
Anatomie 1 Phanérogames
Chapitre VI. ANATOMIE DES PHANEROGAMES
VI.1 CELLULES ET TISSUS DE LA PLANTE
VI.1.1 Méristème apicaux
Pour comprendre le fonctionnement des méristèmes apicaux, nous devons considérer les modifications qui surviennent dans le temps et dans l’espace. Lorsque les cellules du méristème se divisent, elles produisent d’autres cellules méristématiques ainsi que des cellules filles qui s’accumulent, s’allongent et poussent les méristèmes vers l’avant. Ces cellules filles sont « abandonnées » par les méristèmes et commencent à se modifier – elles se différencient- d’abord partiellement ( méristèmes) primaires et ensuite pour constituer les tissus adultes de la plante.
Méristèmes apicaux et leurs dérivés
Situation des méristèmes apicaux :
- Extrémité des racines .
- Extrémité des tiges.
Rôle : primordial dans la croissance de la plante
Le terme « méristème apical » s’applique à un ensemble de cellules contenant les initiales et leur descendance immédiate.
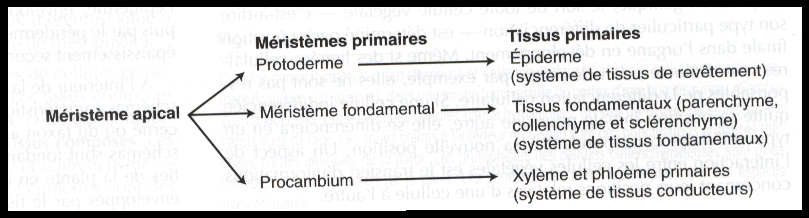
Les initiales sont des cellules qui se divisent en deux cellules filles dont l’une reste une initiale du méristème tandis que l’autre devient une nouvelle cellule végétative de la plante, ou cellule dérivée. Les cellules dérivées peuvent encore se diviser à proximité de la pointe de la racine ou de la tige avant de se différencier. Les divisions ne se limitent cependant pas aux initiales et aux cellules qui en dérivent immédiatement Par exemple ce que l’on appelle les méristèmes primaires – protoderme, procambium et méristème fondamental- apparaissent au cours de l’embryogenèse et s’étendent dans toute la plante grâce à l’activité des
méristèmes apicaux , Ces méristèmes primaires sont des tissus partiellement différenciés qui restent encore quelque temps méristématique avant d’entamer leur différenciation en cellules spécialisées dans les tissus primaires
Ce mode de croissance qui implique l’extension de la plante et la production de tissus primaires est appelée croissance primaire, et la partie de la plante formée de ces tissus et la structure primaire. Nous évoquerons plus loin et en détail la croissance en
largeur ou croissance secondaire,génératrice de la structure secondaire.
VI.1.2 Croissance – morphogenèse – différenciation
Le développement est l’ensemble de tous les évènements qui conduisent progressivement à l’édification de l’organisme, il implique trois processus qui superposent le croissance, la morphogenèse et la différenciation . Le développement dépend de l’information génétique mais est aussi tributaire es facteurs du milieu.
La croissance est une augmentation irréversible de la taille ; elle provient de l’effet conjoint de la division des cellules et de l’augmentation de leur taille. La division cellulaire elle – même ne constitue pas une croissance. Elle peut simplement augmenter le nombre de cellules sans accroître le volume global de la structure. L’apport de nouvelles cellules à l’organisme par les divisions cellulaires du méristème augmente sa capacité de croissance , mais la plus grande partie du gain en volume provient de
l’élongation cellulaire .
La morphologie de la plante et son organisation interne sont fortement influencées à la fois par la multiplication des cellules et par l’augmentation de leur taille. Les racines par exemple ont normalement une forme cylindrique parce que la plupart des cellules dérivées du méristème apical se divisent transversalement, suivant un plan perpendiculaire au grand axe de la racine. Il en résulte de longues files, ou lignées cellulaires qui continuent à s’allonger parallèlement au grand axe de la racine. La réalisation d’une conformation particulière est appelée morphogenèse.
