
Ecologie explicative 1
ECOLOGIE EXPLICATIVE
→L'écologiste doit tenir compte d'un certain nombre de facteurs qui influencent la composition floristique d'une aire géographique donnée ; on peut grouper ces facteurs comme suit :
A) Facteurs ou causes historiques relatifs au peuplement végétal.
B) Facteurs géographiques
C) Facteurs de concurrence ou d'aide mutuelle
D) Facteurs du milieu
E) facteurs humains; interventions animales ( ex : rongeurs, gibier )
I) De son côté, la plante réagit aux facteurs du milieu par sa morphologie, sa physiologie et par la composition chimique de ses tissus. Et on constate que les plantes qui appartiennent à un même groupement végétal présentent des caractères communs à cet égard Phyto-géo-chimique.
2) Il faut encore rappeler que les plantes restituent au milieu, après leur mort - ou encore périodiquement : ex de la chute des feuilles - les substances qu'elles ont élaborées. Cete restitution intervient dans un cycle puisque les matières libérées seront réutilisées, recyclées par d'autres plantes.
Conclusion : le groupement végétal est donc le rouage d'un écosystème complexe auquel participent également des animaux et des microorganismes.
A. LES FACTEURS HISTORIQUES ET GEOGRAPHIQUES
On peut les illustrer par divers exemples.
Exemple 1 : quaternaire, glaciations. Mise en parallèle des flores de l'Amérique du Nord et de l'Europe au point de vue forestier .
La disposition N-S des montagnes en Amérique a permis une migration vers le sud des plantes thermophiles, puis une recolonisation vers le Nord après réchauffement du climat.
En Europe : allongement des montagnes d'Est en Ouest et obstacle de la Méditerranée. Par conséquent, les espèces plus délicates ont disparu de la flore du continent européen, elles n'ont pas pu migrer.
Exemple 2 : Taillis de Chênes et de Bouleaux de l'Ardenne, avec une flore herbacée très pauvre sont le résultat d'une mise en culture après incinération ( = essartages ). Les mêmes sols, dans la même région portent des peuplements de Hêtres s'ils n'ont pas été exploités de manière brutale.
Exemple 3 : certaines espèces ont une aire de dispersion immense en raison de la grande faculté de dispersion de leurs graines ( Ex. Pins, Epicea); D'autres espèces ont une aire restreinte et morcelée ( ex : Sapin pectiné ) parce que leurs grines sont lourdes, à pouvoir germinatif médiocre etc...
Exemple 4 : Certaines vallées alpines peuvent différer entre elles par leurb flore parce que séparées par des massifs élevés , inaccessibles aux semences d'autres plantes.
Ces divers exemples font ressortir qu'à côté d'une aire naturelle existe une aire potentielle des espèces, pourvu qu'elles soient aidées ( par l'homme par exemple ).
La composition de la flore n'est pas immuable dans un territoire donné ; des espèces disparaissent, d'autres viennent d'un autre territoire, voyons des exemples importants dans ce qui suit :
D'après Ascherson (Paul Friedrich August Ascherson ( - ) est un botaniste allemand.), on peut écrire ce qui suit :
Chaque espèce végétale occupe sur le globe une aire naturelle déterminée plus ou moins grande, mais qui ne s'étend jamais à toute la surface de la Terre. Il est même rare qu'une plante soit répandue sur plus d'un tiers de la terre ferme, Ce cas se rencontre en zone tropicale; environ trois ou quatre cents de ces plantes occupent toute l'étendue des tropiques ou même les dépassent .
exemples : les Lycopodium cernuum, Pistia stratiotes, Eleusine indica, Argemone mexicana, Cleone (Gynandropsis) pentaphylla, Waltheria americana, Ricinus communis, Hydrocotyle asiatica, Cassia occidentalis, Mimosa asperata, Herpestes Monniera, Ageratum conyoides, dont quelques-unes sont sans doute arrivées par émigration de l'ancien dans le nouveau continent. Il s'agit là de l'extension topographique de plantes dans un climat déterminé.
Une même plante se rencontre rarement dans deux climats différents ; nous pouvons cependant citer comme appartenant à la fois aux régions chaudes et tempérées les Pteris aquilina, Osmunda regalis, Juncus tenuis, Potamogeton pectinatus, Ruppia maritima, Zannichellia palustris, Panicum crus galli, Dactylus officinalis, Eragrostis minor, Stellaria media, Portulaca oleracea, Oxalis corniculata, Solanum nigrum, Gnaphalium luteo-album.
Majoritairement,les plantes occupe des aires bien plus restreintes. Très faible est même le nombre des espèces qui s'étendent sur toute l'Europe, depuis les provinces arctiques de la Russie jusqu'à l'extrémité des péninsules méditerranéennes;
exemples :
- Capsella bursa pastoris (présente à la Terre de Feu) et Thymus serpyllum.
- Circaea alpina, très répandu dans l'Amérique du Nord et en Asie, mais manque dans le Nord de la Russie et dans les montagnes de l'Espagne et de la Grèce soit un bel exemple de ces aires géographiques allongées dans le sens Est-Ouest, qui sont beaucoup plus fréquentes que les aires allongées dans le sens Nord-Sud. Ces aires sont surtout fréquentes dans l'hémisphère boréal et sur l'ancien continent.
- la région forestière qui s'étend depuis la France jusqu'au Kamtchatka est partout caractérisée par le Pinus sylvestris;
- puis une zone s'étendant dans la région méditerranéenne depuis le Portugal jusqu'en Anatolie et en Syrie et caractérisée par l'Anagyris foetida, le Vitex agnus castus et le Nerium oleander;
- plus au Sud, dans la région saharienne, une aire qui s'étend depuis les îles Canaries et du Cap-Vert jusqu'à l'Indus inférieur avec des plantes telles que le Scrophularia arguta et l'Anticharis linearis.
En Amérique, au contraire, on remarque dans les régions tropicales et suivant les Andes ou le littoral maritime oriental ou occidental des aires de distribution allongées dans le sens du méridien ( nous avons expliqué plus haut, les raisons de ce phénomène )
Dans les régions sèches du globe, les aires sont généralement petites, mais les espèces sont nombreuses dans une même aire. Des plantes se trouvent même parfois à un point unique du globe; ainsi le Statice arborescens ne se rencontre à l'état spontané que sur une seule petite île rocheuse de la côte de Tenerife. Dans les déserts mêmes, la végétation ne peut subsister; cependant on trouve en plein désert, sur le djebel Ahmar (montagne rouge), près du Caire, le Fagonia latifolia, plante très caractéristique. Dans l'Europe moyenne et en particulier dans les Alpes se rencontrent également un certain nombre d'aires très circonscrites pour des espèces absolument caractéristiques; tels sont entre autres le Wulfenia carinthiaca, limité au seul Gailthal, en Carinthie;le Sangirisorba dodecandra, exclusivement propre à la Valteline, etc.
