
Bio moléculaire 4
IV REPLICATION DE L'ADN
Lors de la division cellulaire, quand une cellule-mère donne deux cellules-filles, il est essentiel que l’ADN présent dans les cellules-filles soit la copie identique de l’ADN présent dans la cellule-mère. Cette copie de l’ADN est indispensable à réaliser avant la mitose (ou division cellulaire) on parle de réplication de l’ADN. Préalablement à toute division cellulaire, la quantité d’ADN est multipliée par deux. Les mécanismes de réplication conditionnent donc le déroulement de la division cellulaire.

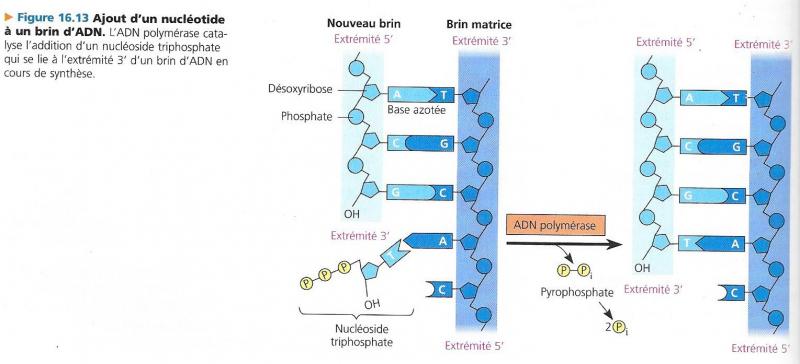
La croissance des brins d’acides nucléiques se fait toujours par leur extrémité 3’-OH terminale.
• La condensation se fait à partir d’un substrat « activé » : un des nucléosides triphosphates. La rupture d’une liaison riche en énergie fournira l’énergie nécessaire à la condensation. Le nucléoside monophosphate restant sera estérifié par une fonction acide de son phosphate sur la fonction alcool libre du carbone 3’ du ribose qui constitue l’extrémité de l’acide nucléique.
• Inversement, en ajoutant une molécule d’eau sur cette liaison ester, on provoquera une réaction d’hydrolyse qui détachera le dernier nucléotide et libérera le carbone 3’ du nucléotide précédent.
L’hybridation des nucléotides complémentaires peut se faire sur toute la longueur d’un brin d’acide nucléique : par des liaisons hydrogène, on associe systématiquement les C avec des G et les G avec des C, les A avec des T et les T avec des A.
• En réunissant tous les nucléotides ainsi associés par des liaisons phosphodiester on constitue une séquence complémentaire de la séquence originale. Ces deux séquences sont obligatoirement orientées dans des sens opposés : on dit qu’elle sont antiparallèles.
• De cette façon, grâce à des enzymes spécifiques, une séquence d’acide nucléique dite « originale » est capable de diriger la synthèse d’une séquence d’acide nucléique complémentaire.
• Dans un second temps, cette séquence complémentaire isolée, sera elle aussi capable de diriger la synthèse de la séquence originale dont elle est le complément.
IV.5.1 La réplication chez les procaryotes
IV.5.1.1 Caractéristiques fondamentales
- La réplication de l’ADN est semi-conservative (Meselson et Stahl, 1958).
A chaque réplication, les deux brins d’ADN parental se séparent, chacun de ces deux brins sert de matrice pour la synthèse d’un brin complémentaire.
Chaque molécule de l’ADN fille portera un brin de l’ADN mère.
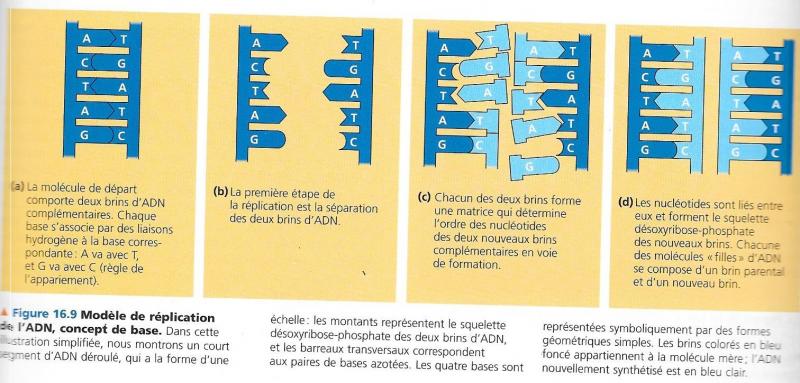
IV.5.1.2 Eléments nécessaires pour la réplication.
