
12. Causes premières de la variation
Les mécanismes de la variation au niveau génétique.
Un préjugé négatif accompagne souvent le terme de mutation car il évoque en premier lieu quelque chose d'anormal. Il faut dire que la science-fiction avec son lot de mutants en tout genre a largement contribué à l'ancrage de ce préjugé. Or si certaines mutations sont préjudiciables pour l'individu qui les porte, d'autres sont à l'origine d'innovation génétique. En effet, les mutations, par définition, sont sources de variabilité et donc d'évolution. La variabilité étant le matériau brut de ce processus.
Il est fortement recommandé de se remémorer les principes de la de la biologie moléculaire par ex :http://www.vdsciences.com/pages/sciences-biologiques/biologie-moleculaire/ dans ce site
I. RAPPELS
I.1. Rappel de notions :
- Revoir la structure des chromosomes :
Un chromosome (du grec χρώμα, couleur et σώμα, corps, élément)[1] est un élément microscopique constitué de molécules d'ADN, de protéines histones et non-histones. Il porte les gènes, supports de l'information génétique, transmis des cellules mères aux cellules filles lors des divisions cellulaires.
Dans les cellules eucaryotes, les chromosomes se trouvent dans le noyau. Dans les cellules procaryotes, les chromosomes se trouvent dans une région du cytoplasme appelée nucléoïde.
Entre deux divisions, les molécules d'ADN constituant les différents chromosomes d'une cellule ne sont pas visibles ; ADN, ARN et protéines forment un ensemble non structuré appelé chromatine. L'ADN se condense progressivement au cours de la division cellulaire pour prendre lors de la métaphase une apparence caractéristique en forme de bâtonnet, de X ou de V.
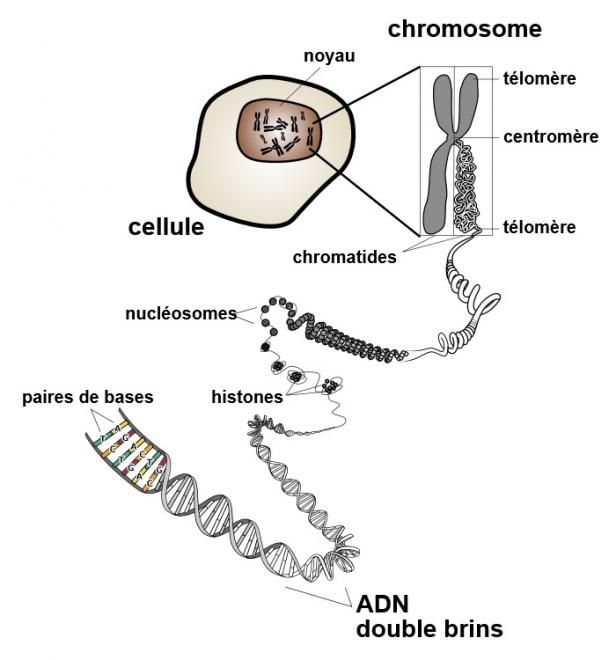
L'ensemble des chromosomes est représenté sur un caryotype, ou carte de chromosomes, où les chromosomes sont habituellement présentés par paires, en parallèle avec leur homologue.
- Position du centromère
Les chromosomes eucaryotes peuvent être distingués selon la position de leur centromère : On parle de chromosome métacentrique lorsqu’il possède un centromère en position centrale (position médiane) ce qui lui donne des bras (ou chromatides) de longueur à peu près égales. Un chromosome submétacentrique est un chromosome dont le centromère est presque en position centrale ; les chromatides de ce chromosome présentent des bras de longueur inégale (un petit bras nommé « p » et un long bras « q »). Si le centromère est plus proche de l’une des deux extrémités (les télomères), le chromosome est dit acrocentrique. Enfin, un chromosome télocentrique présente un centromère très proche de ses télomères. En cas de perte du centromère (anomalie), le chromosome est dit acentrique. D’autres anomalies peuvent provoquer l’apparition d’un chromosome possédant deux centromères nommé chromosome dicentrique. Celui-ci est instable et peut se casser (lors de la méiose) en différents segments qui se répartissent au hasard dans chacune des cellules filles.
- Présence ou absence de satellite
L’ADN satellite se présente sous la forme de séquences d’ADN pouvant atteindre plusieurs millions de paires de bases et constituées d’un grand nombre de répétitions d’une séquence beaucoup plus petite (l’unité de répétition). La longueur de l’unité de répétition est variable d’un type d’ADN satellite à l’autre, de 5 paires de bases pour les satellites III humains à 755 paires de bases pour les satellites trouvés dans les génomes des plantes.
Le nom « ADN satellite » vient de ce que l’ADN de ces séquences, en raison de leur nature répétitive, a une composition en nucléotides fortement biaisée par rapport au reste du génome ; en conséquence, il sédimente différemment et forme une bande secondaire, dite « satellite », dans un gradient de chlorure de césium.
Le génome humain comprend environ 7 % d’ADN satellite.
L’ADN α-satellite (aussi appelé alphoid DNA en anglais) est constitué de répétitions d’un motif de 171 paires de bases. On le retrouve au niveau du centromère de tous les chromosomes, où il s’étend sur des régions allant de quelques centaines de kilobases à plusieurs mégabases — le nombre de répétitions et la taille totale étant variables d’un chromosome à l’autre et d’un individu à l’autre. L’ADN α-satellite contribue à la fonction du centromère, en permettant notamment le recrutement de la protéine CENP-B (Centromeric protein B) par l’intermédiaire d’un motif conservé de 17 paires de base. Néanmoins, il n’est pas indispensable à la bonne fonction du centromère, des néocentromères pleinement fonctionnels pouvant être observés sur des régions génomiques dépourvues d’α-satellites.
L’ADN β-satellite est constitué de répétitions d’un motif de 68 paires de bases. Il s’étale sur des régions de 50 à 300 kilobases sur les bras courts des chromosomes acrocentriques (chromosomes 13, 14, 15, 21 et 22), sur les péricentromères des chromosomes 1, 3 et 9 ainsi que sur le bras long du chromosome Y.
L’ADN satellite III est constitué de répétitions d’un motif de 5 paires de bases (GGAAT), occupant plusieurs centaines de kilobases dans la région péricentromérique de la plup
- Chromomères :
Fragment d'un chromosome. Avec certaines colorations, les chromosomes apparaissent comme une succession de bandes (en réalité de disques) sombres colorables et de bandes claires non colorables. Les chromomères sont ces disques sombres, qui représentent un fragment de la double hélice d'ADN (avec les histones, protéines associées), donc un certain nombre de gènes.
