
10. Théories contemporaines de l'évolution
2.2 Hypothèses sur les types d'architectures des animaux fin Précambrien
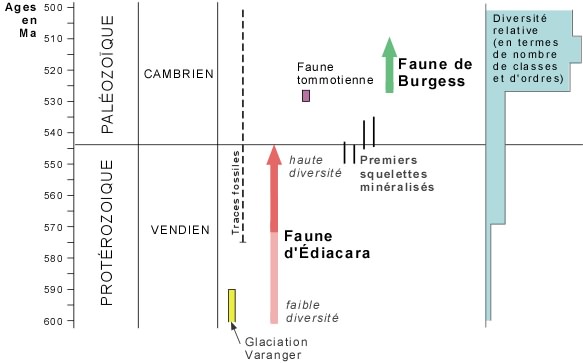
Dès 700 millions d'années, on trouve des terriers des traces d'animaux. Donc, à cette époque déjà, sont apparues des communautés d'animaux fouisseurs qui vivaient de sédiments marins ; De plus, la diversité des terriers indique que ces animaux s'étaient adaptés dès cette époque à des modes de vie différents ce qui implique des organisations structurales différentes.
- Vers existant actuellement et apparus au Cambrien :
- Vers au corps très segmenté : parois transversales ( dissépiments ) divisent le coelome en un grand nombre de cavités. Architecture bien adaptée à un fouissement efficace.
Pourquoi ?
Activité musculaire associée au fouissement affecte seulement les segments situés au voisinage immédiat des contractions renforçant ainsi l'efficacité de l'opération
............ Vers de terre.
- Vers au corps peu segmenté : deux ou trois compartiments, chacun fonctionnant différemment
............. Phoronidiens ( E. des Lophophoriens ).
- Temps forts : - 700 M d'années, apparition du coelome ; - 700 à - 570 M : développement du coelome chez les Vers fouisseurs ; - 570 : apparition des animaux à squelette résistant.
Il y a une diversification intense des Vers au sein de leur zone d'adaptation.
- Les animaux à squelette :
1° - 570 M, apparition des premiers organismes à squelette persistant mais les sédiments ne contiennent que de minuscules éléments squelettiques ; aucun rapport avec les groupes vivant actuellement.
Tout montre qu'il y a eu une importante diversification des phyla au Cambrien.
2° - Qualité du squelette : le squelette fait le groupe
Explication : les brachiopodes n'ont pas pris naissance à partir de brachiopodes nus, ils ont pris naissance d'ancêtres mous, non Brachiopodes et par un chemin évolutif , ils ont développé un squelette minéralisé dont les caractéristiques cruciales permettent de définir l'état Brachiopode.
Le squelette = marque de fabrique
Ces formes primitives ayant possédé des squelettes, avaient un point commun : vivre à la surface du fond marin ( épifaune ).
Le squelette remplace le terrier comme moyen de protection, donc, il s'agit d'une adaptation à la vie sur les fonds marins.
La variété des plans de construction squelettique dans différents groupes d'Invertébrés est le résultat de diverses adaptations aux problèmes de la vie benthique.
Exemples :
1) Des vers fouisseurs acquièrent un squelette articulé externe . Avantage ?..... C'est un avantage, celà facilite le déplacement à la surface du fond par rapport au péristaltisme. Descendant actuels : Phylum des Arthropodes .....
2) Brachiopodes actuels ? Ancêtres comparables aux Phoronidiens actuels :
- Phoronidiens dans un tube chitineux ( endofaune ).
- Phorodiniens de surface ( tube chitineux fixé aux rochers ).
- Développement de la coquille pour soutenir les viscères et protéger.
SCENARIO SUR APPARITION ET DIVERSIFICATION DES GROUPES.
Apparition de la vie : - 3,5 milliards d'années ( Bactéries, Cyanophycées )
Apparition des Eucaryotes : - 1,5 milliards d'années
Apparition des premiers êtres pluricellulaires : - 700 millions d'années : méduses, vers plats. Epanouissement des vers ........... Communautés d'animaux fouisseurs.
Apparition des premières communautés de l'épifaune : - 570 millions d'années.
" Le squelette" s'accompagne de très nombreux changements anatomiques des parties molles, changements qui ont persisté jusqu'aujourd'hui.
