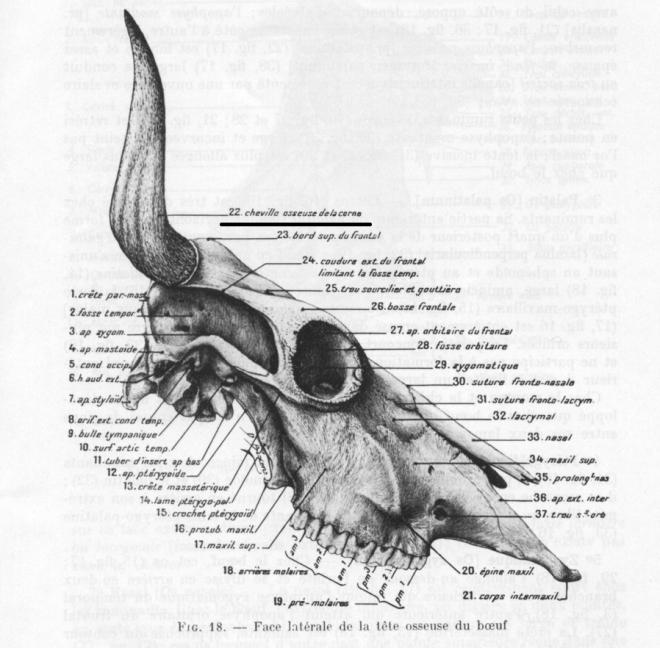
- Les os cornus ( ou chevilles osseuses )
Les os cornus des ruminants cavicornes se soudent précocement au frontal et constituent l’axe osseux souvent pneumatisé autour duquel se différencie l’étui corné épidermique.
Ex cheville osseuse de la corne de boeuf
Des os cornus de petite taille ( 10 à 15 cm ) se développent chez les Girafes des deux sexes. Ils se soudent tardivement au frontal et sont recouverts d’un peau velue avec une touffe de crin à leur extrémité. Chez l’Okapi, les cornillons sont au nombre d’une paire et ne sont présents que chez le mâle.
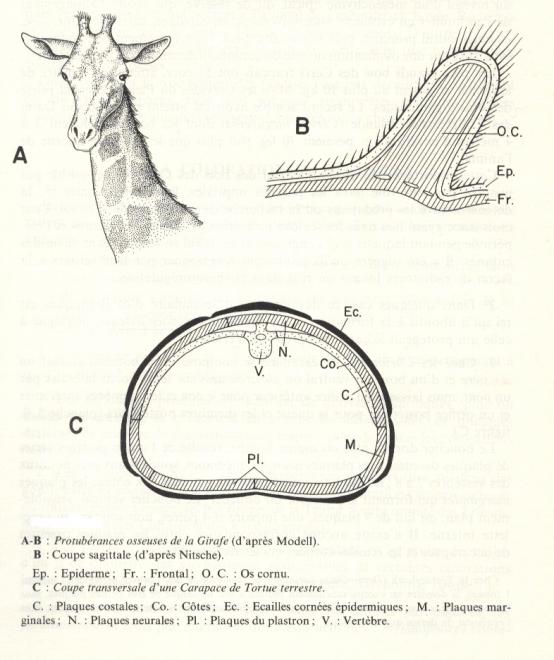
Les bois des Ruminants de la famille des Cervidés
Cerfs, Chevreuils, Elans, Daims, Rennes,…
- Souvent confondus avec des cornes.
- Homologues des os cornus.
- Caducs, ils repoussent chaque année.
- Ils sont recouverts d’un épiderme qui ne se différencie jamais en étui corné.
- Ils ne se développent que chez le mâle, sauf chez le Renne.
L’ossification des bois de Cervidés est un processus complexe qui commence au niveau d’un mésenchyme apical dit « de réserve » qui assure l’allongement de l’andouiller en évoluant, successivement, en cartilage, cartilage minéralisé, os enchondral primaire puis secondaire.