Chaque espèce végétale a un berceau (lieu d'origine) unique ne correspondant pas nécessairement à l'aire actuelle de l'espèce - se rappeler que, lors de la dernière ère glaciaire, nombre d'espèces ont été refoulées vers le Sud, loin de leur berceau -
D'autre part, pour concevoir la formation des aires actuelles, on est obligé d'admettre que les individus ont rayonné de plusieurs points, sinon d'un seul, ce qui paraît le plus vraisemblable; ce point appelé centre de végétation par Grisebach et plus communément centre de dissémination ou de dispersion, se confond avec le centre d'origine lorsqu'il est démontré que les espèces se sont maintenues dans leur aire primitive. Il n'a rien de commun avec le centre géométrique de l'aire et ne peut être déterminé que par des considérations de géologie historique; encore ne peut-on que rarement arriver à une certitude à cet égard.
Chaque espèce a rayonné de son point de départ, aussi loin que le lui ont permis les conditions météorologiques et climatiques, abstraction faite de la compétition avec d'autres espèces qui peuvent en avoir restreint l'aire, ou jusqu'à ce qu'elle ait rencontré un terrain qui ne lui convenait pas, ou un obstacle matériel insurmontable tel que la mer, un désert ou une chaîne de montagnes couverte de glaciers; une plante de plaine est aussi impuissante à franchir une chaîne de cette importance, qu'une plante d'altitude à franchir la plaine. Les limites imposées à l'extension d'une espèce par les conditions climatiques, sont appelées, depuis Grisebach, lignes ou limites de végétation et se reconnaissent à leur forme généralement régulière coïncidant plus ou moins avec certaines lignes climatiques et à ce que la culture de cette espèce au delà de ces lignes ne réussit plus, ce qui n'est pas le cas pour les obstacles mécaniques et les barrières naturelles. Les lignes de végétation sont le plus faciles à constater pour les plantes dites sociales et surtout pour les plantes cultivées, car la culture se fait habituellement jusqu'aux plus extrêmes limites; c'est le cas des arbres de nos forêts, des céréales, etc.
Les plantes ne se comportent pas toutes de la même manière sur leur limite de végétation. Souvent la transition est brusque; c'est ce qui arrive pour l'Euphorbia cyparissias si répandu en France et qui atteint sa limite en Allemagne; encore très commun à Neu-Strelitz, il n'existe plus à Neu-Brandenbnrg, un peu plus au nord. D'autres espèces, plus rares ou plus disséminées, se présentent comme des postes avancés; leurs divers habitats sur une carte formeraient une sorte d'archipel dont les îles, très rapprochées vers le centre de l'aire se trouveraient très éloignées les unes des autres près de sa circonférence. Il en est ainsi par exemple, de l'Inula conyza; étendu sur un grand espace dans l'Europe moyenne et dans l'Asie antérieure, sans jamais être commun, il n'occupe que quelques postes avancés près de Krossen, sur l'île de Rügen, et sur les îles danoises, et manque totalement dans les plaines baltiques.
D'autres aires, en grand nombre, se composent de groupes d'îlots ou d'îles séparés les uns des autres par de grandes distances. C'est le cas de l'Aldrovanda vesiculosa qui se rencontre en Prusse, puis près du lac de Constance, dans l'ouest et dans le midi de la France, dans la Haute-Italie, en Hongrie, en Lituanie, en Inde (Calcutta), en Australie (Rockhampton) et dans l'Afrique centrale (Bahr el-Ghazal); cette énumération est certainement incomplète, mais il n'en est pas moins vrai que les stations de l'espèce sont très disséminées. Du reste le même fait s'observe même pour des plantes dont l'aspect est beaucoup plus frappant et que leur taille ou d'autres particularités ne permettent pas de ne pas apercevoir, pour certains arbres par exemple; ainsi le Cedrus Libani qui existe dans le Sud de l'Anatolie, au Liban et à Chypre, se retrouve sous une forme légèrement différente (C. atlantica) dans les montagnes du Nord-Ouest de l'Afrique; il en est de même du Rhododendron ponticum des montagnes de l'Anatolie et de la Syrie qui reparaît dans le Sud-Ouest de la péninsule ibérique sous une forme un peu différente (R. baeticum).
Les aires d'une même espèce peuvent être très distantes les unes des autres; c'est ce qui arrive par exemple pour les plantes de montagne qui se retrouvent en plaine à une latitude plus élevée; ainsi l'Empetrum nigrum, qui manque dans les tourbières de la plaine, dans l'Europe moyenne, se trouve dans la plaine et les forêts sablonneuses du Nord de l'Allemagne, puis sur les montagnes relativement peu élevées telles que la Forêt-Noire et dans les Alpes. Citons encore le Pedicularis sudetica des plaines de la région arctique en Europe et en Asie qui fait également partie de la flore alpine, le Polygonum bistorta qui est à la fois circumpolaire, alpin et répandu dans presque toutes les régions tempérées de l'hémisphère boréal, puis le Mélèze (Larix europaea) qui existe dans la région arctique sous la forme très voisine du Larix Sibirica, et le Pinus cambra qui est identique dans les Alpes et dans les plaines arctiques forestières. Citons enfin deux espèces très intéressantes sous le même rapport, le Myricaria germanica et l'Hippophae rhamnoides qui descendent des Alpes en suivant les cours d'eau, puis reparaissent, le dernier sur les dunes du Nord-Ouest de la France, les deux au Tibet, dans le Koukou-Nor, à une altitude de 3600 m, où ils acquièrent des dimensions que nous ne leur voyons jamais en Europe; l'Hippophae atteint jusqu'à 6 m.
L'extension d'une même espèce d'une aire à une autre fort éloignée ne s'est certainement pas toujours effectuée pendant la période géologique actuelle; mais des transports éloignés sont possibles par l'intervention des humains, intervention qui est le plus souvent involontaire. Ce fait ressort surtout bien nettement pour le Xanthium spinosum; originaire de la Russie méridionale, cette espèce, essentiellement xérophile, s'est répandue dans l'Europe moyenne et méridionale, où elle est relativement disséminée, et a fini par pulluler d'une manière extraordinaire dans les pâturages secs de l'Amérique méridionale et de l'Australie, au point d'incommoder les troupeaux par ses fruits adhérents par masses énormes à leurs poils.
