

Les principes de l'évolution biologique
LES MECANISMES DE L'EVOLUTION
I.Microévolution et macroévolution
La modification des caractères héréditaires au sein d'une population est à la base de l'apparition de nouvelles espèces à partir d'espèces ancestrales, c'est l'évolution.
De petites différences accumulées au sein d'une population et soumises aux contraintes évolutives sont la clé de la microévolution. Celle-ci pourra permettre la différenciation des espèces.
Certaines innovations génétique comme des remaniements chromosomiques ou des modifications dans des gènes de développement sont à l'origine de modifications fondamentales au niveau de l'organisme, celles-ci provoqueront l'apparition de nouvelles espèces ou de nouveaux groupes, c'est la macroévolution .
II.La naissance de la théorie de l'évolution .
La théorie de l'évolution est née à partir de concepts élaborés au XVIII siècle et a pris forme suite à un ensemble d'observations. Elle z'oppose à une vision fixiste et créationniste qui suppose quer les espèces se perpétuent identiques à elles-mêmes depuis la création du monde Cette théorie de l'évolution a été formulée par Darwin en 1859, elle s'articule en deux grands principes :
- Les espèces actuelles proviennent d'espèces ancestrales par accumulation de modifications,
La sélection naturelle et l'adaptation sont les moteurs de l'évolution.
- L'adaptation est le dispositif qui permet à un organisme de survivre et de se reproduire dans son environnement
La théorie émise pâr Darwin va être étayée par de nombreuses données scientifiques issues de diverses disciplines comme la paléontologie, la systématique et la génétique. Ces recherches donnent naissance à de nouvelles théories de l'évolution.
Ces découvertes sont sans cesse confrontées à de nouvelles découvertes dans divers domaines et sont régulièrement revues.
Mais si les mécanismes de l'évolution sont encore l'objet de recherches et parfois de controverses, le mécanisme évolutif lui-même est bien une réalité..
III.Les mécanismes de l'évolution
1, La sélection naturelle
1.1 Définition
La sélection naturelle occupe une place capitale dans les mécanismes de l'évolution. En effet, tous les organismes n'ont pas les mêmes capacités pour faire face aux contraintes de l'environnement.Ce sont les plus adaptés qui ont le plus de chances de survivre et donc de transmettre leurs gènes à la descendance, ainsi les génotypes les plus favorables dans un environnement donné sont sélectionnés et deviennent plus fréquents, les autres se raréfient. C'est ce que l'on appelle la sélection naturelle.
- Un exemple typique est celui de la phalène du bouleau .dans les populations de ce papillon coexistent deux phénotypes : des formes claires et des formes sombres , selon les conditions du milieu, pollué ou non, l'une des deux formes est privilégiée car elle a plus de chances d'échapper aux prédateurs et donc de transmettre ses gènes à une descendance.
1.2La sélection naturelle a des effets divers sur la variation du phénotype
- Lors de la sélection directionnelle, les individus qui présentent un phénotype extrême ont un avantage par rapport aux autres individus de la population.

Ce type de sélection est souvent rencontré lorsqu’une population subit des changements extrinsèques, par exemple des changements environnementaux, ou si une partie de cette population émigre dans un nouvel habitat non identique à l’original.
- La sélection diversifiante .
Elimine les types intermédiaires et favorise les phénotypes extrêmes, celle-ci tend à diversifier la population en deux groupes distincts, pouvant mener à terme à la séparation en deux espèces différentes,
Si l'alimentation ( par exemple des graines ) peut se présenter sous des formes graines larges et graines étroites, les becs des oiseaux granivores consommateurs y seront adaptés et l'on trouvera deux groupes d'oiseaux avec très peu d'intermédiaires.
- La sélection stabilisante
elimine les phénotypes extrêmes, ce qui a pour effet d'augmenterla fréquence du phénotype intermédiaire. Ce type de sélection réduit la variation au sein de la population et maintien un statu quo
Dans une population d'une espèce d'oiseaux, il y a la potentialité d'avoir des individus de masse corporelle plus élevée et moins élevée.