Remarque : Il est possible que tous les grands groupes d'animaux soient apparus à cette époque charnière PC/C.
On considère que sur X phyla qui ont pris naissance à cette époque géologique X/2 on survécu.
Ce scénario a suscité, cependant, l'existence de nombreux points en suspens.
Trois problèmes se posaient :
- 1) Comment les grands groupes d'animaux ont pu se différencier aussi rapidement , au cours d'un laps de temps de quelques dizaines de millions d'années ?
- 2) Pourquoi une vingtaine de phyla, pourquoi pas 1 ou plusieurs centaines ?
- 3) Pourquoi tous les phyla, ou presque prirent-ils naissance précisément pendant le laps de temps relativement court PC/C ? Quel est l'évènement clef ayant permis la diversification ?
2.3. Essais de réponse à ces problèmes étroitement imbriqués.
a) Aussi rapidement : l'apparition brutale s'opposait à la conception classique évolutionniste ( gradualiste ) ....... micro-évolutionniste : somme de petits changements ( pour une espèce nouvelle, plusieurs millions d'années, pour un phylum , ... de l'ordre du milliard d'années ...??!!!. Actuellement c'est plus acceptable suite à l'explication fournie par Gould et Eldredge par l'alternance de stases et de ponctuations...
A l'époque, l'on a mis en avant l'hypothèse de changements dans la structure des gènes. en effet, il fallait trouver un autre type de mécanisme évolutif.
Peut-être un changement dans le mode d'expression des gènes ?
Explications et images :
Les gènes commandent les synthèses de protéines qui sont à l'image des briques, du bois, du ciment .... des matériaux pour des constructions ......... Avec ces matériaux, on peut construire une maison, un building, une usine... La différence n'est pas dans la nature des matériaux mais dans
- les quantités relatives de chacun d'eux
- la manière dont ils sont répartis, utilisés selon des plans de construction...différents .
De même : différence entre deux Vertébrés ? différence entre tissus musculaire et tissus nerveux ?
Différences dans les constituants ?... Surtout différence dans l'organisation et distribution de ces constituants.
L’épigénome est l'état épigénétique de la cellule. À l'image des cellules embryonnaires qui peuvent avoir plusieurs fonctions finales, un unique génome peut être modifié de multiples manières pour donner des épigénomes différents. Il est actuellement conjecturé par un grand nombre de chercheurs en épigénétique qu'un code épigénétique existe dans chaque cellule eucaryote - par analogie au code génétique. À l'extrême, ce code épigénétique représente le type et la position de chaque molécule de la cellule.
Mais encore : le texte suivant est à méditer :
On a longtemps cru qu’il fallait un très grand nombre de mutations élémentaires, lentement accumulées dans le capital génétique, pour obtenir une autre espèce, en particulier pour obtenir un nouveau plan d’organisation. Cependant on a trouvé que ce n’était pas seulement un caractère qui est concerné par un gène mais le plan d’organisation lui-même. Donc certains gènes permettent d’ordonnancer la confection d’un individu, étape par étape. Il s’agit là de la découverte des gènes homéotiques et de l’homéoboite. C’est un mode de fonctionnement qui règle le développement embryonnaire de l’individu et que l’on va retrouver d’abord chez la drosophile puis chez les insectes, enfin chez les mammifères. En effet, que ce soit chez la drosophile (avec les gènes HOM) ou chez la souris (avec les gènes HOX), on trouve une même base, l’homéoboite, qui doit provenir d’une origine commune. C’est au point que l’on a décidé de ne plus utiliser les termes différents Hom et Hox mais le même terme Hox pour désigner l’homéoboite. Un tel gène homéotique donne un ordre des opérations pour la fabrication d’un individu. Dans l’histoire des gènes du développement, tout a commencé en 1894. On a découvert des transformations homéotiques de la mouche drosophile. Le terme d’homéotique vient des mutants homéotiques que l’on a trouvé dans la mouche drosophile avec, par exemple, des pattes à la place des antennes. Homéotique signifie que ce sont des transformations d’une partie du corps à l’image d’une autre partie, d’où la notion d’homologie qui a donné le terme homéotique. Une drosophile mutante peut recevoir 4 ailes alors que