Dans une même zone climatique on trouve souvent, à de grandes distances et sur des continents différents, des représentants d'un même genre ou d'une même famille, distincts, mais en quelque sorte équivalents et similaires. Tels sont par exemple les Magnolia, les Illicium de la Chine et du Japon et ceux de l'Amérique du Nord, les Chênes, les Hêtres, les Conifères d'Europe et d'Asie et ceux de l'Amérique du Nord, les Cunnaera, les Libocedrus, les Hêtres de l'île Auckland et ceux de la Terre de Feu; de même comme représentant des familles, nous trouvons en Australie les Epacridées remplaçant les Ericées des autres continents; les Lobéliacées de l'Amérique correspondant aux Campanulacées de l'ancien continent. Les Asclepias d'Amérique sont remplacés en Europe par les Cynanchum ou les Vincetoxicum, etc. Enfin d'un pôle à l'autre il y a même des formes équivalentes comme l'Empetrum rubrum à fruits rouges de la Terre de Feu qui correspond à notre Empetrum nigrum circumpolaire; de même les Acaena y représentent nos Sanguisorba, les Colobanthus nos Spergula.
Ces faits nous font déjà pressentir l'importance du rôle que jouent dans la répartition des végétaux à la surface du globe les aires des groupes naturels, sous-genres, genres, tribus, familles. Aussi bien que les espèces, ces groupes ont des aires plus ou moins nettement circonscrites, mais d'une étendue beaucoup plus considérable. L'étude de ces aires a pour le botaniste une importance bien plus grande que celle des aires des espèces; car elle peut, aidée des observations paléontologiques, jeter quelque jour sur la marche suivie par les Végétaux dans leur distribution et leurs migrations et même sur leurs transformations (évolution). On peut dire que quelques familles ont pour aire le globe entier; ce sont surtout les Graminées et les Composées, représentées depuis les pôles jusqu'à l'équateur; immédiatement après viennent les Légumineuses qui s'avancent moins que les précédentes dans les régions froides.
- Parmi les familles sensiblement cosmopolites, dont les représentants sont moins nombreux dans les contrées tempérées que dans les régions torrides, se trouvent encore et surtout :les Aroïdées, Orchidées, Laurinées, Urticinées, Euphorbiacées, Malvacées, Ménispermées, Ampélidées, Rotacées, Convolvulacées, Bignoniacées, Apocynées, Solanées, Rubiacées, etc. Les Palmiers, Zingibéracées, Musacées, Pipéracées, Mélastomacées sont limitées à peu près exclusivement aux régions les plus chaudes.
- Sont au contraire bien moins répandues dans les contrées tropicales que dans les zones tempérées et froides : les Polygonées, Ericacées, Gentianées, Violariées. Les Pinus et Abies, les Bétulinées, Salicinées, Cupulifères, Renonculacées, Crucifères, Papavéracées, Caroyphyllées, Ombellifères, Rosacées, Ribésiacées, Sambucinées, Acérinées, etc., occupent à peu près tout l'hémisphère boréal extratropical; elles ont néanmoins des représentants plus ou moins insolites, soit dans les régions australes extrêmes, soit sur les hautes montagnes des pays tropicaux.
- L'Amérique seule possède les Loasées, Cactées (un seul Rhipsalis est africain), Broméliacées, Sarracéniacées, Margraviacées, Vochysiacées, Hydrophyllacées, Gessnériacées, Tropaeolées, ainsi que les genres Agave et Yucca, etc.
- Les Tamariscinées, Frankéniacées, Diptérocarpées, Pittosporées, Cyrtandracées, Népenthacées, Casuarinées n'habitent que l'ancien monde;
- les Pénéacées, Bruniacées, Restiacées, Sélaginacées, Mésembryanthémées, avec la plupart des Pelargonium, sont confinées à l'Afrique méridionale. Les Protéacées, les Conifères des genres Araucaria, Podocarpus, Libocedrus ont pour aires actuelles les régions subtropicales et tempérées de l'hémisphère austral.
- L'Australie est le berceau à peu près exclusif des Trémandracées, Epacridées, Myoporinées, Stylidiacées, Goodéniacées, ainsi que des acacias à phylIodes, des Kennedya, Chorizema, Eutaxia et quelques autres Légumineuses, qui, avec, les Eucalyptus, Metrosideros, Melaleuca parmi les Myrtacées, impriment un caractère si spécial à ce continent.
Enfin beaucoup de familles ont des genres on des sous-genres confinés à un continent ou à une région encore plus circonscrite;
- tels sont les Nicotiana, les Petunia parmi les Solanées, tandis que les Jusquiames appartiennent à l'ancien continent.
- L'Adansonia (Baobab) est africain ou madécasse.
- De même, dans le grand genre Euphorbe, les espèces charnues (sous-genres Euphorbium et Diacanthos) sont exclusivement propres à l'Afrique entière, à Madagascar et à l'Inde méridionale,
- Le groupe de nos Euphorbes de la section Tithymalus, lui, a une aire qui s'étend en latitude de l'Europe à l'extrême Asie d'une part, à l'Afrique d'autre part.
- En Amérique, au contraire, nous trouvons les groupes contrastants, Poinsettia, Tricherostigma, etc., qui s'étendent dans les régions tropicales et tempérées. Il serait aisé de trouver des exemples analogues dans d'autres familles, comme celui des Stapelia (Asclépiadées) qui sont exclusifs à l'ancien continent. (Dr L. Hahn et A. Jobin).
On appelle espèce naturalisée :
Une espèce introduite dans la végétation naturelle et dont le comportement est identique à celui d'une plante indigène.
Ex : Lagoseris sancta : composée du bassin méditerrannéen. En 50 ans, cette plante a colonisé la France jusqu'au Nord Est et est une adventice sur sols calcarifères, notamment dans les vignes.
Ex : Elodea canadensis : Plante aquatique venant du Canada vers 1850, elle se rencontre maintenant dans tous les milieux aquatiques à eaux riches en substances dissoutes.
On appelle espèce subspontanée :
Une espèce cultivée, échappée des jardins. On rencontre souvent ces plates subspontanées dans des groupements "ouverts" ex. bords de routes ou des associations relativement simples .