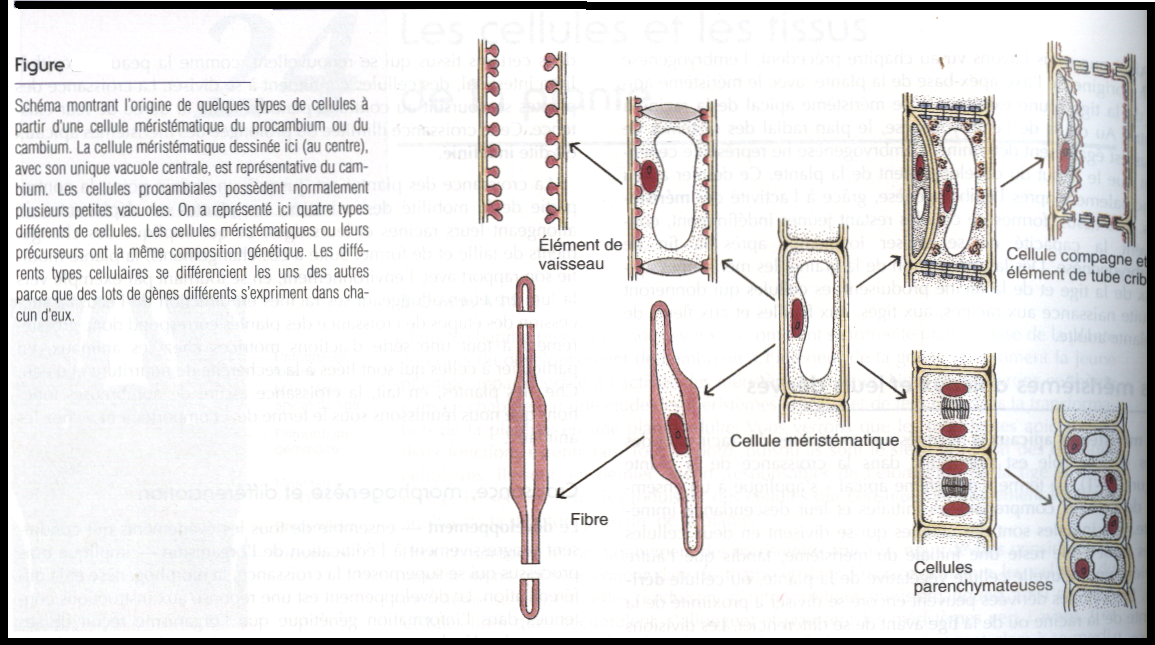
La différenciation – processus par lequel des cellules possédant une information génétique identiques deviennent différentes les unes des autres et des cellules méristématiques dont elles proviennent- débute souvent alors que la croissance de la cellule n’est pas encore terminée . La différenciation cellulaire est soumise au contrôle de l’expression génique, Les différents types de cellules et de tissus synthétisent des protéines différentes parce que les lots de gènes qui s’expriment dans les cellules et tissus ne se sont pas les mêmes que dans les autres,bien que la différenciation cellulaire dépende du contrôle de l’expression génique . le type particulier de différenciation de toute cellule végétale est déterminé par sa position finale dans l’organe en développement ( transfert d’information entre cellules concernant leurs positions relative d’une cellule à l’autre).
VI.1.3 Organisation interne de la plante
Les cellules s’assemblent pour former de vastes ensembles cohérents appelés tissus, au sein de la plante.
Les principaux tissus des plantes vasculaires sont en outre réunis en unités plus grandes en fonction de leur continuité dans l’ensemble de la plante. Ces unités plus grandes ou systèmes de tissus, se reconnaissent aisément, souvent à l’oeuil nu. Il existe trois systèmes de tissus :
- les tissus de revêtement
- les tissus conducteurs
- les tissus fondamentaux
Dans l’embryon, leurs précurseurs sont les méristèmes primaires, respectivement :
-le protoderme
-le procambium
-le méristème fondamental.
Les tissus fondamentaux sont de trois
types :
- le parenchyme, le plus représenté
- le collenchyme
- le sclérenchyme
Les tissus conducteurs sont de deux types :
- le xylème (sève minérale)
- le phloème (sève élaborée)
Les tissus de revêtement sont représentés par :
- l’épiderme, enveloppe des tissus primaires
- le périderme , au niveau des épaississements secondaires
Les tissus sont des ensembles de cellules possédant la même structure et/ou la même fonction.
Les tissus composés d’un sel type de cellules sont les tissus simples ( tissus fondamentaux)
Les tissus composés de divers types cellulaires sont les tissus composés( tissus conducteurs, de revêtement).