La Masse élevée avantagée lors de famines car il y a un plus grand stockage de réserves, les masses faibles sont donc dans ce cas défavorisées, par contre les masses élevées échappent plus difficilement aux prédateurs, ce qui favorise les masses corporelles faibles. Chaque type d'oiseau comporte donc des risques de diminution de l'effectif. Dans ce cas, la sélection naturelle peut favoriser le compromis et favoriser les types intermédiaires, c'est la sélection stabilisante

2.L'évolution des populations.
2.1 Evolution statistique bdu pool génique d'une population
Les mécanismes de l'évolution agissent sur un grand nombre d'individus. C'est au sein d'une population que les modifications apparaissent et qu'elles sont maintenues ou éliminées. La microévolution provoque des modifications au niveau du pool génique d'une population.
- Fréquences alléliques dans une population stable
?Dans une population constituée d'un grand nombre d'individus où les croisements se font de manière aléatoire, les fréquences des allèles et des génotypes sont invariables . Ce phénomène est modélisé par la relation de Hardy-Weinberg. Ainsi pour un locus comportant deux allèles, dans la population, on obtient :
p2 + 2pq + q2 = 1
p = fréquence de l'allèle dominant
q = fréquence de l'allèle récessif
p+q = 1
p2 = fréquence des homozygotes dominants
q2 = fréquence des homozygotes récessifs
pq = fréquences des hétérozygotes
- L'équilibre de Hardy-Weinberg ne se maintient au sein d'une population qu'aux conditions suivantes :
- La population est de très grande taille
- La population est isolée
- Il n'y a pas de mutation nette
- L'accouplement se fait au hasard, de telle manière que les gamètes se mélangent de manière aléatoire.
- Il n'y a pas de sélection naturelle
Ces conditions sont rarement réunies dans une population naturelle. L'équilibre de Hardy-Weinberg décrit la génétique de population théoriques .Néanmoins, il est utile, pour calculer, dans une population donnée,le pourcentage d'individus porteurs ou atteints d'une maladie héréditaire.
3.Les facteurs de la microévolution
Les populations ne sont pas immuables, divers facteurs jouent un rôle dans leur évolution
3.1 Les mutations font apparaître de nouvelles formes alléliques, la sélection naturelle quant à elle augmente la fréquence des formes alléliques adaptées à l'environnement et peu ainsi diminuer la variabilité génique. Des transferts d'allèles par flux génétique entre populations proches peuvent se maintenir et contribuer au maintien de cette variation génétique.
3.2 La diminution du nombre ou la modification de la fréquence des allèles peu se produire en cas de fécondation non aléatoire ou lorsque le nombre d'individus d'une population est restreint, soumettant celle-ci à la dérive génétique .
3.3 Les croisements non aléatoires se rencontrent lorsque dans un groupe les individus se croisent entre eux plutôt qu'avec des individus choisis au hasard dans l'ensemble de la population. Ces types de croisement se rencontrent aussi bien dans le règne animal que le règne végétal. C'est dans ce dernier que l'on rencontre un cas extrême de fécondation non aléatoire : l'auto fécondation. Ce genre de processus engendre une augmentation du nombre d'individus homozygotes.
La fréquence des allèles d'un gène peut être modifiée de façon aléatoire sans que la sélection naturelle n'intervienne. Cela est particulièrement vrai si les divers allèles ont la même valeur adaptative. Des mutations qui ne modifient pas l'activité de la molécule et ne provoquent ni avantage ni désavantage qux individus qui en héritent sont qualifiées de neutres . Celles=ci peuvent se fixer et s'accumuler dans diverses molécules et sont à l'origine du polymorphisme génétique (variété des allèles d'un gène). Ainsi une mutation neutre peut très bien se répandre dans une population même si elle ne présente aucun avantage sélectif., elle peut tout aussi bien être éliminée, c'est aussi une forme de dérive génétique. Ce phnénomène a d'autant plus d'impact que les populations sont restreintes. La diminution des populations peut engendrer la disparition de certains allèles ou au contraire l'augmentation de la fréquence de certains autres .