La réplication de l’ADN nécessite:
- Tout d’abord, une matrice d’ADN constituée par un brin parental.
- La présence de nucléotides propres à l’ADN, c’est-à-dire contenant du 2’-désoxyribose, des bases A, T, G et C et sous forme de nucléosides triphosphates porteurs d’énergie pour les réactions d’assemblage: dATP, dTTP, dCTP et dGTP (on écrit souvent pour les dénommer dNTP).
- La présence de nombreux enzymes pour assurer la séparation des deux brins et accrocher les nucléotides les uns aux autres
- La présence de certains ions (cations bivalents: Mg2+, ce cation est indispensable pour la réplication de l’ADN).
IV.5.1.3 Mécanisme de la réplication
1. Caractéristiques fondamentales de la réplication
La réplication se fait :
- dans le sens 5’ 3’
- de façon complémentaire
- de manière antiparallèle.
2. Ouverture de la double chaîne
L’enzyme responsable de l’ouverture de la double chaîne est appelée hélicase, après une certaine ouverture, celle-ci est bloquée , par l’existence de contraintes.
Une première étape pour la réplication du DNA est constituée par ces changements de forme topologique.
Ces contraintes peuvent être supprimées par des enzymes
- Girase qui crée des supertours négatifs ( Topoisomérase II)
- Topoisomérase I.
Le DNA d’un chromosome qui peut mesurer jusqu'à 1 m déroulé en totalité, change sa forme super enroulée pour passer à une forme moins enroulée, dans laquelle il est accessible aux autres enzymes .
Deux types de topoisomérases
• la topoisomérase de type I coupe l'un des deux brins du DNA, le DNA peut se dérouler en une forme B, puis survient une nouvelle ligature du DNA.
• la topoisomérase de type II agit en coupant les deux brins du DNA (extrémité 3’ du DNA), et joue un rôle dans le relâchement des boucles pour la transcription semi réplicative.
C’est James Wang qui dans les années 70 découvrit une nouvelle classe d’enzyme, les topoisomérases, qui sont capables de changer la topologie de la molécule d’ADN en effectuant une coupure temporaire dans la molécule pour y faire passer soit un brin, soit les deux brins de la double hélice. Ce faisant, ces enzymes permettent de relâcher les contraintes de torsion sur une molécule ou de désenchevêtrer deux molécules entortillées.
La découverte des topoisomérases a permis de résoudre le problème des nœuds dans les molécules d’ADN. Cependant, leur fonctionnement a soulevé d’autres questions : comment des enzymes mesurant quelques nanomètres peuvent-elles relâcher jusqu’au dernier tour d’entortillement des molécules qui s’étendent sur des distances jusqu’à un million de fois plus grandes ? Comment ces enzymes déterminent de quel côté de la brèche il faut transférer une molécule afin de défaire un nœud et non pas, au contraire, en créer de nouveaux ? Les réponses à ces questions nous manquent encore à l’heure actuelle. Certains de nos résultats suggèrent que ces enzymes reconnaissent l’angle formé par les molécules lors de leur croisement. En particulier, un type de topoisomérase que l’on trouve chez les bactéries se comporte très différemment selon que cet angle est positif et négatif. Cependant, les vérifications expérimentales de ces hypothèses sont particulièrement délicates à réaliser dans des expériences de biologie classique, c’est-à-dire faites en tube à essai. En effet, dans ce contexte, comme les molécules d’ADN sont soumises à l’agitation thermique, l’angle qu’elles adoptent lors de leur croisement est largement aléatoire.
Exemple action de la topoisomérase II

Article wikipedia
Toutes les topoïsomérases identifiées utilisent un résidu tyrosine pour cliver la liaison phosphodiester de l'ADN. En fonction de leur mode de coupure, elles sont classées en deux groupes : les ADN topoïsomérases de type I (monomériques et, hormis la reverse gyrase, ATP- indépendantes), qui coupent un seul des deux brins de la double hélice de l'ADN ; et les ADN topoïsomérases de type II (multimériques et ATP-dépendantes), qui réalisent une coupure double-brin de l’ADN. Ces dernières ont une importance particulière dans la séparation des chromosomes durant l'anaphase. Parmi elles, la gyrase est capable d'introduire dans l'ADN des supertours négatifs.