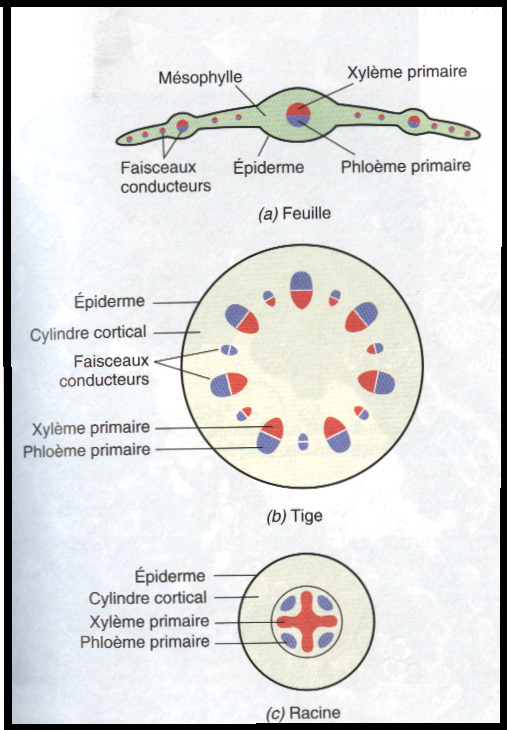
A l’intérieur de la plante les différents tissus sont répartis selon des schémas caractéristiques qui dépendent de l’organe de la plante concerné ou du taxon auquel elle appartient, ou des deux à la fois. Ces schémas sont fondamentalement les mêmes dans les différentes parties de la plante en ce sens que les tissus conducteurs sont toujours enveloppés par le tissus fondamental, le tissus de revêtement formant l’assise externe. Les principales différences de répartition dépendent pour une grande part de la distribution relative des tissus conducteurs et fondamental.Dans la tige des dicotylées par exemple , le tissus conducteur peut former un ensemble de faisceaux interconnectés entourés par le tissus fondamental , la région située à l’intérieur des faisceaux est appelée moelle et la région externe est le cylindre cortical et la région externe est le cylindre cortical. Dans la racine de la même plante , les tissus conducteurs peuvent former un cylindre plein ( stèle) entouré par un cylindre cortical . Dans la feuille, le cylindre conducteur forme normalement une ensemble de faisceaux (nervures) situé à l’intérieur d’un tissus fondamental photosynthétique ( lemésophylle).
VI.1.4 Les tissus fondamentaux
Voir point IV Histologie
Compléments au point IV
En résumé :
On peut concevoir très schématiquement qu’un végétal est composé à ses extrémités d’un méristème terminal orienté vers le haut ( le sommet végétatif d’un bourgeon terminal), et d’un second méristème terminal situé à l’extrémité inférieure de la plante ( la coiffe de la racine principale)
Le but auquel souscrivent ces deux méristèmes vise à faire grandir la plante par l’élongation de ses organes.
Par définition, toute cellule, donc tout tissus et tout organe qui provient du développement de l’un de ces deux méristèmes
d’élongation appartient à la structure anatomique primaire.
En relation avec son allongement, la plante développe latéralement des méristèmes dits secondaires ( par exemple le cambium qui lui permet de s’épaissir). L’épaississement consolide la plante et suit généralement son élongation.
Par définition : toute cellule, tout tissus, toute structure issue d’un méristème d’épaississement appartient à la structure
anatomique secondaire ( ex : le bois)
La structure primaire se rencontre dans tous les organes : tiges, racines , bourgeons , feuilles. Elle est la seule présente au stade juvénile ( plantule) ainsi que dans les organes tendres ( feuilles, fleurs…)
La structure secondaire est limitée à certains organes tels que tiges et racines . Elle prend progressivement une importance quantitative dans une plante adulte.
VI.2 LA RACINE
VI.2.1 La pointe des racines est recouverte par une coiffe
C’est une masse de parenchyme formant un « dé à coudre » autour de la pointe de la racine
- Protection du méristème apical sous jacent
- favorise la pénétration de la racine dans le sol
A mesure que la racine s’allonge et que sa coiffe est poussée en avant, les cellules périphériques de la coiffe se détachent. Ces cellules détruites et la coiffe en croissance sont couvertes d’une gaine mucilagineuse, ou mucigel, qui lubrifie la racine au cours de son passage dans le sol
VI.2.2 Organisation apicale
Caractère structural frappant : disposition de files longitudinales, ou lignées cellulaires qui dérivent du méristème apical.
Le méristème apical comprend un nombre restreint de cellules polygonales nommées initiales et les cellules qui en dérivent immédiatement
Organisation et nombre d’assises d’initiales assez variables dans les méristèmes apicaux des racines
Deux types d’organisation apicale chez les Angiospermes :
- type fermé : Coiffe, cylindre conducteur et cylindre cortical dérivent d’assises cellulaires bien individualisées et indépendantes du méristème apical, l’épiderme ayant la même origine que la coiffe ou le cylindre cortical. Chacune des trois régions possède ses initiales .