Ceci peut être du à deux effets :
- L'effet fondateur qui résulte de la colonisation d'un territoire par un petit nombre d'individus.

- L'effet d'étranglement qui résulte d'une brusque réduction de taille de la population ;
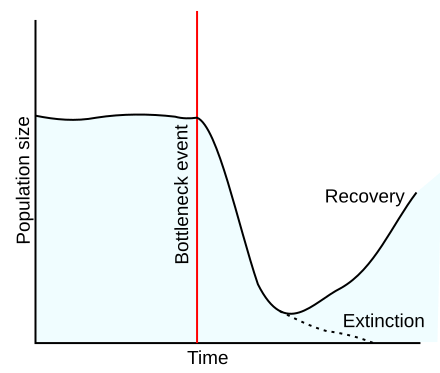
Parmi les différents facteurs qui jouent un rôle dans la modification de la composition allélique, seule la sélection naturelle produit un changement adaptatif, les autres facteurs agissant de façon indépendante de l'environnement.
3.4 Les innovations génétiques
création de familles de gènes
Les mutations ponctuelles ne sont pas les seules modifications spontanées des séquences d'ADN.Il peut en effet se produire des mutations étendues : des séquences plus ou moins longues d'ADN pouvant correspondre à un gène entier, peuvent être perdues, inversées ou bien dupliquées et transposées à un autre endroit du génome sur le même chromosome ou un autre chromosome . La conséquence du processus de duplication/translocation. est donc qu'un organisme hérite d'un deuxième exemplaire d'un gène, situé à un autre locus.. L'évolution ultérieure des deux copies est alors autonome, chaque copie subira des mutations ponctuelles indépendamment de l'autre. Il se formera alors deux gènes distincts codant poue des protéines dont les fonctions être différentes. Il y a ainsi enrichissement du génome avec apparitions de nouvelles caractéristiques phénotypiques.
De tels gènes apparentés présents dans les génomes d'une même espèce constitue ce que l'on appelle une famille (par exemple chez l'homme, les gènes codant pour les différentes chaines de globines).
L'analyse des différences entre les séquences d'ADN d'une famille multigénique permet parfois de retracer l'histoire d'un gène. En effet, on peut admettre que les mutations ponctuelles seront régulièrement se sont régulièrement accumulées dans les copies de gènes après leur duplication . Ainsi , le degré de similitude entre deux gènes d'une même famille traduit le temps écoulé depuis la duplication du gène à l'origine des deux copies.
La duplication est parfois décrite comme une trisomie partielle. Il existe 2 types de duplication :
- Duplication directe (ou en tandem) : le fragment se duplique dans le même sens.
- Duplication inverse (ou en miroir) : le fragment se duplique en sens inverse.
On peut également distinguer les duplications:
Intrachromosomique : le fragment est dupliqué sur le même chromosome à la suite d'un mécanisme de réparation non homologue de l´ADN.
Interchromosomique : le fragment est dupliqué sur un autre chromosome à la suite d'une translocation chromosomique.
Ce phénomène se produit au niveau des cellules germinales n , lors de la fécondation, l'rganisme 2n au niveau du soma et n au niveau de sa lignée germinale conservera le duplication pour certains des gamètes formés.
Les gènes HOX de développement chez les vertébrés sont un exemple de ce genre de fazmille de gènes
Exemple pour comprendre :
- ==========================================================================
Gènes de la globine; drépanocytose I. Généralités
II. Gènes de globine
II.1. Familles de gènes
II.2. Expression séquentielle de gènes au cours du développement
III. Mutations
III.1. Définition
III.2. Exemple d'un gène Platonicien
IV. Clinique - principaux syndromes
IV.1. Drépanocytose
IV.2. β Thalassémies
I. Généralités
Globine :
Protéine entrant dans la composition de l'hémoglobine (Hb).