normalement elle n’en possède que deux. On peut voir des antennes à la place des pattes, des yeux à la place des ailes. Cela ne se fait pas au hasard mais par homologie : par exemple on ne peut pas échanger des pattes poussant à la place d’antennes et non des ailes à la place des pattes. Des structures d’apparence très différentes peuvent s’échanger si elles interviennent au même niveau dans le développement de l’individu. Ce qui détermine le moment de formation de l’organe, ce sont les gènes du développement. Ce sont les gènes Hox, dits encore gènes architectes, qui permettent la réalisation du plan d’organisation et donnent au développement ses étapes, son sens. On retrouve les Hox non seulement chez la souris, la drosophile ou l’homme mais chez tous les pluricellulaires. Il y a correspondance entre chaque gène et une zone du corps. Ces gènes s’expriment successivement suivant leur place dans le cordon ADN correspondent non seulement à l’ordre des zones du corps mais aussi à l’ordre temporel de fabrication de ces zones. Il y a une durée d’expression de chaque gène. Il y a un début et une fin de l’intervention de chaque gène qui est déterminée par l’action d’un gène régulateur. Celui-ci peut activer ou inhiber le gène Hox. Il reste un problème à résoudre qui est une question d’auto-organisation des systèmes dissipatifs, étudiée comme telle par Alan Turing et Brian Goodwin. Les gènes Hox ne se contentent pas de fabriquer un membre ou un système comme le système urogénital, ils précisent dans quelle direction le membre ou l’organisme doit se développer. C’est Wolpert qui en a donné l’interprétation en développant une idée de Turing dont on a parlé précédemment. C’est un phénomène de disposition spatiale par rapport aux voisins proches, au sein d’un tissu en croissance. C’est ce que Wolpert appelle l’information de position. En effet, ce qui permet de fabriquer un individu c’est que les cellules ne se contentent pas de se multiplier ou de se diversifier : elles connaissent leur position au sein d’un tissu en croissance. Mais comment font-elles pour le savoir, pour organiser l’espace. Et comment les différents organes de l’individu se retrouvent bien à leur place par rapport à l’avant/arrière du corps et par rapport à la symétrie dos/face ? Ce sont les gènes Hox qui assignent une identité spatiale aux cellules embryonnaires le long des différents axes du corps. En coloriant ces zones par des couleurs, bleu, blanche ou rouge, Wolpert arrive à la fameuse image du drapeau français. La forme réalisée est un drapeau tricolore mais ce n’est pas tout à fait le drapeau national bien connu car celui des Hox est doué d’auto-régulation avec des effets de seuil. Au fur et à mesure de la différenciation cellulaire, les cellules vont savoir se grouper en fonction de leur type grâce à des messages inter-cellulaires et cette transformation est irréversible. Ce phénomène est non seulement spatial mais temporel. En fonction du temps passé à la réalisation de telle ou telle tâche du programme, la tâche suivante sera ou non enclenchée, avancée ou retardée. Donc les gènes Hox expriment un programme, non prévu d’avance, d’auto-organisation spatio-temporel suivant sur la manière dont le processus se réalise à chaque fois. Le fait qu’il en résulte un individu semblable ne provient pas d’un modèle tout fait imité mais du fait que les gènes Hox agissent dans l’ordre de leur position dans l’ADN. Les gènes Hox permettent de concevoir le mode de régulation du développement, c’est-à-dire le lien entre deux niveaux fondamentaux du vivant : le programme génétique et la morphologie des individus. De petites mutations sur les gènes Hox peuvent causer des modifications importantes des horloges du développement, entraînant des changements morphologiques radicaux. Il n’y a pas de liaison linéaire entre génétique et morphologie. Finalement les gènes Hox ne construisent pas eux-mêmes l’organisme mais donnent aux cellules les propriétés qui leur permettent de le construire. Les gènes du développement ne sont pas un simple programme duplicatif. Un gène va bien lancer la fabrication d’un oeil, mais ce gène est seulement un gène de pilotage de milliers d’autres gènes de structure. Le fonctionnement des gènes du développement est hiérarchisé et