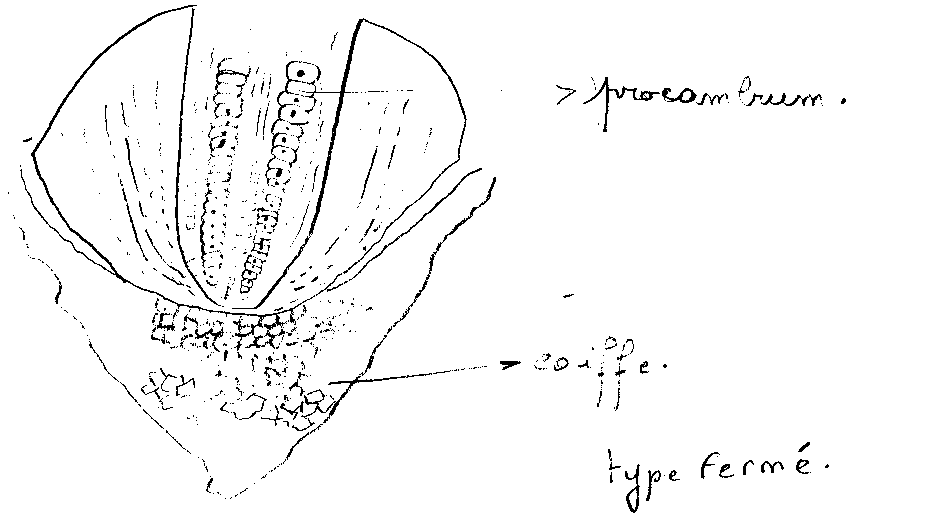
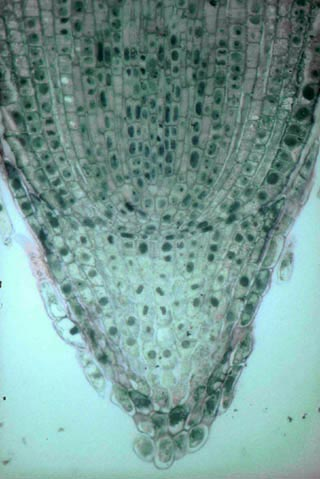
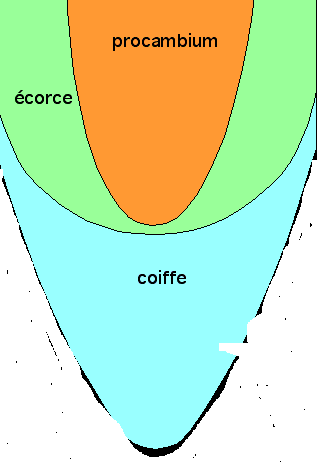
Coupe longitudinale d'un méristème apical racinaire. Dans cet exemple, on distingue trois régions principales indépendantes : la coiffe, le procambium et l'écorce. Ces trois régions sont initiées par des assises de cellules méristématiques différentes. Ici, le rhizoderme est initié par les mêmes cellules que la coiffe.
- type ouvert : Toutes les régions ou du moins le cortex et la coiffe , convergent vers un seul groupe de cellules, toutes les régions possèdent des initiales communes.
Au cours de la croissance les mitoses des cellules situées dans la partie du méristème apical ou se trouvent les initiales deviennent plus rares, la plupart des divisions cellulaires ont alors lieu à courte distance des initiales quiescentes. On appelle centre quiescent la région relativement inactive du méristème apical.
Le centre quiescent aurait un rôle essentiel dans l’organisation et le développement de la racine.
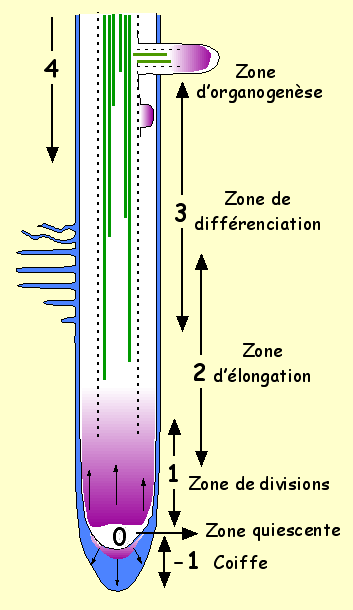
VI .2.3 Structure primaire de la racine
Soit une racine de plante dicotylée
Rappel : coupe longitudinale axiale avec respectivement et de bas en haut : les régions de la coiffe, région lisse,
région pilifère, région subéreuse
Voyons à présent une coupe transversale vers l’extrémité supérieure de la région pilifère.
