
bio moléculaire 3
Nous avons ainsi défini la structure primaire d'un acide nucléique comme étant la suite de ses résidus nucléotidiques unis par des liaisons phosphodiester 3'-5'
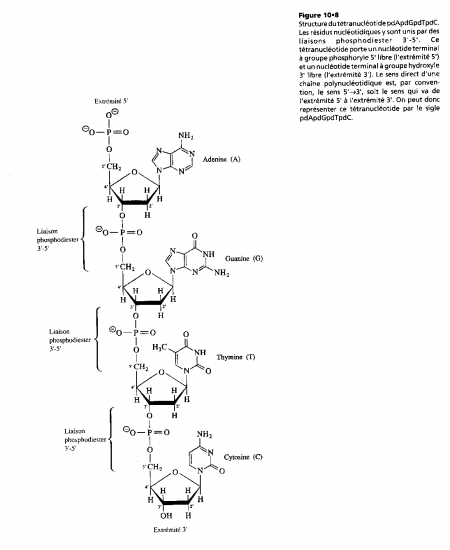
Les groupes phosphoryle en liaison diester portent chacun une charge négative à pH neutre ( pKa voisin de 2) ; aussi, les acides nucléiques sont ilsdes polyanions dans les conditions physiologiques , ils sont normalement complexés à des contre-ions tels Mg2+ ou a des protéines cationiques ( contenant une proportion élevée de résidus basiques d'arginine ou de lysine)
III.3.2 La structure de Watson et Crick (structure secondaire), l'ADN-B
Les chaînes monocaténaires que nous venont d'expliciter ne correspondent cependant pas encore à la véritable structure primaire des molécules d'ADN.
L'élucidation de la structure primaire de l'ADN par Watson et Crick en 1953 marque, dit-on , l'avènement de la biologie moléculaire moderne, à cette époque, on parlait de structure tout court car on ne savait pas encore comment l'ADN s'organisait spatialement.
Cette découverte , considérée comme un des plus beaux succès de la démarche scientifique, réconciliait de nombreuses données, dont certaines étaient encore loin d'ëtre acceptées par tous :
1) Les règles de Chargaff. A l'époque, les relations A=T et G=C restaient inexpliquées même par Chargaff
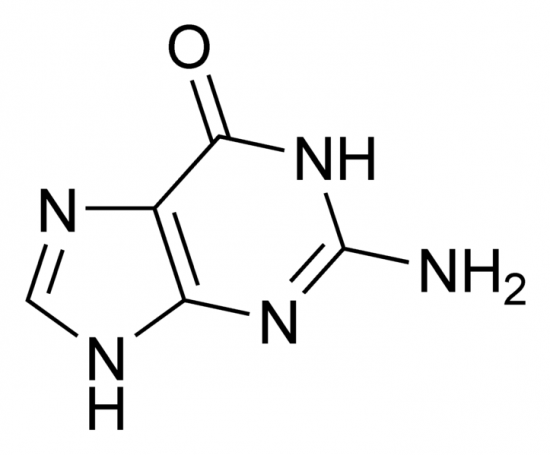
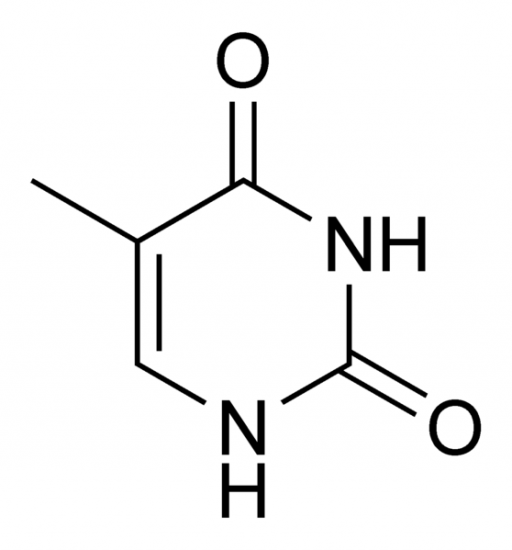
2)On pensait que la Guanine et la Thymine étaient sous forme énolique dans la molécule alors que c'est la forme céto qui est la plus courante.
Guanine (cétone)
Thymine (cétone)
3) On était pas encore persuadé de la forme hélicoïdale de la molécule d'ADN mais la photographie d'une molécule d'ADN diffractant les rayons X prise par Rosalind Franklin y faisait penser.
http://www.sfpnet.fr/fichiers_communs/commissions/SFP-Femmes/fichiers/rosalind.pdf
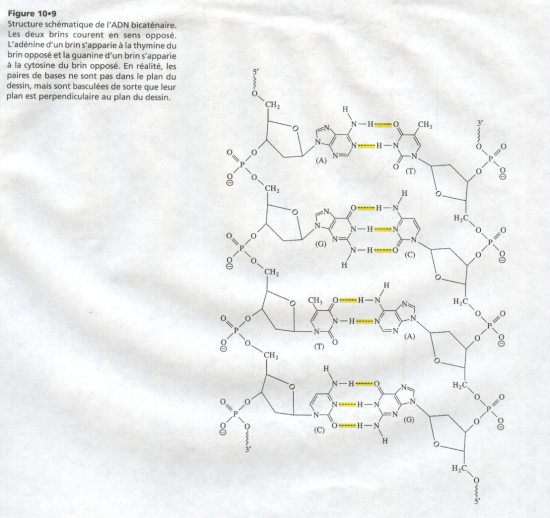
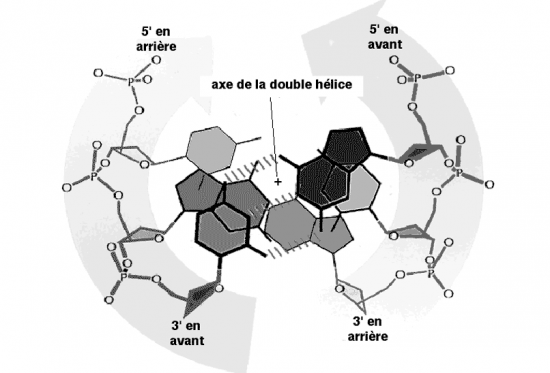
L'ADN est,en réalité,constitué de deux brins antiparallèles d'acide nucléique.