Ex : Asters américains → terrains vagues en périphérie des villes; Asters divers → bords de routes, talus, certaines plantes → voisinage des cimetières, chemins de fer
On appelle espèce adventice:
Une espèce d'origine différente, extérieure qui apparaît dans des associations végétales dont la compositions est influencée par les activités humaines ;
ex des "mauvaises herbes " en agriculture ", des plantes colonisant les ballasts de chemin de fer, les potagers comme Mercuralis annua, Fumaria off. sur sols riches en azote et fréquemment travaillés.
B) LES FACTEURS DE CONCURRENCE ET D'AIDE MUTUELLE
I. Amplitude physiologique et amplitude écologique
L'amplitude physiologique est relativement aisée à établir, par ex dans des cultures en vases de végétation ( Conditions optimales, facteurs limite de vie,...), mais on constate que les conditions expérimentales aboutissent souvent à des conditions qui ne se vérifient pas dans la nature à cause de la concurrence entre espèces.
ex : Jacinthe sauvage ( hyacinthoïdes non scripta ) très héliophile et vigoureuse en culture. En Brabant, elle ne quitte pas les bois et les haies, et vit en stations tamisées.
Certains Asters maritimes ( A.tripolium ) ont une vitalité maximum en sol cultivé sur eau douce ; la croissance diminue progressivement si on ajoute NaCl au milieu, elle s'annule pour une concentration égale à 75 % de celle de l'eau de mer. Or cette plante vit spontanément dans des pré salés ! et dans des groupements végétaux fréquemment envahis par l'eau de mer !! ??.
L'amplitude écologique est différente de l'amplitude physiologique ou potentielle. Cette amplitude écologique correspond aux valeurs limites du milieu entre lesquelles la plante peut vivre : c'est le comportement de la plante qui vit entourée de tous les facteurs qui l'encadrent, en particulier les facteursde concurrence.
Une espèce peut être empéchée d'occuper toutes les stations qui lui conviennent dans le contexte de leur amplitude physiologique à cause de la concurrence entre espèces, cette notion permet de comprendre ce qui différencie l'amplitude physiologique de l'amplitude écologique .
L'amplitude écologique d'une espèce se mesure par sa capacité à se reproduire dans des conditions écologiques différentes. Les espèces occupant des zones très étroites sur le gradient sont des espèces spécialistes, caractérisées par une petite amplitude écologique pour le gradient considéré. D'autres, par contre, occupent de larges zones car elles ont une amplitude écologique beaucoup plus large et sont beaucoup moins exigeantes.
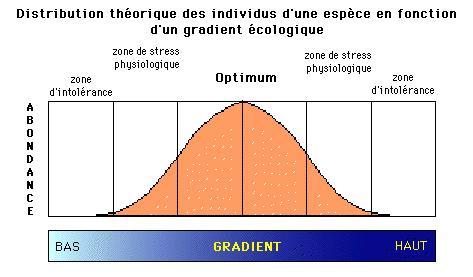
Généralement, la distribution d'une espèce sur un gradient est représentée par une courbe en forme de cloche (ou courbe de Gauss), symétrique, qui indique qu'au fur et à mesure que les conditions écologiques deviennent favorables, l'abondance de l'espèce augmente d'abord jusqu'à un maximum - l'optimum - puis diminue lorsque les conditions écologiques redeviennent défavorables. L'aplatissement de cette courbe est une mesure de l'amplitude ou de la tolérance de l'espèce. Plus elle se sera grande, plus la courbe sera aplatie. Les espèces typiques des extrêmes du gradient n'expriment en fait que la moitié droite ou la moitié gauche de la courbe en cloche si elles montrent leur optimum, respectivement, pour la partie gauche du gradient ou la partie droite.
Les relations existant entre plantes croissant bsur une même parcelle peuvent être directes ou indirectes.
II. Les relations directes entre plantes
- Lianes et épiphytes
- Parasitisme
- Symbiose
Lianes et épiphytes
Les lianes se développent sur une plante hôte qui sert de support et permet aux organes assimilateurs et aux inflorescences de ces lianes de se développer dans une strate plus lumineuse. Il se peut cependant que, parfois, la plante-support soit étouffée par la liane ( ex : Figuier "étrangleur" ), ou encore le liseron sur jeunes peupliers !
Les épiphytes, abordons les par les remarques suivantes :
- Mousses, lichens sur troncs dans nos forêts, surtout si l'humidité est relativement élevée par rapport à la moyenne.
- Pafois, l'humus produit par ces épiphytes peut s'accumuler et servir de substrat à des plantes vasculaires ( ex : la fougère polypodium vulgare ).
- En régions tropicales et équatoriales, le développement d'épiphytes peut être exhubérant : Fougères, Orchidées, Broméliacées .
Parasitisme
Généralement, les végétaux sont caractérisés par leur autotrophie, c'est-à-dire la capacité de synthétiser les molécules organiques à partir d'éléments simples (CO2, H2O, lumière). Ces synthèses se font grâce à la photosynthèse qui a lieu dans les organes chlorophylliens.
Cependant, au cours de leur évolution, certaines plantes Phanérogames (environ 4000 espèces) ont perdu leur autotrophie et sont devenues des parasites d'autres plantes supérieures désormais appelées plantes hôtes. Elles y puisent les nutriments dont elles ont besoin.
Le parasitisme a entraîné le développement d'un organe particulier, le suçoir ou haustorium qui représente un pont structural et physiologique permettant le transit des substances nutritives de l'hôte vers le parasite. Le suçoir assure également la fixation du parasite sur l'hôte.
Selon leur niveau d'hétérotrophie, on distingue les plantes hémiparasites chlorophylliennes, partiellement parasites car capables de réaliser la photosynthèse et les plantes holoparasites totalement dépourvues de pigments photosynthétiques et donc tributaires de leur hôte pour les substances carbonées, l'eau et les sels minéraux.
Selon leur niveau de fixation sur l'hôte, on distingue les plantes parasites épiphytes implantées sur les parties aériennes de leurs hôtes et les plantes parasites épirhizes, fixées sur le système racinaire des plantes hôtes.
Les plantes parasites se rencontrent seulement chez les angiospermes (monocotylédones et dicotylédones). Elles présentent généralement une régression plus ou moins marquée de l'appareil végétatif. Elles peuvent être selon le cas des parasites spécifiques de certaines espèces ou bien plus généralistes.
Toutes les plantes sans chlorophylle ne sont pas nécessairement parasites, il peut s'agir aussi de plantes saprophytes, qui puisent leur alimentation dans la matière organique en décomposition.