Nous y distinguons deux cercles concentriques. Replacé en pensée dans la racine , ces deux cercles sont en réalité les sections de deux cylindres,l’un formant manchon autour de l’autre.
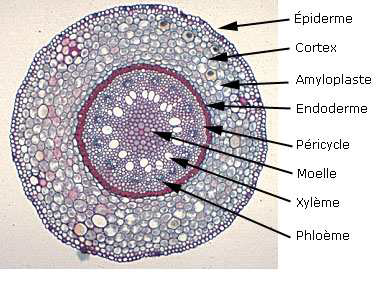
Le cylindre intérieur est dénommé cylindre central ou stèle, il contient des cellules plus ou moins colorées et orientées.
Le cylindre formant le manchon externe est appelé cylindre cortical ou cortex ; il contient généralement des tissus plus tendres que le précédent et se termine à l’extérieur par des poils absorbants.

COMPOSITION DU CYLINDRE CORTICAL
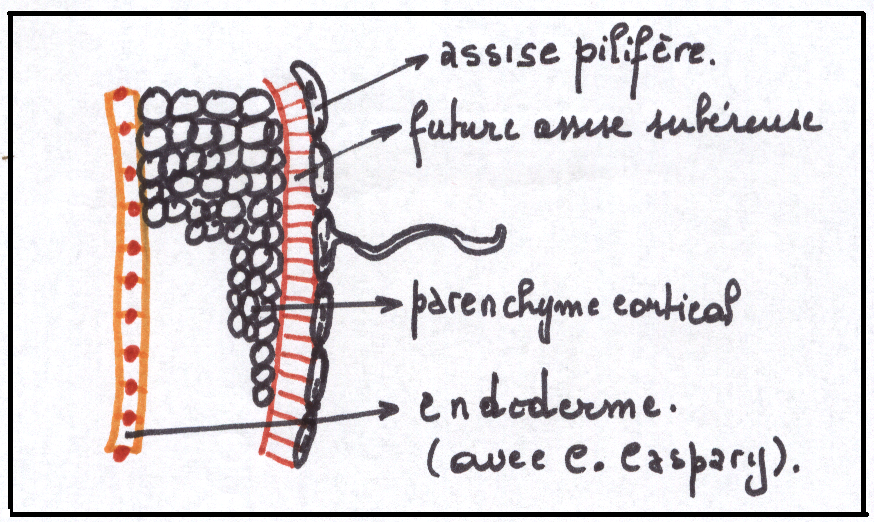
De l’intérieur vers l’extérieur, on distingue successivement :
1) L’assise pilifère , couche externe, méatique , certaines cellules se prolongent en un poil absorbant ( grande vacuole centrale, cytoplasme pariétal,noyau à l’extrémité du poil , membrane pectocellulosique ; perméabilité ! ). Cette assise n’existera plus dans la région subéreuse.
2) La future assise subéreuse comprend une ou plusieurs assises de cellules régulières améatiques, qui après disparition de l’assise pilifère devienfront les plus externes de la racine et joueront un rôle protecteur par subérisation . Ce tissus protecteur
n’est pas fonctionnel aussi longtemps qu’il est sous-jacent à l’assise
pilifère : il le deviendra dans l’assise subéreuse
3) Le parenchyme cortical : c’est une sorte de manchon constitué de plusieurs assises de cellules parenchymateuses , incolores et
méatiques , de plus en plus méatiques à l’intérieur du manchon. Il contient aussi des grains d’amidon à certaines époques de l’année (de plus en plus d’amidon vers le centre).
4) L’endoderme : C’est l’assise la plus interne du cylindre cortical. Elle est composée d’une seule assise de cellules améatiques et très régulières formant une sorte de gaine que l’on peut grossièrement comparer à une cheminée d’usine d’ont l’épaisseur serait limitée à l’épaisseur d’une brique , la brique étant assimilée, en l’occurrence, à une cellule.