- Chromosomes polytènes
Les chromosomes polytènes (ou polyténiques) sont présents dans les cellules de certains organes des larves de Diptères (les glandes salivaires par exemple). Ils correspondent à un certain nombre de copies des chromatides (jusqu'à 1 024) qui sont restées soudées entre elles (phénomène d'endoréplication). La coloration de Feulgen rend visible, au microscope photonique, des bandes fortement colorées, correspondant à des régions de chromatine condensée (appelées hétérochromatine), et des bandes plus claires (renflements ou « puffs »), les régions moins condensées de la chromatine (appelées euchromatine).
On appelle « puffs » les régions diffuses relâchées du chromosome polyténique. Ce sont des sites de transcription active en ARN. Un anneau de Balbiani est un puff de grosse taille sur le chromosome. Il tire son nom de son découvreur Édouard-Gérard Balbiani (1823-1899).
Les chromosomes polytènes ont été largement étudiés par Calvin Blackman Bridges, collaborateur de Thomas Hunt Morgan.
- Les inducteurs des remaniements ou mutations chromosomiques
Rayonnements ionisants ( rayons X,... ), ou non ionisants (UV,...), la radioactivité est également mutagène
substances chimiques dites radiomimétiques ( Ypérite )
Autres agents chimiques. Quelques exemples peuvent être cités :
- le benzopyrène, présent dans la fumée de cigarette et autres résidus de combustion incomplète ;
- le bromure d'éthidium, substance très utilisée en laboratoire de biologie moléculaire ;
- le dichlorométhane, solvant ;
- le diéthylpyrocarbonate, autre produit très utilisé en laboratoire de biologie moléculaire ;
- l'éthanal ;
- le trichloréthylène, solvant très utilisé.
- si une cassure a lieu lors de la mutation, trois possibilités :
- Retour à l'état initial, grâce à un mécanisme réparateur.
- Rupture définitive .
- Recollement dans un état différent de l'initial.
I.2. Rappel mitose
La mitose est le processus de reproduction conforme (à l'identique ) des cellules. A l'issue de la mitose , la cellule mère s'est divisée en cellules filles identiques entre elles et identique à la cellule qui leur a donné naissance.
Cycle cellulaire :
Interphase :
L'interphase est divisée en 3 phases G1-S-G2 (G1et G2 sont des phases de croissance et S une phase de synthèse d'A.D.N. durant laquelle a lieu la réplication de L'A.D.N. )
Mitose: Celle ci peut être décomposée en 4 phases.
1. Prophase :
Disparition de la membrane nucléaire. Les chromosomes deviennent visibles . Ils sont formés de deux chromatides (2 molécules d'A.D.N. double brin) reliés par le centromère dans lequel l'ADN se trouve dans une forme enroulée et compactée. A partir des centrosomes se forme un réseau de microtubules allongés qui va composer le fuseau mitotique . Celui ci va diriger le déplacement des chromosomes.
2. Métaphase :
Les centromères s'alignent au centre du fuseau mitotique (plaque équatoriale) avec une chromatide de chaque côté.
3. Anaphase :
Clivage et séparation des centromères.Les chromatides sœurs migrent vers les 2 pôles opposés de la cellule.
4. Télophase :
A la fin de l'anaphase lorsque les chromosomes ne se déplacent plus, apparaît un sillon de division qui va séparer la cellule en deux cellules filles. Le fuseau mitotique se défait, et il se forme une nouvelle membrane nucléaire.
Puis la cellule passe de nouveau en interphase et le cycle recommence.
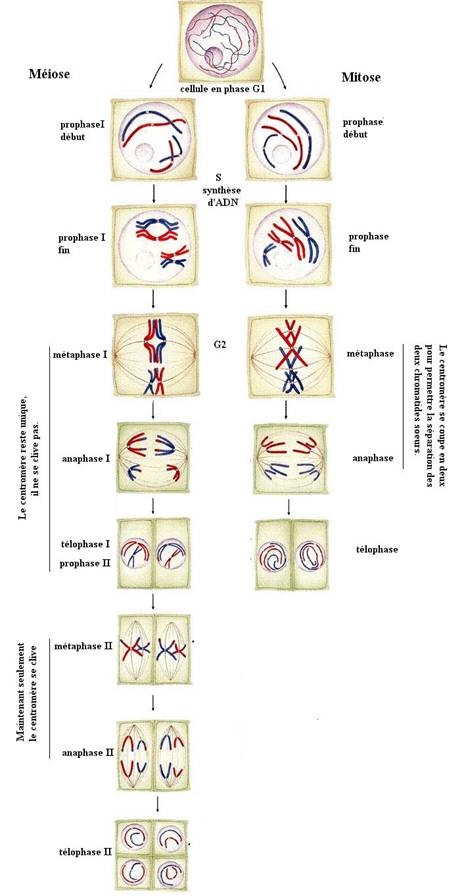
I.3. Rappel meïose
Elle fait suite à la réplication de l'ADN. Elle permet le passage d'une cellule mère dite diploïde à 2n chromosomes homologue doubles à deux chromatides à 4 cellules fille haploïdes à n chromosomes simples.
Première division méiotique : division réductionnelle .Elle comporte quatre phases de toute division cellulaire mais néanmoins présente des particularités.
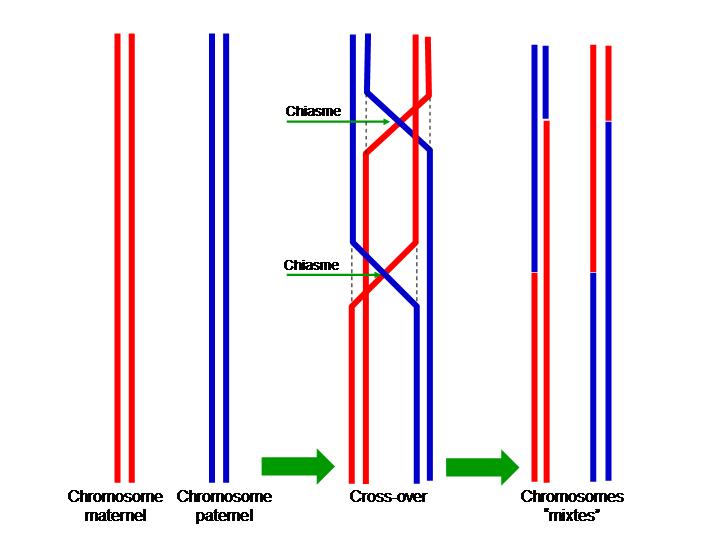
L'enveloppe nucléaire disparaît. Les chromosomes bichromatidiens s’individualisent par condensation de leur ADN à partir de la chromatine du noyau. Ils apparaissent doubles formé chacun de deux chromatides. Ils s’associent ensuite par paires de chromosomes homologues. Cet appariement donne des tétrades : 4 chromatides et des bivalents : 2n chromosomes homologues. Cette phase est divisée en cinq étapes qui correspondent à cinq états caractéristiques de la chromatine : leptotène, zygotène, pachytène, diplotène et diacinèse.