Exemples de plantes parasites : la cuscute, les deux espèces de monotrope, les diverses orobanches
Exemples de plantes hémiparasites : le gui. le santal
Symbiose
Historiquement, le concept de symbiose est né de l’observation des lichens. 25 % des espèces connues de champignons (Ascomycètes et Basidiomycètes) sont impliquées dans des lichens. L’algue (verte, filamenteuse ou unicellulaire) peut vivre seule mais, en général, pas le champignon. Les hyphes du champignon pénètrent dans l’algue.
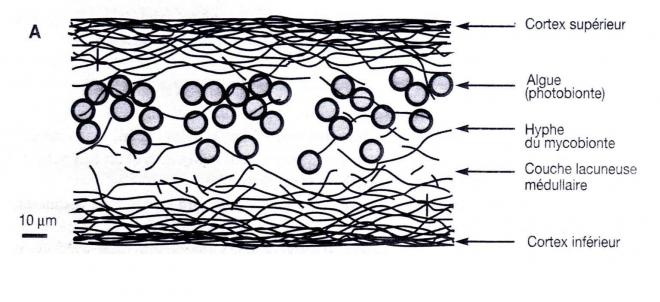
Sélosse M.A.2000. La symbiose, structures et fonctions, rôle écologique et évolutif. Paris, Vuibert
90 % du carbone de l’algue est cédé au champignon (polyols: C4 à C6). Le champignon attaque le substrat avec des acides organiques. Les substances lichéniques (terpènes, molécules aromatiques) ne sont synthétisées par le champignon qu’en présence de l’algue. Elles cristallisent dans les espaces intercellulaires (40 % du poids sec du lichen). Elles protègent contre les herbivores et neutralisent la lumière qui endommage l’appareil photosynthétique.
Le mutualisme est une interaction facultative positive entre deux organismes, Ex: les fleurs et les insectes pollinisateurs
La symbiose est un cas particulier de mutualisme: les organismes « vivent ensemble », en une association réciproquement bénéfique. La symbiose accroît la valeur sélective (contribution à la génération suivante) des deux organismes. Au cours de l’évolution, on peut passer facilement du mutualisme à la symbiose.
Entre 95 et 100 % des végétaux vivent de façons symbiotiques. Le plus souvent, on distingue l’hôte c’est-à-dire l’individu receveur (souvent bénéficiaire) dans le cas des mutualistes, et le donneur pour les deux autres cas (commensalisme et parasitaires), l’individu symbionte sera le principal bénéficiaire dans le parasitisme et le commensalisme.
Dimension relatives des partenaires :
Souvent, les individus sont de tailles différentes. L’hôte est de grande taille et le symbionte est de petite taille. L’hôte n’est pas toujours fournisseur.
Ex : Chez le lichen (champignon + algue), le champignon est l’hôte et l’algue est le symbionte.
Nécessité du symbionte pour le partenaire
La symbiose est dite « obligatoire » pour les organismes ne pouvant pas se reproduire sans leur partenaire. Cependant, elle peut être facultative si l’organisme peut s’en passer même temporairement. Mais la différence entre les deux n’est pas nette car il existe des obligations écologiques.
Il existe aussi une obligation physiologique impliquant un déplacement du partenaire car il ne peut pas se nourrir et survivre dans le symbionte alors que celui-ci peut vivre en s’isolant vis-à-vis de son hôte.
Dans tous les cas, un symbionte hors de son hôte est un « symbionte libre » et un hôte ayant libéré son symbionte est dit « hôte asymbionte ».
Il existe deux sortes différentes de transmission entre l’hôte et le symbionte :
- La transmission directe, elle a lieu quand l’hôte par ses gamètes ou pas un mécanisme transmet le symbionte à ses descendants.
- La transmission indirecte, elle a lieu quand l’hôte et le symbionte doivent se rencontrer par eux-mêmes.
Le symbiote et l’hôte se complète de plusieurs différentes manières.
Ex : Chez les lichens, l’algue apporte la nourriture au champignon grâce à la photosynthèse et le champignon apporte le support, l’humidité et les sels minéraux à l’algue
Les mycorhizes
C’est l’association entre une plante qui a un rôle d’hôte et un champignon ayant le rôle de symbionte.
Une partie du mycélium se trouvera dans la rhizosphère* (rhizome => racine aérienne) de la plante et une partie en contact avec une grande partie du sol.
Pour la plante, cela permet d’avoir une grande quantité de sol exploité sans développer des racines. Elle se développera donc plus qu’une plante non mycorhizée (non champignons).
Ces symbioses ont lieux dans tous les types de milieux que les sols soient secs, humides, pauvres, ou riches, …
Après 40 jours d’infestation par un champignon, un individu mycorhizé voit son poids doublé par rapport à un individu non mycorhizé, 60 jours après son poids est multiplié par 6.
On a donc une relation symbiotique exponentielle.
Dans le cas du phosphore, les champignons explorent grâce à leur mycélium, une zone 10 fois plus grande de sol que les racines de l’hôte. De plus, ils sont capables de prélever les phosphates à des concentrations plus faibles que les racines. Et enfin, ils vont, en plus, concentrer les phosphates dans leurs tissus.
Ex : Le peuplier dans un sol sans phosphate ne manquait pas de phosphate, grâce aux champignons qui le prélevaient dans son milieu phosphaté et le donnaient à l’arbre.D'autre part, les mycorhises sont de véritables accumulateurs d'ions minéraux ( absorbant massivement des sels libérés massivement après la chute des feuilles ).
Bien sûr, réciproquement l'autre partenaire apporte d'autres sustances au champignons ( substances vitaminiques ,...)
Bactériorhizes : Autre cas bien connu de la symbiose
Bactéries → nodosités sur les racines de Papilionnacées qui sont des zones abritant des bactéries du genre Rhyzobium .(application agronomique).
Dan d'autres cas, ce sont des Actinomycètes qui remplacent les bactéries : Aulnes, Argousiers ( Hippophae rhamnoïdes ), Myrica gale. Cas particulier : parfois soudure de racines entre arbres de même espèce : → connexions anatomiques de plante à plante.
On a parfois constaté que les racines d'un seul individu peuvent apporter de l'eau et d'autres substances à l'ensemble d'un peuplement.
III. Les relations indirectes entre plantes
Un des deux végétaux modifie le milieu dans lequel l'autre vit.
Ils sont du type concurrence, et parfois d'entraide.
a) les sécrétions des plantes
- Des expériences assez simples ( essais en germoir réalisés par FROSCHEL) permettent de le montrer → substances freinantes agissant sur semences de betteraves et de trèfles.