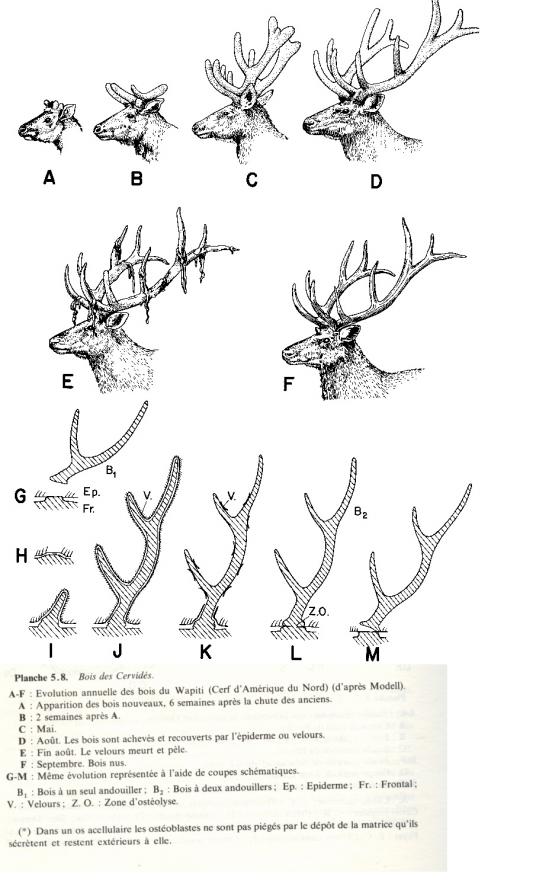
- Carapaces osseuses
Carapaces des Chéloniens :
- Un bouclier dorsal ( dossière)
- Un bouclier ventral ( plastron)
- Dossière et plastron unis sur leurs bords latéraux par un pont.
Orifices :
- Antérieur pour tête et pattes
- Postérieur pour queue et pattes.
Carapace des tatous actuels :
Composée de trois boucliers, céphalique, scapulaire et pelvien. Boucliers scapulaire et pelvien sont séparés par 2 à 13 bandes transversales s’interrompant ventralement pour permettre à l’animal de se rouler en boule.
Bandes et boucliers sont constitués de petites plaques osseuses soudées.
D’autres plaques osseuses indépendantes recouvrent le coyu, la queue, ainsi que toute la face ventrale et les pattes.
Les plaques osseuses sont toujours recouvertes d’écailles cornées épidermiques.
1.5.4.9 La coloration du tégument
La couleur d’un objet est due à l’absorption par celui-ci de certaines des radiations de la lumière blanche incidente . Dans le cas d’un animal, cette absorption peut être le fait :
- De la nature chimique de certaines substances ou pigments situés dans le tégument : colorations pigmentaires
- De la nature physique de la surface du tégument : colorations structurales
- Ou des deux à la fois : colorations mixtes
1° colorations pigmentaires
Les pigments des Vertébrés appartiennent à trois familles chimiques principales :
- Les mélanines : polymères d’oxydation des dérivés phénoliques, en grains de quelques microns de diamètre : les mélanosomes. Colorations brunes ou noires.
- Les caroténoïdes : hydrocarbures terpéniques toujours dissous dans des gouttelettes lipidiques, ils sont à l’origine de certaines colorations jaunes, rouges ou oranges , Poissons rouges, ventre des tritons, plumage jaune des Canaris…
- Les ptérines : Hétérocycles azotés, se présentent en granules de moins de un micron de diamètre , les ptérinosomes, responsables de certaines colorations jaunes ou rouges ( taches jaunes de la Salamandre, Grenouille rousse).
Ces pigments sont toujours élaborés dans des cellules particulières , les chromatophores issus par migration des crêtes neurales : mélanophores contenant des mélanosomes, lipophores contenant des caroténoïdes , ptérinophores contenant des ptérinosomes. Les chromatophores sont localisés en principe dans le derme , mais peuvent éventuellement s’infiltrer entre les cellules épidermiques . Les pigments peuvent rester dans les chromatophores qui les ont élaboré ( Poissons , Amphibiens, Reptiles), ou être excrétés dans des cellules épidermiques en voie de kératinisation ( plumes des Oiseaux et poils des Mammifères ).
2° Colorations structurales
Elles peuvent résulter :
a) Des phénomènes de dispersion de la lumière au niveau des microvacuoles ou des micelles colloïdaux . Selon la taille des microstructures, la dispersion est totale et la coloration est blanche ( plumes et poils blancs ) ou partielle et la coloration est bleue par effet Tyndall ( iris de l’homme, plumes de la mésange…)
b) De phénomènes d’interférence entre les lamelles de kératine de la couche cornée ( plumes de Paon) ou entre les faces paralléles de microcristaux de guanine et d’hypoxanthine ( poissons) de purines et d’adénine (Amphibiens) par eux même incolores , contenus dans des cellules dermiques particulières, les iridophores particulièrement abondants dans la peau des poissons , des Amphibiens et de nombreux Reptiles . Ces couleurs sont toujours irisées, c’est à dire qu’elles varient suivant l’incidence de la lumière.