- Leptotène : (Du grecque Lepto- mince et -tène bandelette). Début de la condensation de la chromatine et attachement des télomères (extrémités des chromosomes) à l'enveloppe nucléaire par la plaque d'attache. Les chromatides s'associent par chromatides sœurs. Les deux brins provenant du père ensemble (bleu) et les deux brins provenant de la mère ensemble (rouge).
- Zygotène : (Du grecque Zygo- liaison entre deux éléments). Début de l'appariement des chromosomes homologues (synapsis) par le complexe synaptonémal (ou synapton) et convergence des télomères vers le centromère (un peu comme une fermeture éclair). Le complexe synaptonémal est une structure complexe constituée d'un élément central, SYCP1 qui forme un homodimère, relié à deux éléments latéraux. Les éléments latéraux sont en fait les cohésines SMC1, SMC3 formant un hétérodimère maintenu en place par mREC8 hHR21. Les cohésines se trouvent de part et d'autre par des filaments transverses, à celles-ci se lie la chromatine de chaque zone des chromosomes impliqués dans le phénomène ultérieur d'enjambement (ou crossing-over). Il y a organisation « en bouquet » des chromosomes. L'ensemble des deux chromosomes homologues s'appelle une tétrade (car 4 chromatides) ou un bivalent (car 2 chromosomes).
- Pachytène (phase la plus longue ~ 2 semaines pour des spermatozoïdes humains) : (Du grecque Pachy- épais). Appariement strict des chromosomes homologues et apparition des nodules de recombinaison et de nodules tardifs qui permettent les enjambements (échanges entre chromatides homologues). Cette phase a une importance considérable dans le brassage chromosomique (crossing-over).
- Diplotène : (Du grecque Diplo- double). Désynapsis (séparation des chromosomes homologues), mais les chromosomes restent attachés en plusieurs points au niveau desquels deux des quatre chromatides semblent s'entrecroiser (chiasma). Pour le bon déroulement de la méiose il en faut au minimum un par chromosome, en moyenne 2-3. Il y a décondensation de la chromatine et formation des grandes boucles permettant un fort taux de transcription. Cette étape de la prophase I peut durer plusieurs années chez l'ovocyte. En effet au cours de ce stade, l'ovocyte I augmente de volume, la décondensation des chromosomes permet la synthèse d'ARN messager et d'ARN ribosomiques qui seront stockés dans le cytoplasme et serviront de réserve pour le futur zygote lors des premières division de segmentation.
- Diacinèse : (Du grecque Dia- à travers). Recondensation de la chromatine et détachement des télomères de l'enveloppe nucléaire. Glissement des chiasmata vers les télomères (terminalisation des chiasmata). À la fin, il y a disparition de l'enveloppe nucléaire.
Métaphase I
Les paires de chromosomes homologues (bivalents) se placent en vis-à-vis de part et d'autre du plan équatorial. Pour chaque bivalent, les centromères se placent de part et d'autre ainsi qu'à égale distance du plan équatorial. Leur orientation se fait de façon aléatoire : on appelle ce phénomène la « ségrégation indépendante ».
Anaphase I
La contraction de la fibre du fuseau achromatique entraîne la disjonction des deux chromosomes homologues de chaque paire et migrent aux pôles opposés, tirés par des microtubules kinétochoriens (microtubules accrochés à un kinétochore au niveau d'un centromère) dû à la dépolymérisation de tubuline. Il n'y a pas clivage des centromères, ceci est dû au fait que la séparase dégrade hREC8 (et donc la construction des cohésines) mais au centromère est inefficace étant donné que Sga1 y protège hREC8
Télophase
Les enveloppes nucléaires réapparaissent dans chaque cellule, il y a donc formation de deux cellules haploïdes à n chromosomes à deux chromatides (chromosomes bichromatidiens)(n chromosomes, 2n ADN). La cellule se divise en deux, grâce à un anneau contractile fait d'actine.
Deuxième division méiotique : division équationnelle
Chaque chromosome étant resté dupliqué (2n ADN), il n'y a pas de réplication de l'ADN et elle se déroule immédiatement. Cette division consiste en une simple mitose classique : pour chaque cellule, on passe de n chromosomes doubles à deux cellules à n chromosomes simples.
- Prophase II : Phase identique à la prophase I mais brève car les chromosomes sont restés compactés.
- Métaphase II : Les chromosomes se placent sur la plaque équatoriale par leur centromère. Leur condensation est maximale.
- Anaphase II : Les chromatides sœurs de chaque chromosome se séparent après rupture de leur centre et migrent vers des pôles opposés de la cellule tirés par les fibres protéiques.
- Télophase II (fin) : Les quatre cellules haploïdes issues de la méiose possèdent n chromosomes simples.
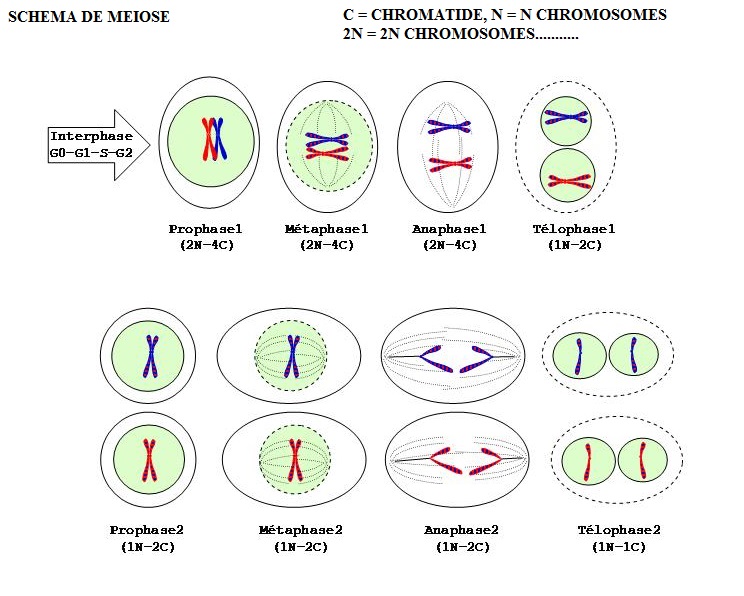
La diversité des gamètes
Les gamètes créés par la méiose sont différents bien qu'ils descendent de la même cellule. Cette différenciation joue un rôle clef dans l'évolution des espèces et leur diversité génétique.