- Les agriculteurs savent depuis longtemps qu'on ne peut cultiver la même espèce sur la même parcelle → imputé à des sécrétions radiculaires accumulées dans le sol.
- On constate parfois que la plantation de plantes étrangères à un milieu ( ex : Robinier faux Acacia dans les Alpes maritimes pour fixer des sols riches en calcaire ou en dolomie , ce qui provoque un enrichissement du sol en azote et fait apparaître en grande quantité une diversité de plantes nitrophiles - Orties dioïques, Gaillet gratteron, Bryonne dioïque - alors que ces plantes étaient absentes du même milieu ) peut modifier complètement la composition du groupement végétal de départ.
- Les graines de beaucoup de plantes parasites ne germent qu'au voisinage immédiat des racines d'une plante d'une espèce ( ou groupe d'espèces ) déterminé : ces espèces libèrent des substances stimulant sélectivement la germination pour les semences du futur parasite.
Ex : nombreuses espèces d'Orobranches : Les Orobanchacées ( Ordre des Scrophulariales ) regroupent des plantes herbacées, sans chlorophylle, ni vraies racines et de fait, se trouvent condamnées à parasiter d’autres plantes.Si quelques espèces d’orobanches semblent assez spécifiques d’un hôte, beaucoup semblent davantage « spécialistes » d’une famille : Orobanche ramosa, par exemple, parasite les solanacées ; O. alba, les lamiacées et O. minor, les légumineuses.
- Il y a aussi des sécrétions ( minérales ou organiques ) potentielles par les organes aériens
Ex . eau de pluie récoltée en pleine forêt plus riche en éléments minéraux que celle récoltée à l'air libre : il y a lessivage des feuilles et dans une moindre mesure intervention des poussières atmosphériques.
Exemple cité par DENAYER - DESMET
| AIR LIBRE | SOUS COUVERT FORESTIER |
| Teneur en K+ : 0,1-2,2 mgrs/litre | 2,5-42 mgrs/litre |
| Teneur en Ca++ : 4-14 mgrs/litre | 9,1-53 mgrs/litre |
En automne et en régions tempérées : beaucoup d'éléments minéraux sont enlevés aux feuilles et restitués au sol par les pluies.
Les Spaignes contribuent à l'acidification des tourbières dont elles sont un élément de formation.
 Sphaignes et Sphaignes mi-immergées.
Sphaignes et Sphaignes mi-immergées.
b) La compétition passive entre plantes
Les plantes qui croissent ensemble sont en compétition.
- pour la lumière
- pour l'eau du sol
- pour les éléments minéraux
- pour l'espace vital
Une expérience facile peut être faite : la même espèce dans des bacs différents demée à des densités différentes, les autres conditions étant égales, on constate que :
- Le poids individuel des plantes diminue avec l'augmentation de la densité
- Le poids total est identique pour les différents bacs
- La mortalité ultérieure sévit le plus là où il y a la plus grande densité, d'où diminution de plants jusqu'à homogénéisation du nombre de pieds
La lumière est également un point important à examiner, on distingue :
- Les héliophytes ou plantes héliophyles, elles recherchent la lumière directe.
- Les sciaphytes ou plantes sciaphiles, qui peuvent croître en sites peu éclairés.
En forêt, il y a diminution progressive de la lumière, de haut en bas, chaque "strate" en consomme de plus en plus → testent +/- 2% au sol dans une vieille futaie. Ceci veut dire que beaucoup de plantes et et de très nombreuses espèces meurent au stade herbacé.
Ceci veut dire qu'un sol vierge de végétation est à l'inverse essentiellement formé de par des plantes héliophiles. Celles-ci disparaitront progressivement à mesure que le couvert se refermera → plantes de plus en plus sciaphiles. Ultérieurement, la disparition de plantes sciaphiles sur de grandes surfaces a lieu à la suite : d'incendies, tornades, ou chute d'un grand arbre etc... ce qui ramènera des essences héliophiles.
L'eau du sol, les éléments nutritifs
Cette concurrence aboutit à une stratification des organes souterrains ; les plantes qui peuvent vivre ensemble sont complémentaires en ce sens que les domaines parcourus par leurs racines sont distincts.
c) Importance des plantes dominantes
Au bout d'un certain temps, une espèce ou plusieurs espèces sont dominantes sur une parcelle donnée. L'espèce ou les espèces dominantes influencent définitivement la composition de la Communauté végétale.
Ex : Hêtres dominants dans une vieille futaie → microclimat tempéré, humide et ombragé → disparition des héliophiles au profit des sciaphiles → sécrétions foliaires et radiculaires qui influencent leurs espèces compagnes. Si le sol est pauvre en bases : mauvaise décomposition des organes morts qui s'accumulent et isolent le substrat minéral; ceci empêche ou inhibe la germination de certaines espèces et en particulier du Hêtre, d'ou la Hêtraie se tue d'elle même .
d) les facteurs du milieu physique et chimique
Il s'agit de : la température, l'eau, l'éclairement, les vents, la composition chimique du sol, la structure physique de celui-çi. Tous ces facteurs agissent simultanément. Abordons les successivement.
I. Facteurs climatiques : par exemple l'alternance des saisons, la durée des jours et des nuits, la température de l'air qui circule entre les organes aériens des plantes, température du sol ( racines !) hauteur des précipitations et leur répartition, importance de l'évaporation, neige, brouillard, vents ( direction, puissance ) ; lumière.
2. Facteurs orographiques
Le formes du relief influencent la température, la circulation de l'eau et la luminosité .
3. Facteurs édaphiques
Composition chimique du sol , humus, texture, structure, etc ...
Mais ces facteurs dépendent à leur tour de la température, de l'eau qui tombe au sol, de l'eau qui y circule, de la topographie.
Remarques :
a) Une hiérarchie existe dans les facteurs du milieu, surtout lorsque l'un d'eux existe au minimum :
ex : les précipitations :
Si très faibles → désert
Si variations locales → différences dans la composition floristique comme par ex. 2 versants opposés d'une même colline
Si faibles variations dans des précipitations abondantes ( ex > 2000 mm) alors il n'existera pas de différences dans la composition floristique .
Il s'agit de "facteurs limitants" : précipitations, la teneur en sels , etc ... ( ces facteurs du milieu jouent un rôle essentiel dans certaines communautés végétales ).
b) Il se peut que des facteurs différents produisent le même effet sur la végétation. Ex. l'augmentation des pluies provoque l'apparition de groupements végétaux différents dans une région où les pluies sont insuffisantes. On conçoit aisément que l'on peut atteindre le même résultat que si la nappe phréatique est plus superficielle dans la même région.