Son mécanisme d’action permet à la molécule d’ADN de se relâcher, de produire des super hélices et de faire ou défaire des nœuds. La topoisomérase II va venir se fixer à l'aide du magnésium afin que l’enzyme puisse s’accrocher aux doubles brins de l’ADN et celle-ci va changer de conformation en se fixant au segment d’ADN. Par la suite, deux molécules d’ATP sont essentielles afin d’effectuer une coupure sur l’un des deux segments et d’en faire passer la seconde. En effet, la captation de l’ATP amène un changement conformationnel du domaine N-terminal des monomères, entraînant la prise du double brin d'ADN. On peut constater une libération du segment d'ADN lorsque les deux extrémités du premier segment d'ADN se rejoignent à l’aide d’une ligase.
Le processus de coupure-refermeture de l'ADN correspond à une réaction de transestérification réversible catalysée par le groupement OH d'une tyrosine appartenant au site actif.
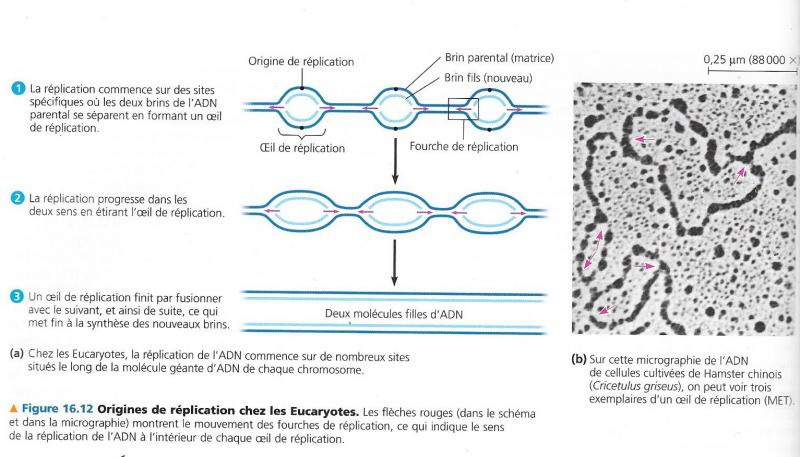
3. Propagation bidirectionnelle de la réplication
Après l’action des topoisomérases et girases, des hélicases séparent les deux brins de DNA., formation d'un oeil de réplication
Puis, des polymérases sont actives, commençant la synthèse par un brin du DNA (du côté 3' : brin direct), complétant ensuite sur le brin retardé (voir point suivant), à travers une RNA primase.
Chez l'homme, toutes les enzymes impliquées ne sont pas totalement connues : on distingue des polymérases a , b , g , d et e n'ayant pas toutes les propriétés des polymérases des procaryotes chez qui l’on distingue les polymérases I, II et III.
Des protéines accessoires de la réplication existent chez l'homme : le PCNA (proliferating Cell Nuclear Antigen) est le premier décrit. Il s'agit d'un puissant activateur de la polymérase d , stimulé lors de la division cellulaire. Le PCNA semble avoir aussi un rôle dans la réparation des anomalies du DNA (cf. plus bas).
Certains antibiotiques inhibent l’action de ces enzymes. Chez les bactéries, arrêt de réplication signifie mort.
Les brins séparés de l’ADN sont stabilisés sous forme simple brin grâce à la fixation de protéines appelées SSB (pour « single strand binding »). Ces protéines SSB empêchent les deux brins d’ADN de se réapparier.
A partir du point d’initiation se formera cet œil de réplication qui va s’agrandir jusqu’à la fin de la réplication et la séparation des deux ADN bicaténaires.
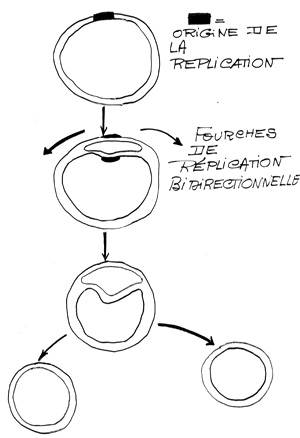
Agrandissement de l’oeil de réplication , progression de la réplication
La fourche de réplication est une transformation de la forme de l'ADN. lorsqu'un œil de réplication est créé d'autres enzymes interviennent, les hélicases. Ces enzymes vont se fixer aux extrémités de l’œil de réplication et la topoisomérase commence à dérouler l'ADN pour enlever les supers tours. L'hélicase va détacher les deux brins, pour pouvoir augmenter la taille de l’œil de réplication. En se faisant les hélicases vont transformer les extrémités des yeux de réplication en forme de Y, appelées fourches de réplication.