L'hémoglobine A (forme prépondérante chez l'adulte) a un poids moléculaire de 64400, et est composée d'un tétramère α2β2 et de 4 molécules d'hème, de poids 614 (x 4).
Il existe en effet différents tétramères: chez l'adulte, en majorité, les tétramères sont α2β2, chez le foetus, ils sont plus souvent α2γ2, ils sont encore différents chez l'embryon. En effet, existent toute une série de gènes de globine. Ceci nous amène à 2 notions fondamentales:
Celle de famille de gènes (et la notion corrélée de pseudogène);
Celle d'expression séquentielle de gènes d'une même famille au cours du développement, en particulier embryonnaire (voir aussi le paragraphe sur la famille Hox, dans Gènes du développement; exemple: développement du squelette).
Les anomalies génétiques de la globine peuvent provoquer des anémies hémolytiques: drépanocytose (Hbs), α thalassémies, β thalassémies, que nous verrons plus loin, tableaux plus ou moins sévères selon la mutation et/ou le nombre de mutations (la délétion d'un gène α est latente, celle de 4 gènes α provoque l'hydrops foetalis et la mort in utero ou en période néonatale).
II. Gènes de globine
Les gènes de la globine (et de la myoglobine du muscle) représentent une famille de gènes; l'ancêtre commun a plus de 500 millions d'années. Des duplications du gène ancêtre se sont succédées, et des mutations sur chacun des gènes ont assurés une certaine diversité. Parmi ces gènes dupliqués, beaucoup sont fonctionnels, ils vont entre eux se répartir la tâche à accomplir au sein de l'individu, d'autres ne le sont pas car ils codent pour des protéines non fonctionnelles, ils sont appelés pseudogènes.
Localisation des gènes:

Chaque gène est formé de 3 exons (séquences codantes) séparés par 2 introns (séquences non codantes).
Les gènes de la globine se répartissent la tâche: certains s'expriment chez l'embryon, d'autres prennent la relève chez le foetus, d'autres enfin chez l'adulte. L'expression séquentielle correspond, de plus, à l'emplacement physique des gènes de 5’ vers 3’. Nous verrons aussi qu'un gène “foetal” peut compenser un gène “adulte” défaillant (persistance héréditaire de l'hémoglobine foetale):
II.1. Famille de gènes
Chromosome 11 : localisation en 11p15.5.
Gènes issus d'une duplication ancienne (existence de séquences homologues), ayant dérivés par mutation et recombinaison.
Les gènes Gγ et Aγ codent pour la chaîne γ , avec 1 acide aminé différent en 136. Existence d'un pseudogène ψβ, apparenté aux gènes normaux mais présentant des mutations telles qu'il ne code pour aucune protéine.
Chromosome 16 : localisation en 16p13.3.
Duplication plus récente des gènes α et α qui présentent des séquences nucléotidiques proches et une séquence codante identique. Le gène θ est faiblement exprimé.
II.2. Expression séquentielle de gènes au cours du développement
- Chez l'embryon, tétramères: α2ε2, ζ2ε2, ζ2γ2 et α2γ2 ;
- Chez le foetus: α2γ2 (et α2β2);
- Chez l'adulte, en majorité, tétramère: α2β2 (mais aussi α2δ2 et α2γ2).
3.5 Évolution des génomes par réarrangement chromosomique.
-Fusion de deux chromosomes différents

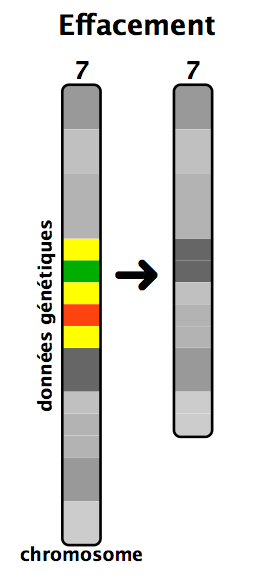
-Suppression ( délétion) d'un fragment
Une délétion peut avoir lieu dans n'importe quel chromosome et peut atteindre n'importe quelle grandeur. Les conséquences d'une délétion dépendent de sa longueur et des gènes qui sont amputés.