- Ex. plantes succulentes ( type Sedum ) apparaissent soit sur milieux très secs ( vieux murs etc...) ou sur sol sablonneux filtrant.
c) l'âge de la plante intervient : certaines plantes appellent des soins quand elles sont jeunes ; après ce n'est plus nécessaire .
Ex : buis dans les jardins, cimetières mais par contre les plantules de ce même buis sont fragiles...
IV . LES FACTEURS CLIMATIQUES
1. Le climat général, ou macroclimat ou climat régional
Est définit par des appareils : ex. thermomètres sous abri, à 2m au dessus d'un espace vert; pluviomètres; insolation, vitesse et direction des vents, etc...
Les mesures sont réalisées en conditions peu naturelles, mais uniformisées (afin que les résultats soient comparables ) : Il faut des mesures régulières et continues étalées sur une longue période ( résultats statistiques, avec moyennes " significatives " ).
Remarques :
-a) Ces indications définissent le climat général et intéressent surtout les phytogéographes ( grande région naturelle ), mais elles n'apprennent rien au niveau de la plante.
-b) Mais les caractères du climat général ne sont pas tous introduits dans les moyennes calculées par les météorologistes alors qu'ils ont une importance pour la végétation ( ex. rythmes quotidiens et saisonniers de l'éclairement et de la température, photopériodisme , iportance du nombre de jours froids influençant la dormance des bourgeons, des graines ). Cela explique la venue en végétation , la venue en fleur des plantes équatoriales, pour lesquelles le jour égale la nuit durant toute l'année; d'autres réclament des jours longs et des nuits courtes ; nécessité de périodes froides pour certaines plantes etc...
2. Les microclimats.
Un microclimat, est celui qui règne au niveau de l'être vivant, de la plante, de l'animal. Plus exactementc'est celui qui existe entre les bourgeons terminaux d'une plante et les extrémités radiculaires. Le volume ainsi défini varie d'une formation à l'autre, et il n'est pas constant entre la pointe d'une racine et l'extrémité d'un sommet végétatif de bourgeon.
Il y a donc des microclimats ( qui s'écartent du climat général parce qu'ils résultent principalement des modifications apportées par la présence de masses végétales ) selon les strates etc ...; ils sont atténués ou exacerbés par rapport au climat général ( ex pelouse <--> sous bois); explique la succession des aspects saisonnier ( qui dépend surtout des variations du microclimat thermique et lumineux au niveau du sol).-Ex. début du printemps, dans certaines forêts d'Europe Occidentale : floraison précoce de plantes à rhyzomes, bulbes etc : Ficaire, Perce-neige, Anémone des bois, Primula elatior, Ail aux ours, Jacinthe,Jonquilles... avant l'ouverture des bourgeons des arbres. Plus tard, apparition des sciaphiles, lorsque le couvert est refermé: ( Graminées ).
3. Les microclimats lumineux --> l'éclairement au niveau des plantes . Toutes les espèces n'ont pas les mêmes exigences . On distingue :
a) les héliophyles : végètent exclusivement en pleine lumière d'où prairies naturelles, clairières, sols exposés au sud, dunes ; il y a beaucoup d'adventices dans nos cultures ainsi que sur le bord des chemins : ce sont des héliophytes.
Les scyaphytes, peuvent végéter et se reproduire dans des milieux où la quantité de lumière est faible ( ex Lunaire vivace et Cardamine impatiente peuvent vivre parfaitement dans des milieux où n'existe qu'un nonantième de la lumière totale.).
b) Certaines plantes ont un comportement qui dépend de la quantité de lumière reçue . ex. Lamier jaune : forme des colonies avec rameaux étalés sans fleurir sous un taillis dense. Si la lumière augmente, --> les tiges se redressent et fleurissent.
c) Conséquences --> les héliophytes et les sciaphytes se groupent.
- si on a une seule strate, on a généralement des héliophytes.
- si plusieurs strates : la strate supérieure est composée d'héliophytes; les strates basses --> héliophytes; héliophytes et scyaphytes sont aussi une notion liée à l'âge des plantes ; Les épiphytes sont généralement situées dans des branches et troncs, là où une forte humidité atmosphérique règne et un air pur. On peut extrapoler cette notion à la répartition des mousses et des lichens en forêt, dans les forêts denses : mousses en épais manchon sur les troncs plus ou moins hauts et si le microclimat est plus lumineux --> lichens foliacés, mousses et hépatiques entre les écorces d'arbres. Dans les parties des cimes les plus éclairées et en lisières -->lichens pendants. Dans les lacs existet aussi une zonation de plus en plus nette depuis les rives jusqu'au centre, plus profond.
d) La qualité de la lumière est aussi modifiée après qu'elle a traversé un couvert végétal ou une nappe d'eau. ex : en forêt : le rouge et le bleu de la partie visible du spectre sont fortement absorbés par les feuilles : le vert l'est moins.
ex : champs de maïs : 7,5% du rayonnement visible et 19% des infrarouges , l'importance des deux rayonnements entre le pied des plantes et leur sommet est évidente.
e) Conséquence : les pertes d'énergie lumineuses par absorption font qu'il y a une diminution importante de l'énergie pour les plantes au niveau du sol
4. le vent
le vent cause des effets négatifs, mais aussi des effets positifs. Voyons les effets négatifs :
- La violence : bris, dessication d'organes délicats, déformations diverses, déformations diverses, déplacements de particules solides ou liquides, sable, limon,eau salée, grelons, floconsde neige, embruns ou bords de mer. Certains endroits sont particulièrement agressés par les vents ( sommets de montagnes, bords de mer, certains versants, certaines vallées,...). Il arrive que la prise du vent est parfois telle que la végétation se limite à des branches basses, collées au sol, même dans des régions où la croissance d'arbres serait possible. De plus il y a des déformations de port ; laminage des cimes, excentricités, aspect buissonnant et assymétrique.
Ajouter encore : le déchaussement des plantes , leur ensevelissement. Le vent provoque des érosions, des entailles de rives,etc..., en fonction de la morphologie de celles-ci, selon qu'elles sont enrichies en matières organiques ou pauvres ... C'est aussi le vent à l'occasion des tornades , provoque des hécatombes de végétaux ---> nouvelle colonisation par des héliophiles.