Les cellules endodermiques sont surtout caractérisées par un cadre de renforcement, sorte de ruban lignifié ou subérisé selon les espèces, plissé, qui constitue une paroi secondaire tapissant intérieurement les faces radiales et transversales . Ce ruban plissé ou cadre endodermique ou cadre de Caspary apparaît surtout en coupe transversale sous la forme d’un petit tiret d’épaississement au niveau des parois endodermiques radiales
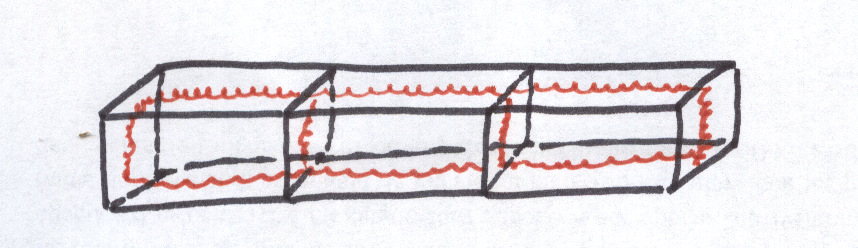
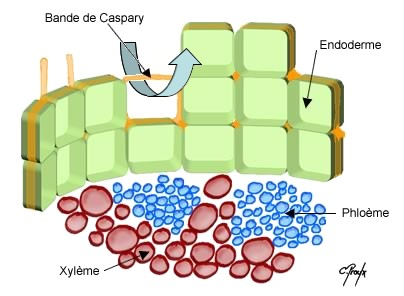
Le cadre de Caspary est un renforcement squelettique qui doit en même temps permettre les perméabilité cellulaire,
c’est pour cette raison qu’il se situe au niveau des faces transversales et radiales et non aux faces tangentielles des cellules.
LE CYLINDRE CENTRAL
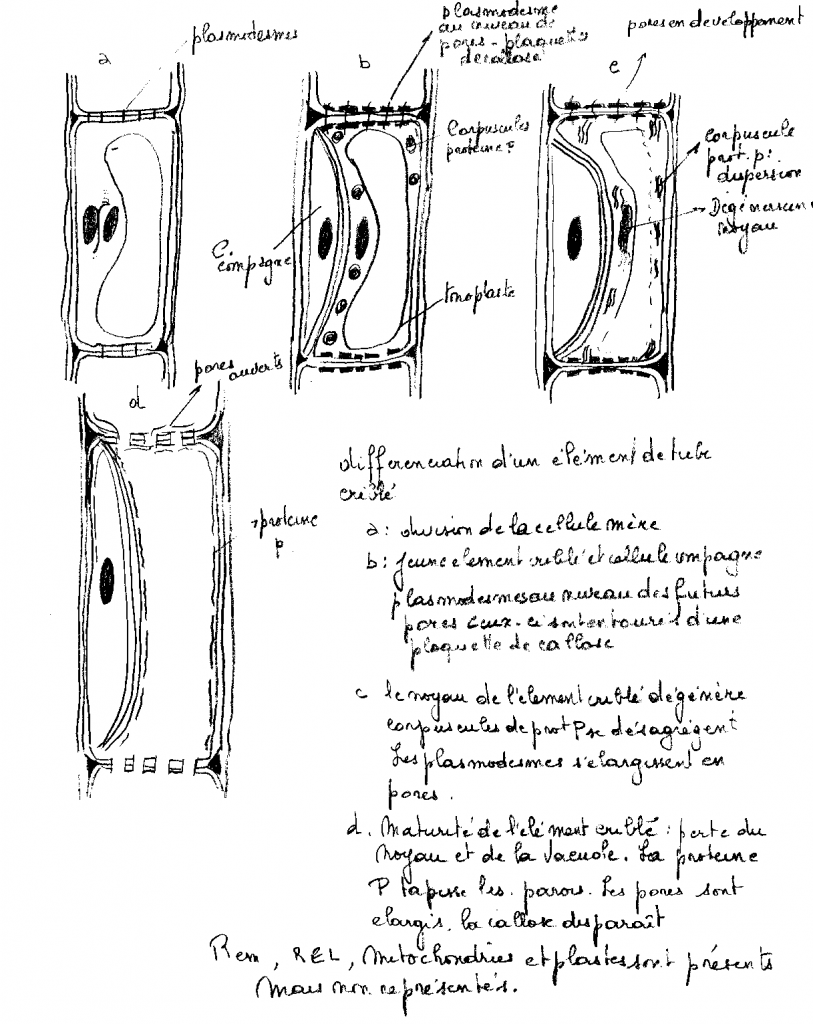
Il occupe l’espace délimité intérieurement par l’endoderme . Le rôle du cylindre central est essentiellement conducteur : sève brute par les lames vasculaires, sève élaborée par les massifs criblés
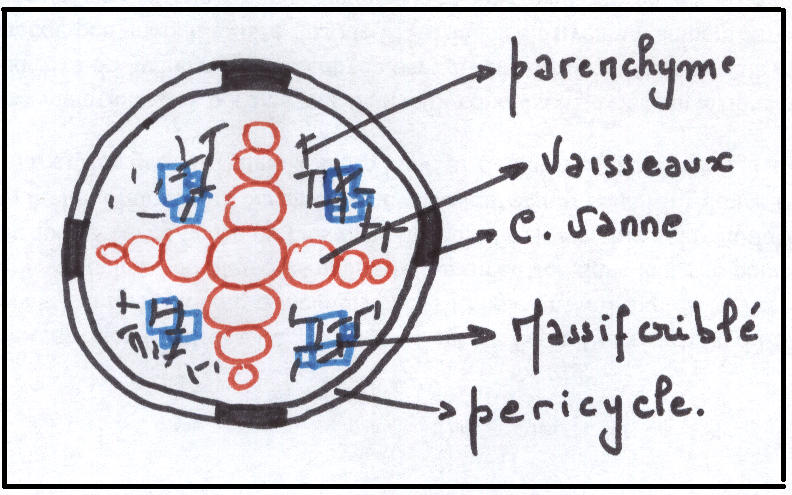
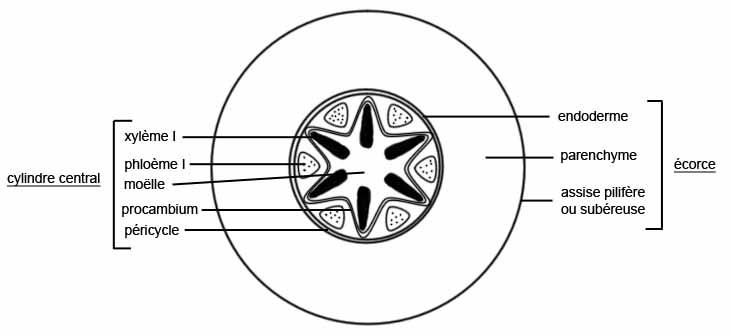
Les lames vasculaires ( LV) sont un ensemble devaisseaux ( trachées) situées plus ou moins radialement et partant du type
annelé ( vers l’endoderme) vers le type ponctué ( au centre ) en passant respectivement par les vaisseaux spiralés, réticulés

Les massifs criblés réunissent un ensemble de tubes criblés avec cellule compagne,groupés de manière que les tube criblés à lumen large sont vers le centre du massif criblé , les tubes criblés à lumen plus ou moins étroit étant vers l’extérieur des massifs criblés.