Brassage allélique par ségrégation indépendante des chromosomes homologues (brassage inter-chromosomique)
(cas des espèces à caryotype 2n)
Un premier facteur de diversité facile à comprendre provient de l'attribution aléatoire des allèles, c'est-à-dire de chacun des deux chromosomes d’une même paire (chromosomes homologues) vers les cellules filles haploïdes. Au moment de la métaphase I de la méiose, les chromosomes se disposent aléatoirement de part et d’autre du plan équatorial. Chaque chromosome (allèle) d'une paire migre ensuite vers un pôle (anaphase I), sans influencer les sens de migration des allèles des autres paires. Chaque cellule fille possèdera donc un jeu de chromosomes (et donc de gènes) différent de celui de la cellule mère. Cette différenciation est appelée brassage inter-chromosomique.
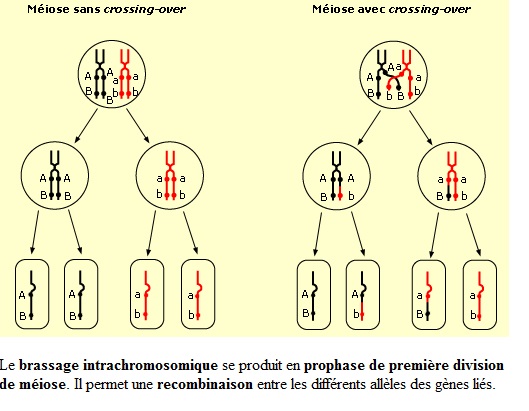
Échange d'allèles au sein d’une paire de chromosomes (brassage intra-chromosomique)
À chaque méiose, sauf cas exceptionnels (drosophile mâle par exemple), il peut se produire un échange réciproque de fragments de chromatides appartenant à deux chromosomes homologues : c’est le phénomène d’enjambement, également appelé crossing over, qui survient pendant la prophase I (donc avant la séparation métaphasique des chromosomes homologues). Cet enjambement est provoqué par un module de recombinaison (complexe multi-enzymatique). Les chromatides recombinées se distinguent des chromatides d'origine ; on parle alors de brassage intra-chromosomique.
C'est aussi durant cette période que des échanges par crossing-over asymétriques de fragments inégaux de chromatine peuvent être réalisés et donner naissance à des gènes dupliqués sur l'une des chromatides, faisant apparaître une famille multigènique. En anaphase I, lors de la disjonction des chromosomes, les deux chromosomes homologues de chaque paire se séparent. Chaque chromosome migre vers l'un ou l'autre pôle de la cellule.
C'est un phénomène aléatoire et le nombre de combinaisons ou lots possibles entre les n paires est infini : ainsi le chromosome d'une paire peut être associé avec l'un ou l'autre chromosome d'une deuxième paire ; ceci est valable pour les n paires. Un tel brassage est qualifié d'interchromosomique. Les différents chromosomes se séparent donc indépendamment les uns des autres : on parle même d'une « loterie » de l'hérédité. Le nombre de combinaisons possibles est de : 2n.
Dans le cas de l'espèce humaine, n = 23, donc un individu peut produire 223 spermatozoïdes ou ovules différents (soit plus de 8 millions de spermatozoïdes ou d'ovules différents).
La superposition des deux brassages permet une diversité considérable des gamètes.
- Si l'individu possède
 gènes hétérozygotes, le seul brassage intrachromosomique permet
gènes hétérozygotes, le seul brassage intrachromosomique permet  arrangements possibles.
arrangements possibles.
- S'il possède
 paires de chromosomes, le seul brassage interchromosomique permet
paires de chromosomes, le seul brassage interchromosomique permet  arrangements possibles.
arrangements possibles.
Anomalie de la méiose
- anomalie de nombre : non disjonction des chromosomes = aneuploïdie (par exemple la trisomie 21)
- anomalie de structure : translocation
- erreur de réplication de l'ADN : mutation
Mitose et méiose
La méiose et la mitose sont différentes en plusieurs points mais elles ont aussi de grandes similitudes. La mitose se produit au cours de la multiplication asexuée alors que la méiose a sa place dans la reproduction. Presque toutes les cellules peuvent subir une mitose alors que la méiose ne concerne que celles des organes de reproduction chez les espèces diploïdes (les ovogonies et les spermatogonies) ou la cellule-œuf chez les espèces haploïdes. À la fin de la mitose, il y a deux cellules génétiquement identiques alors qu'à la fin de la méiose il y a quatre cellules qui ne sont pas nécessairement génétiquement identiques.
Les étapes du cycle cellulaire : La méiose. On représente la cellule par un ovale et le noyau par un cercle vert. On ne montre qu'un type de chromosome. Les rayures bleues-rouges des chromosomes suggèrent le mélange des gènes paternels et maternels obtenus par l'enjambement en Prophase1.
II.INTRODUCTION
2.1 Mutations historiques
Le premier rapport de mutation a été fait par Sprenger en 1590 à propos d'une Chélodoine ( herbe à verrues, plante à petites fleurs jaunes ) qui présentait des feuilles laciniées ( fortement découpées ) au lieu de feuilles lobées . Un siècle plus tard, Marchand rapporte ( 1715-1716 ) la production de deux nouvelles formes de Mercuriale et disserte en 1719 dans les comptes rendus de l'académie des Sciences sur une rose monstrueuse et sur la production de nouvelles espèces de plantes. Gmelin en 1749 découvre 6 mutations nouvelles de Delphinium ( Pied d'Alouette ). Les Duchesne père et fils, horticulteurs versaillais, découvrent en 1763, une mutation affectant le Fraisier sauvage ( Fragaria vesca ) présentant des feuilles entières au lieu de feuilles trifoliées.