La fourche de réplication est la structure formée lorsque l’ADN se réplique, et sur laquelle l'ADN polymérase vient se fixer. L'ADN polymérase est une enzyme catalysant la formation des liaisons nucléotidiques. Le complexe enzymatique intervenant dans la réplication est appelé réplicase.
La réplication peut être divisée en trois étapes principales : l’initiation, l’élongation et la terminaison.
En résumé
En [1], l’hélicase sépare les deux brins du DNA. + action des topoisomérases
En [2], les protéines de liaison empêchent les deux brins de se recoller.
En [3], les polymérases synthétisent les nouveaux bras de 5’ en 3’.
En [4], le brin direct est retranscrit directement.
En [5], le brin retardé est synthétisé sous forme d’éléments d’Okazaki, qui sont reliés entre eux grâce à une ligase.
4. Réplication discontinue
Lors de la division cellulaire, les deux chaînes de DNA sont séparées à une extrémité. La lecture du code génétique se fait, chez les Procaryotes, par l’ADN polymérase III de l'extrémité 3' vers l'extrémité 5' ( brin matrice), par appariement des bases selon la loi de Watson et Crick. La molécule fille croît donc de l'extrémité 5' vers l'extrémité 3' .t elle ne peut croître que dans ce sens.
Par conséquent, la molécule d'ADN étant anriparallèle, un des brins se présente à la polymérase par son extrémité 3', par la complémentarité, ce brin peut donc être directement copié et construire le brin fille dans le sens 5'-->3', l'autre brin d'ADN lui se termine par une extrémité 5'P, il devrait donc ce construire dans le sens 3'-->5', ce qui est impossible, on aura alors une réplication discontinue
La synthèse de DNA est semi discontinue, et nécessite un segment d’initiation à base de RNA ou. Un des brins du DNA, celui ayant l’extrémité 3’ constitue le brin leader( brin matrice) : sa copie se fait directement par l’intermédiaire de la DNA polymérase III. L’autre brin nécessite la synthèse de petits fragments (fragments de Okazaki .Les fragments d'Okazaki ont entre 100 et 200 nucléotides de long chez les eucaryotes et entre 1000 et 2000 chez les procaryotes), en sens inverse du brin leader (de 5’ vers 3’) grâce à la même DNA polymérase III. Ces fragments sont ensuite rassemblés par une DNA polymérase I et par une DNA ligase.
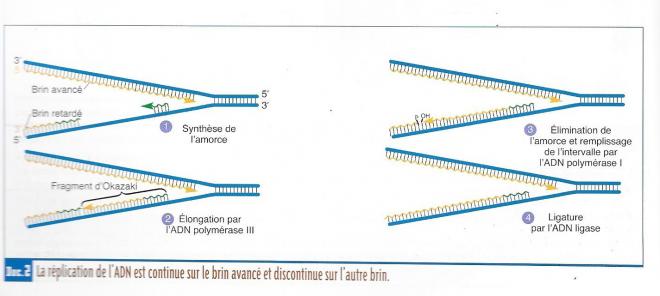 Schéma de la fourche de réplication.avec répliction discontinue et continue
Schéma de la fourche de réplication.avec répliction discontinue et continue


5. Initiation de la réplication.
Que la réplication soit continue ou discontinue, l’ADN polymérase III ne discerne pas les sites ou elle doit commencer son élongation, elle ne peut qu’ajouter un nucléotide à une extrémité 3’ OH.
En fait, c’est une ARN polymérase primase qui initie la réplication en produisant une amorce d’ARN à partir de laquelle pourra travailler l’ADN polymérase III.Cette primase fait partie d’un complexe volumineux parfois appelé « Primosome » ( primase + 16 polypeptides) L’amorce formée comporte quelques dizaines de ribonucléotides.
6. Hydrolyse et remplacement des fragments d’ADN
Les amorces seront détruites et remplacées par de l’ADN.
Ce travail est effectué par une ADN polymérase I qui a deux propriétés :
1. Propriété exonucléasique qui sert à hydrolyser les amorces
2. Propriétés polymérasiques qui lui permet de remplacer amorces par de l’ADN.
7. Soudure des morceaux d’ADN.
Les fragments d’ADN seront finalement reliés par une ligase.
8. La fonction d’édition
L’enzyme ADN polymérase III a deux propriétés :
- Propriété exonucléasique 3’ 5’
- Propriété polymérasique bien sûr
La fonction d’édition est à mettre en relation avec cette première propriété permettant de « relire » le dernier nucléotide mis en place.