Ils existe donc une succession de cellules conductrices de manière que les éléments les plus larges ( à parois les plus épaisses) occupent une position plus centrale , et que les éléments les plus étroits ( les moins rigides) soient les plus extérieurs dans le cylindre central, on dit que la différenciation des MC et des LV
est centripède ou exarche.
Les LV et MC alternent régulièrement
Entre les LV et les MC existent des cellules améatiques : c’est un parenchyme très peu différencié (remplissage)
Entre l’endoderme et le point terminal des LV existe une assise régulière de cellules améatiques , à parois minces , appelée péricycle (parfois le péricycle peut être épais de plusieurs cellules)
Le péricycle est également une assise cellulaire parenchymateuse très peu différenciée.
On appelle cellules-vannes les cellules du péricycle qui sont situées dans le prolongement des LV, plus précisément entre le dernier vaisseau annelé et l’endoderme. Ces cellules vannes forment donc autant de rangées verticales qu’il existe de lames
vasculaires dans la racine ; nous verrons plus loin que les cellules vannes sont à l’origine des racines latérales ou radicelles.
On appelle pôle de vascularisation le point de tangence entre une LV et le péricycle (coupe transversale).
Les LV sont composées théoriquement d’une succession de tuyaux situés plus ou moins en ligne droite et partant du plus étroit (vers l’extérieur) au plus large ( vers l’intérieur) ; il se peut que cependant , certains types de vaisseaux manquent , que d’autres se répètent etc… On se limite donc à un schéma général autour duquel gravitent des variantes.
En outre, le nombre de lames vasculaires - et partant de MC qui alternent avec ces dernières – varie d’une espèce végétale à une autre, tout en demeurant assez constant pour une espèce donnée.