Une délétion partielle du bras court (p) du chromosome 5 est responsable de la maladie du Cri du chat
-insertion d'un fragmentde chromosome par fracture en deux points, retournement du fragment et soudure dans cette nouvelle position
 Insertion simple
Insertion simple
Une insertion résulte de l'intégration d'un fragment de chromosome à un autre endroit que son lieu d'origine. Les porteurs d'une insertion peuvent être phénotypiquement sain, car aucune information n'a été perdue.
 Insertion inversion dans un même chromosome
Insertion inversion dans un même chromosome
-translocation qui fait passer un fragment d'un chromosome à un autre ou qui échange des fragments entre deux chromosomes ( translocation réciproque )
![]()
Les anomalies de structure sont le résultat de cassures des chromosomes durant la méiose. Une délétion, une duplication se traduiront par un phénotype anormal, tandis que l'insertion, l'inversion, ainsi que la translocation peuvent être équilibrées. Ceci signifie que les porteurs de ces anomalies de structures sont phénotypiquement sains, car la totalité du matériel génétique est présente.
des chromosomes entiers peuvent aussi se dupliquer ;
Des réarrangements de plus grande ampleur comme la duplication de génomes entiers ou la combinaison avec le génome d'autres espèces sont à l'origine de la polyploïdie . La polyploïdie est un processus clef de l'évolution : plus de ma moitié . Plus de la moitié des plantes actuelles descendent d'espèces formées par polyploïdie . Chez les animaux, ce processus est moins important , mais il a probablement été à l'origine de la formation de certaines espèces d'insectes , de lézards et de poissons.
3.6 Les mutations dans les gènes du développement
- La construction d'un organe exige une succession complexe d'évènements . On estime par exemple que 2500 gènes au moins interviennent pour construire un organe comme l'oeil.
On a cependant découvert qu'un gène unique contrôle toute la cascade d'évènements nécessaires à la construction d'un tel organe . La mutation de ce gène se traduit par une anomalie importante comme l'absence totale de l'organe en question
de tels gènes qui contrôlent le développement des organes à leur juste place sont appelés gènes de développement, une seul gène peut rn effet déterminer le cours du développement d'un individu, le devenir d'un territoire ou la mise en place d'un organe en entier. Une mutation ponctuelle sur un gène de développement peut avoir comme conséquence phénotypique une modification profonde du plan d'organisation
Certains de ces gènes sont des gènes homéotiques, c'est à dire qui codent des protéines qui se lient à l'ADN et contrôlent la transcription d'un grand nombre de gènes. Des gènes homéotiques homologues ont été trouvés chez la plupart des animaux.
Les gènes de développement contiennent une boîte homéotique
================================================================
Une boîte homéotique est composée d'environ 180 paires de bases codant un domaine protéique de 60 acides aminés, l’homéodomaine (ou domaine homéotique) qui est bien conservée (i.e. avec relativement les mêmes acides aminés dans les différents homéodomaines). Cet homéodomaine est capable de se lier à l'ADN pour contrôler l’expression des gènes. Ainsi, l'homéodomaine permet la reconnaissance d'une séquence spécifique d’ADN d’un gène. Les protéines contenant ces homéodomaines, les homéoprotéines, agissent comme facteurs de transcription régulant l'expression de ces gènes cibles durant le développement embryonnaire, la différenciation cellulaire et la spécification du destin de la cellule.
L’homéodomaine est une structure tridimensionnelle appelée hélice-tour-hélice (HTH) et comporte trois hélices alpha séparées par une boucle et un tour (voir Figure 2). La troisième hélice alpha va venir s'insérer dans le sillon majeur de la double hélice d'ADN au moment de la reconnaissance et va rendre cette dernière active pour la transcription.