2.2 Mutations et mutationnisme
Cependant, le rôle des mutations fut totalement ignoré par les scientifiques du XVIIIème siècle. Ces variations aléatoires ne furent considérées que comme des curiosités de la nature, voire des monstruosités. En effet, le fixisme était encore fermement ancré jusque la moitié du XVIIIème siècle. Les espèces étaient considérées comme immuables, fixes au cours du temps. Selon les uns, les espèces apparaissaient par génération spontanée, selon les autres elles étaient le fruit d'une création divine. lorsque le fixisme s'effaça devant le transformisme avec les travaux précurseurs de Buffon puis de Lamarck, le rôle des mutations ne fut cependant pas considéré comme à la base des phénomènes évolutifs. Il fallut attendre la fin du XIXème siècle , pour que différents auteurs comme Bateson en 1894 prennent en compte l'existence de variations brusques, discontinues et héréditaires chez les organismes ( Bateson en 1906est à l'origine du terme de génétique pour désigner la nouvelle science de l'hérédité.°. En 1901 et 1903, Ugo De Vries publie les deux volumes d'un ouvrage intitulé " Die Mutationsthéorie ". Ses travaux portent sur une plante cultivée originaire d'Amérique, Oenothera lamarckiana. Il donne le terme de mutation aux variations brusques et héréditaires qui seraient, selon lui, à l'origine de la diversité des espèce. Le mutationnisme voit le jour . Cette théorie met en avant le rôle des transformations héréditaires dans le changement des espèces biologiques.
2.3 . Mutationnisme et sélection naturelle
Après les travaux de De Vries portant sur les mutations et la prise en considération des lois de Mendel établies en 1862 et redécouvertes indépendamment en 1900 par De Vries, Correns et Von tschermak, la génétique et la notion d'évolution des espèces ( Darwin a alors publié "l'Origine des Espèces, en 1859 ) vont devenir compatibles.
Cependant, deux théories s'affrontent. Les darwiniens défendent le rôle de la sélection naturelle dans le processus de diversification des espèces, tout trait biologique étant alors expliqué par ce processus,. En revanche Bateson et les premiers généticiens défendent la théorie du mutationnisme, les mutations pouvant expliquer à elles seules l'apparition de nouvelles espèces. La théorie du mutationnisme sera approfondie par les travaux de génétique de Thomas Hunt Morgan et de ses collaborateurs sur les drosophiles. De nombreuses expérimentations de croisement sont réalisées confirmant les résultats obtenus par Mendel. De plus, en utilisant les chromosomes géants de la drosophile, , les premières cartes chromosomiques sont établies et l'effet des crossing-over, à l'origine de l'échange de matériel génétique entre deux chromosomes homologues , est étudié. En 1915, Morgan et ses collaborateurs publient " The Mechanic of Mendelian Heredity ", puis en 1926 " The theory of the gene"
Il faudra attendre les progrès de la génétique des populations dans les anées 1930, pour que mutation et sélection naturelle trouvent leur place dans la théorie synthétique de l'évolution , exposée par George Gaylord Simpson en 1944 dans " Rythme et modalité de l'évolution".
Il est maintenant incontestablement établi que les mutations sont la source de la variabilité génétique , et que dans certaines conditions, la sélection naturelle " fait le tri" parmi les mutations , mais n'est pas génératrice de variabilité.
III. CARACTERISTIQUES DES MUTATIONS
III.1 Les mutations touchent le génotype d'un individu et peuvent affecter son phénotype
- On appelle mutation, tout changement, toute modification aléatoire touchant une séquence d'acide nucléique ou affectant l'agencement de gènes ou leur nombre.
- On désigne par le terme génotype la constitution génétique d'un organisme.On désigne par le terme phénotype l'aspect ou les caractéristiques d'un organisme résultant de l'interaction entre sa constitution génétique et son environnement . Ces caractéristiques peuvent être morphologiques, physiologiques , biochimiques ou comportementales.
D'un point de vue génétique , lorsque l'on se réfère au gène muté ou à une séquence contenant une mutation, on parle d'allèle muté par référence à un allèle sauvage, non muté. Si une mutation affecte la fonction d'un gène la désignation allèle muté/allèle sauvage est explicite. Cependant, dans bien des cas, la notion de mutation est relative et la terminologie " sauvage"/"muté" est arbitraire.
Un changement qui provoque le retour de l'allèle muté à l'allèle sauvage se nomme une mutation réverse, une réversion ou une mutation en retour .
Notons que le terme mutation désigne un processus mais est également utilisé pour désigner le produit de ce processus, c'est à dire la séquence , le gène ou le chromosome muté. Si tout changement de phénotype résulte d'une mutation affectant le génotype de l'individu, toute modification du génotype ne conduit pas nécessairement à un changement de phénotype. Cette mutation peut ne pas être visible phénotypiquement. Différentes raisons peuvent rendre compte de cette situation, notamment :
- Chez les organismes diploïde lorsque l'allèle muté est récessif.
- Dans le cas d'une mutation affectant une région codante lorsque la dégénérescence du code génétique prévient l'effet de la mutation ( Le code est dégénéré et redondant, voir biologie moléculaire http://www.vdsciences.com/pages/sciences-biologiques/biologie-moleculaire/bio-moleculaire-9.html)
- Lorsque la mutation intervient dans une région non informative ( non codante et non régulatrice )
- Dans le cas de maladies humaines lorsqu'il existe une pénétrance incomplète de la maladie ou une expressivité variable.
Un organisme dont le phénotype présente une modification est désigné sous le nom de mutant
De plus, une seule mutation peut provoquer une cascade de modifications pouvant entraîner des changements importants au niveau du phénotype en modifiant en même temps plusieurs caractéristiques de l'individu, on parle alors de mutations pléiotropes .
III.2. Les mutations peuvent être somatiques ou germinales .
Toutes les mutations ne sont pas héréditaires. L'hérédité d'une mutation dépend du mode de reproduction de l'organisme et du type de cellule mutée.
Chez les organismes unicellulaires, toute mutation viable atteignant la cellule sera transmise aux cellules filles issues de sa division.
Chez un organisme pluricellulaire, une mutation peut affecter soit une cellule de la lignée somatique , on parle alors de mutation somatique, soit une cellule de la lignée germinale, on parle alors de mutation germinale.
Si une mutation affecte la lignée somatique d'un organisme, lors d'une reproduction sexuée, la modification ne sera pas transmise à sa descendance et ne sera portée que par l'individu. En revanche , en revanche si l'organisme se multiplie par voie sexuée , une mutation somatique peut être transmise au nouvel individu, si la cellule mutée participe à la construction de ce nouvel individu.