Cette fonction consiste à retirer un nucléotide mal apparié et à le remplacer par le nucléotide approprié ( fonction d’excision – réparation.).
9. Mécanisme présidant au choix du nucléotide correct
La sélection des nucléotides résulte d’un équilibre entre plusieurs réactions en compétition ; il est possible de placer n’importe quelle base en face d’une autre, mais du point de vue énergétique, l’appariement correct est le plus favorable.
La stabilité du complexe « ADN polymérase – matrice ADN – nucléotide triphosphate est maximum lorsque ce nucléotide triphosphate est complémentaire du nucléotide de la matrice .
Voilà la raison pour laquelle l’ADN polymérase ne peut commencer une chaîne d’ADN ,elle doit toujours pouvoir relire le dernier nucléotide mis en place.
Grâce à ce mécanisme haute fidélité de la réplication . L’ADN nouvellement synthétisé comporte environ un nucléotide non complémentaire sur 100.000.
Un autre système enzymatique par exonucléases , permettrait encore de diminuer ce pourcentage d’erreur.
Notons l’importance de l’erreur de réplication vue sous l’angle des mutations.
Les mutations sont un des moteurs de l’évolution.
Rem : succession des enzymes pour le brin retardé :
1) primase + hélicase ( primosome) , 2) topoisimérase I et II , 3) ADN polymérase III
‘4) ADN polymérase I, 5) ligase
II.5.2 La réplication chez les Eucaryotes
Le mécanisme de réplication est identique :
- Réplication bidirectionnelle.
- Réplication complémentaire, antiparalléle.
- Discontinuités pour un brin.
- Zones amorces
Différence : L’ADN est plus long que chez les Procaryotes il y a donc plusieurs points d’initiation. Notons également que la réplication doit s’accompagner simultanément de la synthèse d’histones pour un brin sur deux
La réplication se fait en de nombreux points d’initiation. Elle fait intervenir un nombre d’ADN polymérases plus important que chez les procaryotes. De nombreuses protéines interviennent comme facteurs de réplication.
On connaît mal les changements subis par les nucléosomes au cours de la réplication de l’ADN eucaryote.
On connait au moins 5 ADN polymérases chez les eucaryotes.
- La polymérase alpha/primase. Cette polymérase est impliqué dans l'initiation de la réplication (tétramère protéique PM 180.000,55000,75000, 50000).
- La polymérase béta.
- La polymérase gamma. Cette polymérase est à localisation mitochondriale, bien que codée par un gène du noyau cellulaire.
- La polymérase delta.
- La polymérase epsilon.
La polymérase alpha/primase synthétise les amorces d'ARN. Une protéine appelée PCNA ("proliferating cell nuclear antigen") intervient.
L'ADN simple brin au cours de la réplication est stabilisé par des protéines correspondant aux protéines SSB du colibacille.
Les amorces d'ARN sont détruites par la RNase H. Les lacunes formées sont comblées par les polymérases béta ou alpha.
Chez les Eucaryotes, il semble que l’origine de la réplication est située aux points d’ancrage de la chromatine aux protéines du cytosquelette nucléaire
V. LE COURANT D’INFORMATION CELLULAIRE
V.1 Principe
- Dans le noyau : notamment composé d’ADN comportant un nombre très élevé de séquences. L’ADN est , en quelque sorte » une phrase écrite avec un alphabet de quatre lettres.
- Dans le cytoplasme : Présence de protéines édifiées avec 20 acides aminés différents. Beaucoup de ces protéines ont un rôle fonctionnel. On peut de manière similaire à l’ADN, considérer les protéines comme des phrases écrites avec un alphabet de 20 lettres.
On sait actuellement que la structure primaire des protéines se trouve sous forme codée dans la molécule d’ADN.
Conclusion : de manière imagée, on peut dire que : A un mot protéine correspond un mot ADN. Ce mot ADN , c’est le gène composé d’une somme de paires de nucléotides.
Mécanisme de transfert de l’information
ADN_____1______ ARNm_____2______ Protéine.
1)= LA TRANSCRIPTION
2)= LA TRADUCTION
![]()
Dans ces mécanismes interviennent essentierllement deux types d'ARN :
ARNm: ARN messager, au niveau de la transcription
ARNt : ARN de transfert : au niveau de la traduction
Date de dernière mise à jour : 17/06/2020
Ajouter un commentaire