Les effets positifs : renouvellement de l'air, brassage des pollens, dissémination des semences , dessication des feuilles le matin ( maladies ! )
5. La neige = Ecran entre l'atmosphère et le sol.
Permet la survie de nombreuses plantes basses en cas d'hiver rigoureux . ex; pelouses recouvertes de neige en décembre environ de Bruxelles . epaisseur de 13 à 16 cm et temps serein ( D'après DE BACKER )
|
|
26 décembre | 30 décembre |
|
T° min air T° max air à - 5 cm à - 20 cm à - 100 cm |
- 7,5 °C - 0,5 °C + 0,2 °C + 1,5 °C + 5,7°C |
- 17,3 °C - 6,5 °C 0,0 °C + 1,5 °C + 5,7 °C |
Dans un sol nu voisin et non couvert de neige, pénétration du gel ( = 0°C ) à - 35 cm .Dans un sol nu voisin couvert de gazon mais non couvert de neige, le gel ne pénètre ne pénètre pas plus profond que 13 cm . Dans les deux cas, après 3 semaines de gel. En outre, la neige doit fondre rapidement au printemps surtout en montagne, sinon risque de retard dans la photosynthèse --> d'où les exposition sud sont généralement occupées par des héliophiles termophiles, riches en espèces et individus <--> exposition nord : les pentes ombragées en général et les lieux où la neige s'accumule et persiste longtemps .
Constatation : des groupements apparaissent en auréoles +/- concentriques autour d'une neige centrale qui fond lentement, c'est à dire :
- d'abord : végétation en croûte de qq mm d'épaisseur ( = hépatiques ).
- ensuite : végétation progressivement plus développée avec, à l'extérieur : le saule herbacé ( Salix herbacea ).
6. L'alternance du gel et du dégel
Surtout importante aux hautes altitudes et latitudes.
a) Fragmentation des roches --> pierriers qui ne sont colonisés que par peu de plantes très adaptées ( ex. Oxyria digyna = Polygonacée ).
b) Mouvements de solifluxion/La solifluxion désigne originellement un type de reptation du sol que l'on rencontre dans les régions où la terre gèle à une très grande
profondeur.Lors de la saison chaude, la partie superficielle du sol, en fondant, chemine vers le bas des pentes sur les matériaux toujours gelés. Ce sol, en état de liquide visqueux, peut descendre une pente aussi faible que de 2 ou 3 degrés et transporter des roches d'une taille considérable, qu'il tient en suspension.
Plus largement, on désigne par solifluxion tout écoulement superficiel des sols. Outre le transport de roches, parfois extrêmement volumineuses - la solifluxion est à l'origine de la plupart des chaos granitiques - la solifluxion a pour effet d'infléchir le port des arbres jeunes et encore mal enracinés. Le glissement, certes lent, mais continuel, tend à obliquer la tige de la jeune plante laquelle, par géotropisme négatif, cherche à se redresser verticalement. Cette lutte permanente a pour effet de donner une forme courbée très caractéristique à la base de la plante dont l'observation atteste sans aucune ambiguïté qu'elle croît sur un sol instable. Cette forme n'est pas sans rappeler celle que prennent les bouleaux tortueux (Betula tortuosa) que l'on rencontre dans les régions septentrionales. (Voir Bétulacées).
L'observation de grands mouvements de solifluxion se fait aisément en constatant des nappes de charriage superficielles à la surface des sols, un peu comme les marques que laissent les troupeaux de vaches sur des versants pentus.
Voiçi ce que wikipédia en dit :
La solifluxion est la descente, sur un versant, de matériaux boueux ramollis par l'augmentation de leur teneur en eau liquide. Quand l'eau liquide provient du dégel on parle de gélifluxion. Pendant la saison chaude la couche en surface (ou couche active) se met à fondre et glisse littéralement sur la couche inférieure solidifiée par le gel à une vitesse de 0,5 à 1,5 cm par an. En Allemagne, les depôts de solifluxion datant du dryas récent ont une épaisseur homogène de 0,4 à 0,7 mètres.
Les dépôts en bas des pentes qui résultent de la solifluxion sont appelés colluvions. Contrairement à d'autres types de glissements de terrain tels que les coulées de boues après des pluies torrentielles ou les coulées de débris, le processus de solifluxion est graduel, voire imperceptible. On parle de coulée, de chenal et de loupe de solifluxion, de lobes, de terrasses, ou de glissements en pieds de vache.
Mouvements de solifluxion vers le bas, si le substrat est constitué de terre fine et que le gel a pénétré à grande profondeur. certaines plantes peuvent s'accrocher au substrat mouvant et freinent, par leur lacis radiculaire, le mouvement descendant, d'où un relief en bourrelets et palliers successifs ( = marches d'escalier )
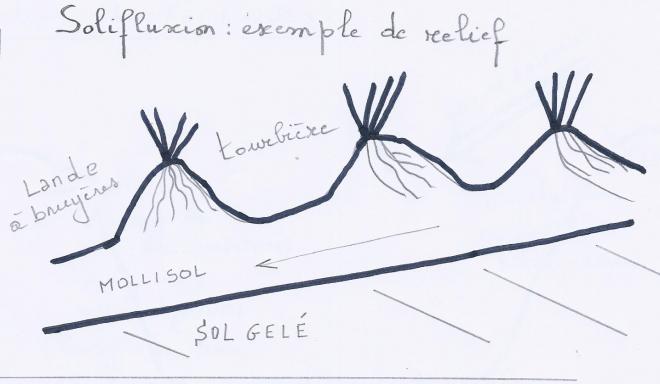
Sur ce relief particulier --> pentes abruptes +/- 1m séparant des palliers. Sont bien drainés et sont des milieux occupés par une lande. --> surfaces en pente douce , longtemps humides --> parfois tourbières en miniature.
c) En sol horizontal, il y a des mouvements de convection au dégel. Ces mouvements se traduisent par la formation de sols polygonaux : des éléments de calibre différents sont présents dans la masse pâteuse étalée sur un sous-sol imperméable. De tels sols sont généralement nus parce que la végétation ne peut s'y enraciner.
Explication des polygônes : climat périglaciaire - ex . Islande : les fragments rocheux débités par le gel sont mis en mouvement lorsque la partie superficielle du sol est dégelée : ils sont triés dans le sol mobile . Dans certains cas, les blocs les plus volumineux sont au centre des polygones, tandis que la périphérie est occupée par des éléments fins. NB : Il peut également y avoir des sols réticulés, les particules fines au centre et les grossières à l'extérieur.
d) Dégel brusque en montagne : --> avalanches qui se répètent souvent au même endroit d'où corridors non boisés avec groupements végétaux spécialisés.
Date de dernière mise à jour : 05/07/2021
Ajouter un commentaire