Les mutations somatiques peuvent résulter de recombinaisons mitotiques . Quelle que soit la nature de la mutation somatique , la cellule mutée par divisions mitotiques conduit à des clones cellulaires. Par exemple chez les végétaux, une cellule somatique mutée peut être à l'origine d'une partie de la plante portant un phénotype différent des autres parties. Certaines mutations sont à l'origine de maladies humaines . Ainsi certains cancers sont dus à des mutations somatiques affectant des gènes dits proto-oncogènes impliqués dans le contrôle de la division cellulaire . Certaines maladies dites du vieillissement sont dues à des mutations somatiques affectant le génome mitochondrial
Si la mutation affecte la lignée germinale , cette mutation pourra être héréditaire à condition qu'elle soit viable. Pour être viable, la mutation ne doit pas entraîner la mort de la cellule ou de l'individu qui la porte , c'est à dire qu'elle ne doit pas être létale. De plus pour être héréditaire, cette mutation ne doit pas entraîner la stérilité de l'organisme qui la porte et doit affecter le matériel génétique d'un gamète impliqué dans la reproduction conduisant à un nouvel organisme.
Chez l'l'homme, 15 à 20 % des zygotes sont porteurs de génotypes létaux est se traduisent par des avortements spontanés dans les 20 semaines après la fécondation. De 60 à 70 % des foetus avortés présentent des anomalies du nombre ou de la morphologie des chromosomes.
Chez les eucaryotes métazoaires à rerpoduction sexuée une mutation, pour être héréditaire doit être portée par les cellules de la lignée séminale susceptibles de participer à la reproduction.
III.3. Les mutations peuvent être spontanées ou induites.
La fréquence de mutation est la fréquence à laquelle on retrouve une mutation donnée dans une population de cellules ou d'individus
Le taux de mutation se définit comme étant une évaluation de la probabilité qu'une mutation donnée se produise en un temps donné
Parmi les mutations, on distingue les mutations dites induites qui apparaissent suite à un traitement par des agents mutagènes et les mutations dites spontanées qui apparaissent en l'absence de traitement mutagène.
Les mutations spontanées résultent le plus souvent d'erreurs se produisant au cours des divisions cellulaires, lors de la réplication de l'ADN ou de la ségrégation des chromosomes . Au cours de la réplication des erreurs de copie ( remplacement d'une base par une autre ), un bégaiement de la polymérase ( insertion de bases ) ou des oublis ( délétion de bases ) peuvent se prouire. Cependant ces taux d'erreur de copie sont très faibles et sont de l'ordre de 10-5 à 10-7 cycles par site de réplication ou d'une moyenne de 10-6 gène gène/génération. Les mécanismes de correction abaissent ces taux à environ 10-8 chez Escherichia coli . Les erreurs de ségrégation des chromosomes donnat lieu à des remaniements chromosomiques varient de 4.10-3 à 1,3.10-4/gamète/génération.parmi ces mutations, les duplications jouent un rôle évolutif important. A ces mutations s'ajoutent les mutations d'insertion de séquences d'ADN par transpositions , comme l'insertion d'éléments transposables ou de rétropseudogènes . Le séquençage complet de certains génomes a mis en évidence l'importance de ces mutations . Certaines séquences sont plus susceptibles que d'autres aux mutations lors de la réplication . Ainsi les séquences de type microsatellite possédant un motif répédé subissent des dérapages réplicatifs conduisant à la délétion ou à l'insertion de répétition du motif . Les taux de mutation de ces séquences sont estimés expérimentalement à 10-2-10-6 kb.Ces régions n'étant en général pas codantes , les mécanismes de réparation ne sont pas mobilisés et le nouveau variant n'est pas éliminé. Du fait de leur polymorphisme, ces séquences ont utilisées comme marqueurs moléculaires. De plus l'expansion de triplets par ce même mécanisme est à l'origine de maladies génétiques humaines comme le syndrome de l'X fragile. Cette maladie est due à un allongement important d'une séquence répétée ( CGG) située dans la région 5'UTR du premier exon du gène du gène FMR1 ( pour "Fragile X Mental Retardation ")ce qui provoque une méthylation de ce gène qui n'est alors plus exprimé.
Des lésions spontanées affectent l'ADN avec une fréquence de 10-5 à 10-8 mutations/organismes /génération. Celles les plus fréquentes sont la désamination et la dépurination. La désamination conduit par exemple à la conversion d'une cytosine en uracile . La dépurination conduit à la perte d'une base purine . Certains sites de l'ADN sont plus fréquemment désaminés et constituent des points chauds mutationnels comme les vytosines méthylées. Certains radicaux libres comme les radicaux superoxydes, hydroxyles ou le péroxyde d'hydrogène qui sont des sous-produits du métabolisme sont à l'origine de mutations notamment impliquées dans les maladies du vieillissement.
De nombreux agents mutagènes augmentent les taux de mutation comme des agents chimiques ( bromure d'éthidium, acide nitreux, acridine, acridine orange ...) ou physiques ( rayons UV non ionisants, rayons X, α,β,γ, ionisants). Certains de ces agents mutagènes sont aussi des facteurs cancérigènes car induisant la formation de cellules cancéreuses . Les agents mutagènes peuvent avoir différents effets.
- Le bromure d'éthidium agit en modifiant l'espacement entre les paires de bases. On parle alors d'agent intercalant. On utilise expérimentalement la propriété de ces produits ainsi que sa fluorescence aux UV pour révéler la présence d'ADN.
- L'acide nitreux provoque l'incorporation de bases substituées dans l'ADN, c'est à dire de base où le groupement a été changé. Par exemple l'adénine peut être désaminée en hypoxanthine qui est un mimétique de la guanine , ou la cytosine en uracile qui est un mimétique de la thymine.
- L'acridine provoque la délétion ou l'insertion de petites séquences nucléotidiques.
- Les rayons ultra-violets provoquent des cassures d'ADN ou des changements dans les bases suite à l'absorption de l'énergie de la lumière. Les mutations les plus fréquentes sont la formation de dimères de pyrimidine notamment de thymines au sein du même brin d'ADN ou d'un brin à l'autre de la double hélice.
III.4. Les mécanismes de réparation des légions d'ADN sont variés
Les lésions de l'ADN provoquent des mésappariemments des bases ou une distorsion de l'hélice d'ADN ce qui peut bloquer la transcription et la réplication. Différents systèmes de réparation interviennent pour réparer ces différents types de lésions spontanées ou induites de l'ADN.Dans certains cas, une réversion est possible . Par exemple , les dimères de thymines formés par l'action des UV sont reconnus par différentes enzymes conduisant à une réversion des dimères par coupure de la liaison entre les pyrimidines du dimère.
Le système NER ( pour "Nucleotide excision Repair") permet la réparation d'un segment d'ADN endommagé par excision de nucléotides, re-synthèse de la séquence qui fait intervenir des ADN polymérase et ligation. De plus si le système NER n'est pas mobilisé, deux mécanismes post réplicatifs, un mécanisme de recombinaison et un mécanisme de synthèse translésionelle. Des erreurs dans la réparation de ces lésions d'ADN sont à l'origine de mutations.