il l’est à trois degrés : d’abord les gènes de structure, puis les gènes architectes et enfin les gènes régulateurs supérieurs. Un gène régulateur pilote de milliers de gènes de structure. Les gènes architectes sont un petit nombre de gènes qui envoient des ordres à des gènes dits ouvriers et commandent ainsi aux diverses opérations visant à lancer des développements du corps, de la tête, des membres. Le pilotage, l’activation et l’arrêt des gènes architecte est réalisé par des gènes de régulation. En somme, il y a celui qui bâtit, celui qui donne le plan de la maison et répartit les tâches, et enfin celui qui ordonne d’agir ou d’arrêter le travail. Le mot clef du développement est donc celui d’organisation. C’est l’attribution de fonctions aux différents modules des gènes du développement qui change sous l’action des gènes régulateurs, les homéogènes. Certains gènes peuvent être activés ou inhibés, ce qui explique aussi qu’un même ADN de la cellule originelle de l’individu puisse servir à produire des cellules d’un grand nombre de types différents. François Jacob, dans sa conférence, déjà citée déclarait : « chez la mouche ont été mise en évidence les gènes qui assurent dans l’oeuf la mise en place des axes du futur embryon, puis qui découpent le corps de l’embryon en segments, puis ceux qui déterminent le destin et la forme de chacun de ces segments. A la stupéfaction générale, ces mêmes gènes ont été retrouvés chez tous les animaux examinés ; coup sur coup grenouille, ver, souris, homme. Qui eut dit, il y a encore quinze ans que les gènes qui mettent en place le plan d’un être humain sont les mêmes que ceux fonctionnant chez une mouche ou chez un ver ? » Gould écrit dans « Quand les poules auront des dents » : « Nous avons trois niveaux hiérarchiques de contrôle : les gènes de structure qui construisent les différentes parties de chaque segment, les gènes régulateurs homéotiques qui déclenchent la mise en service des batteries de gènes de structure, et les gènes régulateurs supérieurs qui déclenchent la mise en service des gènes homéotiques au moment et à l’endroit voulu. » Il n’y a pas de gène qui soit relié linéairement à un pied. On a greffé un gène de patte de souris chez un poulet, et c’est une patte de poulet qui a poussé. Le gène de patte dit seulement : « tu allonges un membre ». Ce n’est pas ce gène qui dit ce qui va pousser car il se contente de transmettre l’ordre et c’est avec le plan de la maison que cet ordre est interprété. Du coup le plan de l’oeil est, comme le plan d’une patte, le même pour tous ces animaux. La capacité des gènes de se définir temporellement fait d’eux des horloges du développement. Du coup, on ne distingue plus ce qu’on croyait être deux types de mutations : les mutations homéotiques et les mutations hétérochroniques, c’est-à-dire les altérations de la chronologie des horloges du développement. En termes plus simples, cela signifie que l’évolution des espèces serait une modification de rythme d’horloge. Le premier exemple, pr, est un travail très ancien, celui de D’Arcy Thompson en 1917, dans son ouvrage « Formes et croissance », où il indique notamment comment de simples modifications des vitesses du développement peuvent modifier la morphologie finale. Le deuxième exemple est l’étude de David Raup, sur la base des idées de D’Arcy Thompson, qui a montré qu’on peut transformer la coquille d’un escargot en palourde simplement en faisant varier trois paramètres C’est le lien entre les gènes architectes et les horloges du développement que nous allons examiner maintenant. Nous sommes en pleine actualité de la recherche puisque ce sont plusieurs découvertes très récentes. Ces découvertes changent fondamentalement la situation dans le débat sur la théorie de l’évolution, à mon avis en faveur de la thèse de Stephen Jay Gould et Jean Chaline. Ce sont trois découvertes fondamentales :
premièrement on sait maintenant comment l’embryogenèse est pilotée par des gènes, appelés les gènes architectes. Leur fonctionnement est une véritable cybernétique fondée sur des rétroactions sur un mode hiérarchique. Cela ouvre la possibilité qu’une ou deux petites modifications sur un gène qui pilote des milliers de gènes entraînent un changement de toute la morphologie de l’être vivant.