3° Colorations mixtes
Elles sont dues à la combinaison d’une coloration pigmentaire et d’une coloration structurale . Ainsi , la coloration verte de nombreux Vertébrés ( Rainette, Grenouille verte , Lézard vert, Perroquet, Perruches…) n’est pas due à un pigment vert, mais à une coloration structurale bleue par effet Tyndall combinée à une coloration pigmentaire jaune due dans la plupart des cas à des caroténoïdes
4° Changements de coloration.
Quelques Vertébrés hétérothermes sont capables de changer leur coloration , parfois de façon rapide et spectaculaire : nombreux Poissons ( Carrelet, Fundulus), quelques Amphibiens ( Grenouilles ) et Reptiles ( Caméléon, Lézard arlequin). Ces changements de coloration sont dus essentiellement à la morphologie et à la physiologie très particulières des mélanophores.
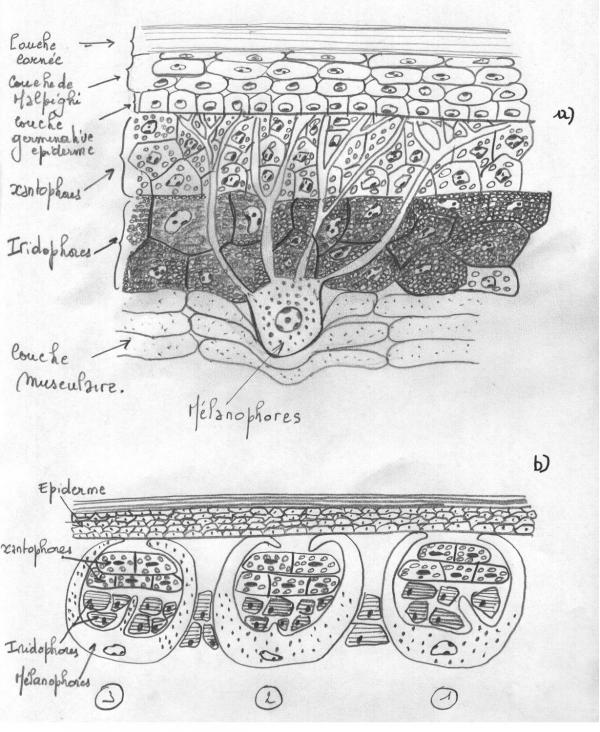

Les mélanophores sont des cellules étoilées et branchues à allure de neurone , capable de modifier la répartition des grains de mélanine dans leur cytoplasme. Leur corps cellulaire est généralement situé sous une couche superficielle de guanophores et de chromatophores à pigment jaune ( lipophores, ptérinophores , xantophores ) entre lesquels s’insinuent jusque l’épiderme leurs prolongements arborescents . Quand les grains de mélanine sont regroupés dans le corps cellulaire profond , les sources de coloration bleue ou jaune sont démaquées. Au contraire, quand le pigment mélanique migre vers l’épiderme, la coloration verte disparaît et est remplacée progressivement par une coloration brune
Ces migrations des mélanosomes se font par l’intermédiaire de microtubules abondant dans les mélanophores.
Ces changements de répartition des grains de mélanine ont pour point de départ des informations visuelles qui atteignent l’encéphale par le nerfs optique . Ces informations sont transmises aux mélanophores par voie nerveuse ( Caméléon), par voie hormonale ( Amphibiens ) soit par les deux voies ( Anguilles).
Au moins trois types d’hormones interviennent :
- Hormone mélanophoro stimulantes provenant de la pars intermédia de l’hypophyse ( M.S.H.) qui dispersent les mélanosomes et assombrissent le tégument
- L’adrénaline et la mélatonine ( d’origine épiphysaire) qui concentrent les mélanosomes et éclaircissent le tégument ou démasquent d’autres sources de coloration.
Les rôles de ces changement de coloration sont :
- Homochromie avec le milieu
- Protection contre l’éclairement
- Thermorégulation.
Date de dernière mise à jour : 30/03/2018
Ajouter un commentaire