Les homéoprotéines en tant que facteurs de transcriptions remplissent une grande variété de fonctions biologiques. Il n’existe probablement pas de tissus dans les plantes ou les animaux qui ne requièrent pas ces protéines pour fonctionner correctement. Chez les animaux, elles agissent dès les premiers stades de développement et elles sont essentielles dans les cellules souches embryonnaires4. Plusieurs sont impliquées dans le développement du système nerveux et la perturbation des gènes homéotiques peut mener à divers troubles et maladies génétiques. Également chez les plantes, les gènes homéotiques régulent de nombreux aspects du développement comme la maintenance des cellules souches, la réponse au stress et la réponse à la lumière.
=================================================================
Des mutations dans certains gènes du développement peuvent changer la chronologie du développement. On appelle hétérochronie la modification de la durée et de la vitesse du développement de l'organisme au cours de l'évolution
Chez l'homme, l'augmentation de la durée du développement embryonnaire et de la phase juvénile est à l'origine des traits humain caractéristiques , comme le développement du cerveau et de la bipédie.
Des différences phénotypiques importantes entre des espèces génétiquement très proches sont régulièrement observées. L'évolution n'est donc pas uniquement due aux petites modifications progressives du génome au cours du temps. Des modifications génétiques limitées au point de vue des gènes concernés ont pu se traduire par des modifications de grande ampleur responsables de l'apparition des caractères fondamentaux d'une espèce.
4. Apparition de nouvelles espèces ou de groupes entiers d'organismes : la macroévolution. Voir théories de l'évolution de la partie enseignement supérieur de ce site
?4.1 Microévolution et macroévolution
la plupart des mutations génétiques n'engendrent que de faibles différences , elles contribuent à élargir le pool généique au sein de la population . La sélection naturelle, la dérive génétique, les flux génétiques et les croisements non aléatoires modifient ce pool génétique, ces agents seront responsables de la . Microévolution . Par la suite l'isolement des populations et l'accumulation des changements génétiques pourront aboutir à la différenciation des espèces.
Certaines mutations génétiques dans les gènes de développement ou des remaniement chromosomiques importants modifieront profondément le phénotype et seront à l'origine de la formation de nouvelles espèces voire de nouveaux taxons., c'est la macroévolution . Certaines macroévolutions peuvent aussi être la conséquence de microévolutions s'étendant sur de longues périodes.
4.2 L'apparition de nouvelles espèces
L'espèce est définie comme l'ensembl des individus capables de se reproduire entre eux pour donner une descendance féconde . Cette définition biologique de l'espèce basée sur l'interfécondité se heurte à de nombreuses limitations , elle ne peut s'appliquer aux espèces fossiles ni aux organismes qui ont majoritairement une reproduction asexuée ( telles que les procaryotes ou certaines plantes ). Dans ce cas, l'identification des espèces s'effectue sur base des caractéristiques morphologiques ou génétiques.
-La spéciation ou processus de formation de nouvelles esp-ces nécessite l'isolement des populations. En effet, sans isolement , le flux génétique tend à maintenir les populations homogènes en réduisant les différences dues à la sélection naturelle ou à la dérive génétique.
-L'isolement peut être provoqué par l'apparition d'une barrière géographique , c'est la spéciation allopatrique . D'aqutres types d'isolement sont néanmoins possibres et engendrent la spéciation sympatrique . ce type de spéciation se rencontre lorsqu'un ou plusieurs individus sont isolés ,sur le plan de la reproduction, des autres individus de leur espèce . Diverses causes différentes peuvent provoquer le sélection sympatrique comme la polyploïdisation ou la sélection diversifiante.
La spéciation est un processus généralement lent qui nécessite un grand nombre de générations et dure des milliers voire des millions d'années . Cependant lors des changements phénotypiques suite à des polyploïdisations, ou a des mutations dans les gènes homéotiques , une nouvelle espèce peut apparaître en unbe seule génération.
Date de dernière mise à jour : 01/06/2018
Ajouter un commentaire