deuxièmement la découverte la plus étonnante est que les gènes et les protéines sont quasiment les mêmes pour un insecte et un mammifère. Ce qui change d’une espèce pluricellulaire à une autre, n’est pas tant les protéines ou les gènes, mais leur temps et leur vitesse d’action, leur organisation et leur rythmologie. Notamment les gènes architectes sont les mêmes pour tous ces êtres vivants. Ils sont seulement un peu bricolés par l’évolution et finissent par remplir des fonctions très diverses. Nous trouvons donc le hasard et le bricolage au sein de la génétique qu’on croyait être le domaine de l’ordre fixe. La génétique obéit à une rythmologie d’horloges capables d’interactions. Ce ne sont donc pas des horloges périodiques.
troisièmement, il existe un mécanisme biochimique, permettant au milieu extérieur (en cas de changement climatique important et brutal, d’agression chimique ou par rayonnement) de provoquer chez l’être vivant un stress, qui inhibe le mécanisme de conservation de l’espèce. C’est la découverte des biologistes Rutherford et Lindquist concernant la mouche drosophile. Ces deux biologistes américaines viennent de montrer qu’un stress intense peut provoquer chez cet animal des mutations héréditaires. Ces mutations entraînent des anomalies impressionnantes concernant la morphologie de l’animal, par exemple la dimension des pattes ou des ailes ou la pilosité. Publiée en décembre 1998, leur étude montre qu’un effet de choc produit de nouvelles espèces. Un choc qui casse l’ordre puis produit un nouvel ordre, voilà qui fait penser au chaos déterministe. Ces deux chercheuses ont trouvé la cause de ces mutations de la mouche du vinaigre, la drosophile : un gène qui code une protéine appelée Hsp90. Comme l’indique le nom Hsp, « heat shock protein » c’est-à-dire protéine de choc thermique, cette protéine sert à protéger d’autres protéines des chocs thermiques extérieurs. Les chercheuses ont montré qu’en période de stress intense, la protection se relâche, favorisant des possibilités de biodiversité qui étaient inhibées jusque-là. En effet, Hsp90 aurait comme rôle, en temps normal, de s’assurer que les autres horloges ne sont pas décalées dans le temps. Du coup, il inhibe le potentiel de difformité que permet le fonctionnement des gènes et des protéines. Hsp agit de sorte que le développement mette de côté les petites modifications des protéines. Or les protéines qui sont ainsi protégées contre toute erreur de route sont justement celles liées à des processus de multiplication et de développement cellulaire. Avec la mutation du gène de protection puis celle de la protéine Hsp90, des mutations deviennent à nouveau possibles et peuvent se produire sans contrôle. Il n’y a plus qu’à imaginer qu’elles portent sur des cellules embryonnaires pour disposer pour la première fois d’un mécanisme génétique permettant de comprendre comment apparaît une nouvelle espèce. Les drosophiles mutantes qui sont parvenues à atteindre l’âge adulte ont été pour la plupart des nouvelles mouches viables. Non seulement la mutation est héréditaire mais elle se conserve en cas de croisement entre deux animaux ayant l’un Hsp mutant et l’autre un Hsp normal. Ce processus suggère que de nombreuses espèces pourraient acquérir de la nouveauté dans un environnement qui changerait brutalement, par exemple sur le plan climatique. C’est une interprétation intéressante des bouffées de variabilité que l’on a constaté sans les expliquer jusque là. On a constaté en effet que de nombreuses espèces subissent en même temps des variations. Les protéines chaperon, type Hsp, qui inhibent les variations des autres protéines et constituent un mécanisme de type homéostasie ont été trouvées également chez les mammifères, les végétaux et les insectes. Quant à la protéine Hsp90, elle est la même pour toutes sortes d’êtres vivants, de la levure aux mammifères. C’est donc une hypothèse très tentante. C’est même la première fois qu’un mécanisme moléculaire postule à expliquer, en un tout petit nombre de mutations, des changements visibles et héréditaires de la morphologie. Or, justement, une découverte a été faite au cours de l’année 1998 sur le mécanisme des