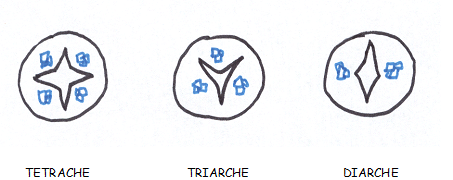
Quelques exemples :
- Racine diarche ; Lupin , Betterave
- Racine triarche ; Ortie
- Racine tétrarche ; Fève, Renoncule
Cours anatomie racine : tpanat1.ppt
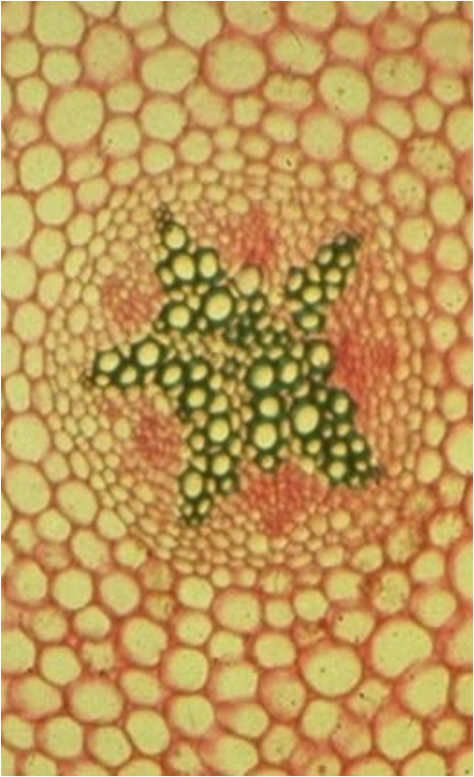
Le nombre de lames vasculaires dépasse rarement 5 chez lesplantes dicotylées
Chez les plantes monocotylées il est au contraire supérieur à 5, parfois très largement : voir plus loin.
RECAPITULATIF DES FONCTIONS ATTRIBUEES AUX DIVERSES PARTIES DE LA RACINE
I. CYLINDRE CORTICAL
- Assise pilifère : poils absorbants = rôle d’absorption radiculaire
- Future assise tubéreuse : deviendra protectrice dans la région subéreuse.
- Parenchyme cortical : méatique, amylifère, épais d’où : remplissage, réserve et manchon amortisseur de la racine
- Cadre endodermique : soutien qui s’oppose à la compression du cylindre central dont la fonction est conductrice ( complémentarité du parenchyme cortical et de l’endoderme).
II. CYLINDRE CENTRAL
- Péricycle : cellules parenchymateuses, améatiques.
Certaines cellules du péricycle sont capables de redevenir méristématiques et de se rediviser : sont génératrices des racines latérales : voir plus loin.
- Les lames vasculaires et massifs criblés : rôle de conduction des deux sèves
- Parenchyme améatique : véritable tissus de remplissage.
VARIANTES, CAS PARTICULIERS PARMI LES SPERMATOPHYTES
1. ABSENCE DE RACINES
Cas de diverses espèces ou familles de plantes vivant sur des substrats particuliers :
- Plantes aquatiques flottantes : Utricularia, Cératophyllum
- Plantes saprophytes vivant sur l’humus ou vivant en épiphytes ( Broméliacées) ou plantes parasites ( Cuscute : convolvulacée).
2. ABSENCE DE COIFFE
Aesculus = Marronnier ( Hippocastanée).
3. ABSENCE D’ASSISE PILIFERE
Surtout le cas des plantes aquatiques qui absorbent de toutes parts ; de plantes à racines aériennes (Orchidées) ; (Monstera deliciosa, Aracée ) : le parenchyme cortical est très développé et entièrement cellulosique.
4. NOMBREUX CAS PARTICULIERS DE RACINES SURTOUT CHEZ LES MEGATHERMES
Racines lianes ; racines échasses ( Pandanacées) ; racines-contrefort ; racines à pneumatophores (plantes de la mangrove) ; racines-épines ; racines-vrilles (Vanille) ; racines-suçoirs ( = haustorias) ; racines de plantes parasites ( Ovobranchacées) ; racines symbiotiques ( mycorhizes ; nodosités bactériennes : fixation d’azote ) ; racines drageonnantes.
5. CAS DES PLANTES MONOCOTYLEES
Les monocotylées présentent deux particularités remarquables :
a) Cadre endodermique en fer à cheval,
la concavité étant tournée vers l’extérieur de la racine ; interruption du cadre au niveau des LV. L’endoderme dit en fer à cheval existe chez la plupart des plantes monocotylées ; on le rencontre toutefois exceptionnellement chez quelques plantes dicotylées.
b) L’élément le plus caractéristique d’une racine de plante monocotylée est l’aspect de son cylindre central.
Alors que chez les dicotylées , les lames vasculaires sont peu nombreuses et se rejoignent au centre , les plantes monocotylées sont pourvues de lames vasculaires très nombreuses , chacune étant alternée avec un massif criblé. Elles ne se rejoignent pas mais sont réunies au milieu du cylindre central par un parenchyme plus ou moins absorbant et dénommé moelle ; elles sont en outre généralement limitées à quelques vaisseaux.
La moelle se transforme souvent en un parenchyme scléreux ou même en sclérenchyme.
Date de dernière mise à jour : 06/03/2018
Ajouter un commentaire