Les lésions ponctuelles sont réparées par des systèmes spécifiques d'excision. Par exemple les ADN glycosylases excisent les bases endommagéesproduisant des sites apuriniques ou apyrimidiques (AP) réparés par le système endonucléase AP. Par ailleurs certains systèmes enzymatiques neutralisent les radicaux libres et préviennent ainsi certaines mutations . Ainsi la superoxyde dismutase catalyse les radicaux superoxyde en péroxyde d'hydrogène qui est ensuite converti par la catalase, en eau
IV. TYPES DE MUTATION
IV.1.Classification des mutations
Plusieurs termes sont utilisés pour évoquer les mutations
De manière générale on a quatre catégories :
1) Les mutations nucléotidiques : n'affectant qu'un nombre limité de nucléotides, on peut utiliser le terme mutation ponctuelle pour un nombre restreint de nucléotides ou pour 1 seul
Pour cette catégorie de mutation sur un seul nucléotide on utilise parfois le terme de mutation génique, mais il y a confusion car celles -ci peuvent avoir lieu sur des parties non codantes, elles ne sont alors pas " géniques ".
2) Les mutations ou remaniements chromosomiques : affectant une portion de chromosome
3) Les mutations caryotypiques affectent le nombre de chromosomes
4) Les mutations d'insertion dues à la transposition d'ADN étranger
Une mutation est dite ponctuelle quand elle ne touche qu'un seul nucléotide dans l'ADN (ou l'ARN pour les virus à ARN)., c'est une variété de mutation nucléotidique
IV.2. Les mutations nucléotidiques peuvent être des substitutions des insertions ou des délétions
Comme mutations nucléotidiques on distingue :
- Le remplacement d'une base par une autre ou substitution ( transitions et transversions ).
- L'ajout d'une ou quelques bases ou insertion
- La perte d'une ou plusieurs bases ou délétion
| Séquence d'origine | Séquente mutante | |
| ATGCCGTA | Substitution | ATGCGGTA |
| ATGCCGTA | Insertion | ATGCCGGTA |
| ATGCCGTA | Délétion | ATGGTA |
- Mutations nucléotidiques ou ponctuelles par substitution
- Mutations faux sens : Cette mutation ponctuelle se traduit par le remplacement d'un nucléotide par un autre. Dans certains cas, cette modification entraîne une modification de l'acide aminé codé, laquelle peut avoir ou non une répercussion sur la fonction de la protéine produite par le gène, dans le cas d'un gène codant, ou sur l'affinité pour un facteur de transcription, dans le cas d'une zone promotrice de l'ADN.
- Mutations non-sens : Le changement d'un nucléotide provoque le remplacement d'un codon spécifiant un acide aminé par un codon-stop. Cela entraîne la production d'une protéine tronquée.
- Mutations silencieuses : Ce sont des mutations qui ne modifient pas la séquence d'une protéine, à cause de la redondance du code génétique (le nouveau triplet code le même acide aminé que le triplet original), ou parce qu'elle touche une région non codante de l'ADN, ou un intron. Mais cette mutation peut néanmoins avoir de graves conséquences sur le phénotype. En effet, le changement d'un seul nucléotide peut changer le site donneur d'épissage, sans pour autant changer la séquence en acides aminés. Cela peut donc se traduire par une délétion entière d'un exon de la séquence peptidique, l'exon n'étant pas reconnu car le site d'épissage a été muté. Une mutation synonyme désigne une mutation silencieuse qui touche un exon, sans changer la séquence de la protéine. Ces mutations feront apparaître des allèles nouveaux d'un gène.
Dès lors couple d'allèles, séries d'allèles......
On parle de mutation de transition lorsqu’il y a substitution d’une base purique à une autre base purique (ou d’une base pyrimidique à une autre base pyrimidique), il y a quatre possibilités.
A ↔ G , T ↔ C, il s'agit bien de 4 possibilités en comptant les " double sens représentés par les doubles flèches.
Au contraire, une mutation de transversion est une mutation causée par le remplacement d’une base purique par une base pyrimidique (ou d’une base pyrimidique par une base purique), il y a huit possibilités.
A ↔ T , A ↔ C et G ↔ T , G ↔ C
- Les transitions sont en moyenne deux fois plus fréquentes que les transversions, alors que si le changement de base s'effectuait au hasard , il devrait y avoir plus de transversions que de transitions , puisque la possibilité de transversions est deux fois plus grande . Ceci est probablement lié , d'une part , aux propriétés des ADN polymérases qui incorporent plus fréquemment une base erronée de même nature lors de la réplication et d'autre part, aux propriétés des mécanismes de correction qui reconnaissent plus facilement une transversion qu'une transition car une transversion entraîne une anomalie de la double hélice plus importante.
De plus , les erreurs au cours de la réplication peuvent être dues à l'incorporation de bases substituées ( tautomères ou isomères ) . Par ailleurs les transitions C → T peuvent se produire par désamination d'une base C méthylée.
- Notion de taux de mutation
Taux de mutation : c’est la probabilité qu’un événement mutationnel se produise en un laps de temps donné. Le taux de mutation est obtenu en mesurant la fréquence de mutant par unité de temps choisi.
Soit le génotype A1/A2.
Probabilité pour qu'un allèle soit A1 = 1/2, idem pour A2.
supposons maintenant un zygote formé par l'union de deux gamètes l'une portant le gène A1 l'autre le gène homologue Ax.
Ce zygote donne un zygote diploïde qui à son tour forme des gamètes .
Certains portent un gène issu, par réplications successives du gène A1 soit °A1.
si pas de mutation, °A1 = A1
Si mutation, l'évènement °A1 = A2 a une probabilité non nulle
P(°A1=A2) = u.1-2
C'est le taux de mutation par cycle de reproduction
IV.3. Les remaniements chromosomiques affectent la structure des chromosomes
Le nombre de chromosomes et leur structure ( caryotype ) caractérisent chaque espèce. Tout changement de structure ou du nombre de chromosomes peut être dommageable pour l'intégrité d'un organisme.