mutations héréditaires : Chaline a étudié un poisson du Pacifique, près de la côte du Mexique, l’axolotl, dont l’évolution est inachevée et qui, normalement, reste à l’état larvaire, intermédiaire entre le poisson et l’animal terrestre. Il a démontré qu’un des produits biologiques d’un cycle de rétroaction, la thyroxine, est déterminant dans l’évolution de l’espèce avec une bifurcation, soit vers le poisson, soit vers l’animal terrestre. En effet, l’excès ou la carence de ce produit enclenche l’évolution de l’espèce. La concentration de ce produit biologique modifie l’équilibre rythmique des cycles de rétroaction biochimiques lesquels règlent la formation de l’animal. Or modifier l’un de ces cycles, c’est choisir une bifurcation dans un sens ou dans l’autre, soit vers le poisson soit vers l’animal terrestre. Et c’est enclencher un processus qui ne concerne pas que l’individu mais toute sa descendance car il met en jeu les gènes concernés. On dispose donc d’un exemple de retardement de l’horloge biologique et on sait comment il est relié à la génétique. On sait également qu’une action chimique permet d’intervenir sur le cycle qui commande cette horloge, en inoculant ce produit. De ces diverses recherches il découle :
premièrement, que c’est au niveau de l’embryon qu’une espèce peut changer et que la comparaison des espèces ne doit pas partir des comparaisons d’adultes comme le faisait la phylogenèse, c’est-à-dire la recherche des lignées d’espèces (les phylums). Ce qui rapproche les différents animaux qui se succèdent dans l’évolution, et permet de concevoir le passage d’une forme à une autre, n’est pas l’état adulte mais embryonnaire. C’est un renversement complet de perspective dans la relation entre phylogenèse et ontogenèse (l’ontogenèse étant la succession des phases par lesquelles s’élabore un individu).
deuxièmement, qu’un changement morphologique d’espèce peut tout à fait se produire brutalement, avec une ou deux modifications sur l’ADN et non des millions, comme le supposait le néo-darwinisme. Cela signifie que des espèces apparaissent brutalement et non sur des temps très longs comme le défend cette thèse. Il peut suffire d’un tout petit nombre de mutations sur des gênes, les gênes du développement, pour entraîner un changement dans l’horloge qui commande la fabrication embryonnaire de l’individu. ![]() troisièmement, ce sont des altérations du rythme et des vitesses du développement de l’individu qui changent d’une espèce à une autre. Cette propriété est due au type chaotique de ces horloges produites par l’auto-organisation de réactions biochimiques rétroactives.
troisièmement, ce sont des altérations du rythme et des vitesses du développement de l’individu qui changent d’une espèce à une autre. Cette propriété est due au type chaotique de ces horloges produites par l’auto-organisation de réactions biochimiques rétroactives.
jeudi 26 juin 2008, par
Bref,
L'hypothèse de base était que l'élaboration d'un nouvel organisme aurait impliqué un changement de l'appareil régulateur plutôt que des changements au niveau des gènes de structure. toutes les cellules d'un organisme portent le même génotype, mais partant de l'oeuf fécondé et durant tout le développement embryonnaire, un programme génétique détermine :
- quand et comment les lignées cellulaires différenciées apparaissent.
- quand et comment les différentes protéines seront synthétisées et leur quantité ( qualité et quantité variant dans l'espace et dans le temps ).
Les gènes régulateurs dirigent ce programme.
toute modification dans un gène régulateur entraîne donc des modifications chez plusieurs gènes de structure ( voir ou revoir la théorie sur l'opéron lactose, l'opéron tryptophane ), ce qui se marquera par des modifications/variations anatomiques ; anatomie qui dépend très fortement de la "séquence temporelle" du développement .
De tels changements brutaux ne correspondent pas à la philosophie graduelle du Néodarwinisme. Dès lors, il faut trouver un autre mode d'explication pour solutionner l'apparition brutale de grand groupes d'animaux : Ces changements se feraient au niveau de la séquence temporelle, ce que l'on appelle néoténie ou hétérochronie.
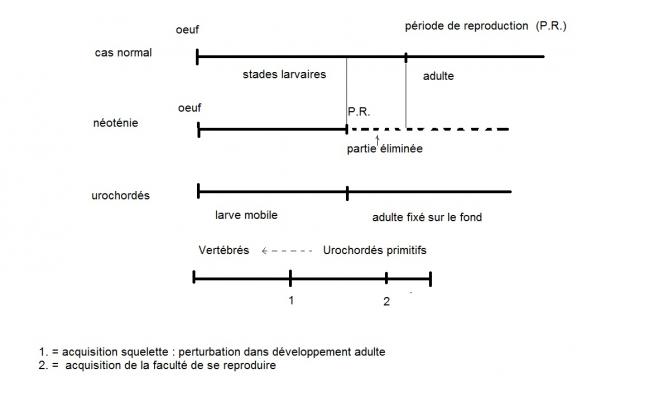 Résultat : adulte d'une forme nouvelle, individu libre et nageur .
Résultat : adulte d'une forme nouvelle, individu libre et nageur .
Le squelette va permettre d'exploiter cette opportunité locomotrice.
CONCLUSION : HETEROCHRONIE =RESSORT MAJEUR du mécanisme évolutif de la MACRO-EVOLUTION.
b) Elaboration d'un arbre généalogique ramifié
- Macro évolution : changements brutaux dans la morphologie ( = percées évolutives ) , naissance de phyla
- Micro-évolution : accumulation lente de changements mineurs à partir d'un stock fondamental, naissance des espèces. Pas de percées évolutives.
Un phylum nouveau subira une maturation , s'il s'adapte aux changements du milieu extérieur et les espèces apparaîtront.
Donc si pas de macro-évolution, toutes les espèces animales auraient le même plan de base et toutes seraient à classer dans le même phylum
La micro-évolution raffine les caractères d'une population en fonction de l'environnement auquel elle est confrontée.
Le troisième et dernier problème : pourquoi jadis beaucoup de percées évolutives et rien actuellement ?
Il est prouvé que le nombre d'espèces que l'environnement peut supporter est limité .
Au Cambrien, contrairement au Quaternaire, les habitats potentiels vacants sont nombreux. Les percées évolutives avaient une probabilité élevée de réussir.
CONCLUSION :
1° Le nombre de phyla est déterminé par la compétition entre deux facteurs
- La saturation des habitats par les descendants modifiés d'autres phyla.
- La différenciation de phyla entièrement nouveaux.
2° Du Cambrien au Quaternaire, les types d'organisations faibles s'accrurent aussi.
Quels sont les déclics qui ont fait fonctionner ces mécanismes évolutifs.
Spéculations :
- stimulation du rythme évolutif par changements climatiques ?
- stabilisation de la productivité des écosystèmes par les populations végétales ?
- accroissement de la concentration en O2 jusqu'au niveau requis pour subvenir au métabolisme des formes animales de plus en plus actives ?
- explications possibles à chercher dans la structure des animaux mêmes ?
IV.4 La non-disjonction méiotique ou mitotique entraîne un changement du nombre de chromosomes
On appelle ploïdie, le nombre de copies des différents chromosomes chromosomes du génome nucléaire des eucaryotes . L'euploïdie désigne une cellule ou un individu dont tous les jeux de chromosomes sont complets. L'aneuploïdie désigne un écart à cet état ( jeux chromosomiques incomplets).
La ploïdie chez les eucaryotes est en général égale 1 ( organismes haploïdes ) ou 2 ( organismes diploïdes ). Si un organisme se reproduit de façon sexuée , plusieurs niveaux de ploïdie coexistent chez ces organismes. Il existe différents types de changement du nombre de chromosomes, résumés dans le tableau suivant.
| Types de mutation ( caryotype ) | Nombre de chromosomes | ||
| Aneuploïdie |
par défaut -nullisomie -monosomie Par excès -trisomie -tétrasomie -disomie |
2n-a 2n-2 2n-1 2n+a 2n+1 2n+2 n+1 |
|
| Euploïdie |
-diploïdie -triploïdie -polyploïdie |
2n 3n a(2n) |
Tout changement du nombre de chromosomes sans modification de la structure est appelé mutation génomique ou caryotypique.
L'aneuploïdie désigne un changement du nombre de chromosomes dû à une perte ( aneuploîdie par défaut : 2n-a) ou à un gain ( aneuploïdie par excès : 2n+a ), d'un ou de plusieurs chromosomes mais ne concerne pas le lot entier de chromosomes.
Date de dernière mise à jour : 05/07/2021
Ajouter un commentaire