Tout changement de structure des chromosomes par perte, addition ou réarrangement de segments de chromosomes est appelé remaniement chromosomique
Chez les eucaryotes, l'ADN s'individualise en chromosomes lors des divisions cellulaires mitose et méïose. Chez les organismes diploïdes les chromosomes sont regroupés en paires de chromosomes dit homologues. Deux chromosomes non homologues ( conc appartenantà une paire différente ) sont dits hétérologues . Un chromosome est caractérisé par un centromère divisant le chromosome en deux bras dont les extrémités constitue les télomères . Le centromère est essentiel lors des divisions cellulaires car il constitue le point d'attache au fuseau mitotique ou méiotique. Les téloméres sont dépourvus de caractéristiques morphologiques mais possèdent une structure moléculaire particulière dont l'intégrité est requise afin que le comportement du chromosome reste normal. Généralement nous distinguerons deux types de chromosomes selon la position du centromère :
- Les chromosomes métacentriques dont le centromère est situé au milieu du chromosome. Le centromère divise le chromosome en deux bras égaux. Un chromosome submétacentrique présente un centromère à proximité de son milieu.
- les chromosomes acrocentriques dont le centromère est proche d'une extrémité, le bras court est dans ce cas de très petite taille , certains auteurs distinguent des chromosomes télocentriques pour lesquels le centromère est en position terminale.
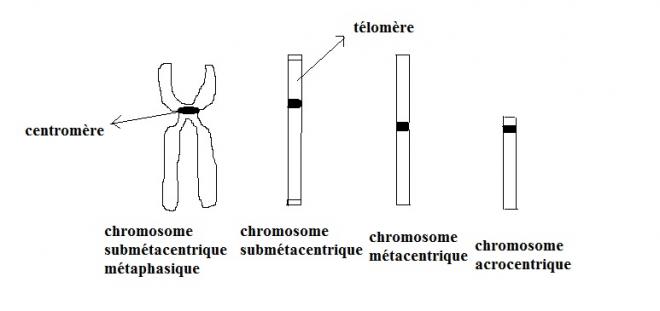
Par convention , lorsque les deux bras sont de longueurs inégales, le bras le plus long désigné par la lettre q, est le bras inférieur et le bras le plus court désigné par la lettre p est le bras supérieur.
Dans ce chapitre, nous simplifierons des chromosomes sous forme de "bâtonnets" .
Les remaniements chromosomiques peuvent être divisés en quatre catégories : délétions, duplications, inversions et translocations qui comprennent les translocations réciproques, les insertions et les fusions. Ces mutations se produisent lors des divisions cellulaires lors des étapes les plus critiques que sont l'appariemment des chromosomes homologues et la ségrégation régulière des chromosomes.
Les anomalies de structure sont le résultat de cassures des chromosomes durant la méiose. Une délétion, une duplication ou la formation d'un isochromosome se traduiront par un phénotype anormal, tandis que l'insertion, l'inversion, ainsi que la translocation peuvent être équilibrées. Ceci signifie que les porteurs de ces anomalies de structures sont phénotypiquement sains, car la totalité du matériel génétique est présente.
Les DELETIONS
Elles se traduisent par la perte d'un segment de chromosome ne portant pas de centromère. En effet, le segment de chromosome délété, dit acentrique ( sans centromère ) ne pourra plus se déplacer et sera perdu. Les délétions ne mettent en jeu qu'un chromosome et peuvent être de deux types :
- Lorsque la délétion se situe en position terminale , on parle de déficience . Dans ce cas, il n'y a qu'une cassure chromosomique et le segment ne comprenant pas de centromère est perdu.
- Lorsqu'il y a deux cassures chromosomiques, on parle de délétion segmentaire ou intersticielle. Il y a perte du fragment situé entre les deux cassures.
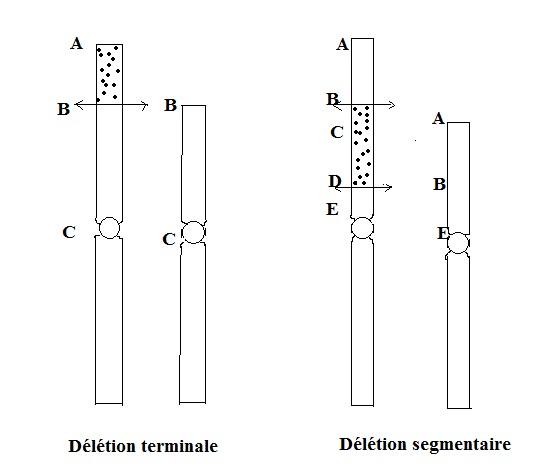 Une délétion peut avoir lieu dans n'importe quel chromosome et peut atteindre n'importe quelle grandeur. Les conséquences d'une délétion dépendent de sa longueur et des gènes qui sont amputés.
Une délétion peut avoir lieu dans n'importe quel chromosome et peut atteindre n'importe quelle grandeur. Les conséquences d'une délétion dépendent de sa longueur et des gènes qui sont amputés.
Une délétion partielle du bras court (p) du chromosome 5 est responsable de la maladie du Cri du chat
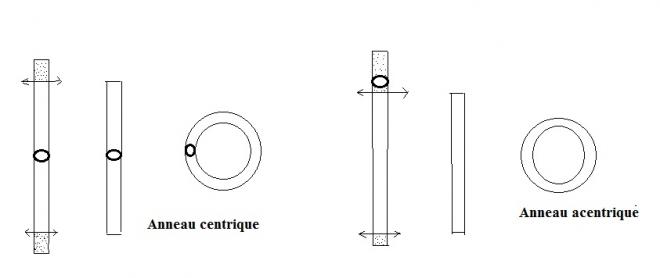
Parfois les bouts des chromosomes (télomères) peuvent se casser et être perdus. Dans ce cas, un chromosome en anneau résultant de la fusion des 2 bouts peut se former. Si aucune information génétique essentielle est perdue, ces réarrangements sont équilibrés et phénotypiquement invisibles. Des problèmes surviennent uniquement lors de la formation des gamètes.
La formation d'un chromosome en anneau résulte de l'occurence d'une double délétion intervenant sur les deux extrémités du même chromosome qui sont ensuite " recollées" . Si le centromère est présent, sinon il est dit acentrique et est rapidement perdu au cours des divisions cellulaires.
Les INVERSIONS
Les inversions correspondent à un retournement à 180° d'un segment de chromosome . Dans ce cas, il y a également une double cassure chromosomique , mais non accompagnée d'une " perte de matériel chromosomique, car le segment est "recollé" après rotation. Les inversions ne concernent qu'un seul chromosome. Selon que le centromère est impliqué ou non dans l'inversion, on distingue :
- Les inversions paracentriques sil le centromère n'est pas inclu dans l'inversion.
- Les inversions péricentriques si le centromère est inclu dans l'inversion, dans ce cas une inversion peut transformer un chromosome un chromosome acrocentrique en un chromosome métacentrique et inversément.


.
Date de dernière mise à jour : 05/07/2021
Ajouter un commentaire