Association de deux chaînes polynucléotidiques.
Ces deux chaînes ont trois caractéristiques :
- Elles sont antiparallèles
- Elles sont complémentaires
- Elles sont hélicoïdales : double hélice de Crick et Watson.
a) Antiparallèles
5’ = = 3’
3’ = = 5’
b) Complémentarité
Règle de complémentarité ( Chargaff) :
A T
C G
Il résulte de cette propriété que deux chaînes de nucléotides peuvent se lier ensemble par leurs bases azotées si leurs bases sont complémentaires (i.e. A face à T et C face à G) ce qui est le cas dans la molécule d'ADN.
L'ADN est donc une double chaîne de nucléotides complémentaires.

Comment expliquer cette complémentarité
Deux explications :
1° Complémentarité pour des raisons stériques, question de place, d’encombrement. Une base purique a deux cycles tandis qu’une base pyrimidique n’en a qu’un. Chaque paire de bases aura donc la même dimension ce qui implique une structure régulière de la double hélice.
2° Complémentarité pour une raison de liaisons hydrogène ( formule ).
- entre A et T : 2 liaisons hydrogène
- entre G et C possibilité de 3 liaisons hydrogène.
L’édifice constitué par l’association de deux chaînes peut se diviser en deux grandes parties :
- Un squelette externe formé par l’assemblage P-S-P-S-P-S-P……..
- Une partie centrale formée par les séquences de paires de bases. Séquences caractéristiques de chaque molécule d’ADN.
Cohésion de l’association composant l’ADN
Les facteurs de cohésion de cette molécule sont :
- L’existence de liaisons hydrogène.

Une liaison hydrogène est une liaison de faible énergie entre deux atomes attirés l’un vers l’autre pour des raisons électrostatiques l’un étant riche en électrons donc nucléophile et l’autre n’ayant que les protons de son noyau donc électrophile.
• Ainsi l’atome d’hydrogène, dont l’unique électron est par nécessité dans l’orbitale qui unit cet atome au reste de la molécule, ne peut qu’être électrophile et comme tel attiré par les atomes ayant des doublets électroniques libres. • Les atomes d’azote et surtout d’oxygène possèdent respectivement deux et quatre électrons
de leur couche périphérique qui ne participent pas aux orbitales des liaisons covalentes de la molécule. Ceci leur confère un caractère nucléophile qui leur permet d’exercer une attraction sur les atomes électrophiles voisins, en particulier les atomes d’hydrogène : cette attraction
constitue une liaison hydrogène. • Lorsque les orbitales qui unissent d’un côté l’atome d’hydrogène à la molécule de gauche et de l’autre côté les doublets électroniques libres avec l’atome nucléophile sont dans le même
axe, la liaison hydrogène est plus forte.
- L’existence de liaisons hydrophobes entre plans superposés de bases.
- L’hydratation des phosphates situés vers l’extérieur de la molécule.
La structure de l’ADN ressemble à une échelle dont les montants sont constitués par la succession P-S et les échelons par les associations de bases azotées 2 à 2.
Notons en guise de remarque que l’ADN est thermolabile ( t° de fusion ) par rupture des liaisons hydrogène. La température de fusion sera d’autant plus élevée que la molécule d’ADN concernée contient beaucoup de paires G-C.
Lorsqu’un acide nucléique est en solution les molécules forment des liaisons hydrogènes associant les nucléotides deux par deux, de sorte qu’un nucléotide à adénine se lie avec un nucléotide à thymine (ou à uracile dans un RNA) et un nucléotide à guanine avec un nucléotide à cytosine.
• On désigne cette liaison sous le terme d’hybridation.
• L’hybridation guanine-cytosine est plus stable (3 liaisons hydrogène, -63 kJ) que celle entre adénine et thymine.
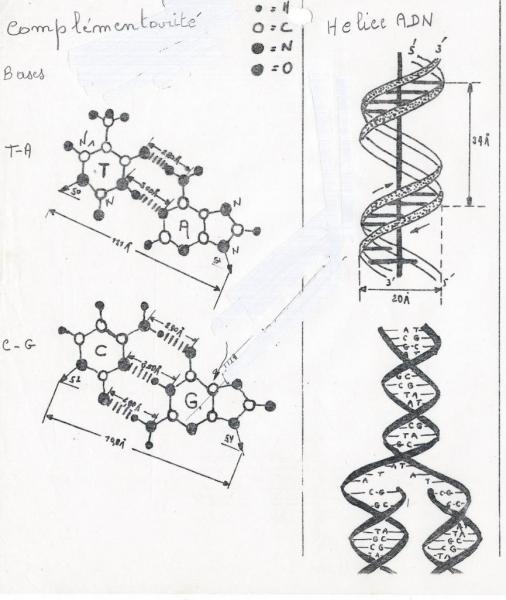
c) Hélicoïdale : double hélice droite.
La double hélice peut se présenter sous deux formes : ADN – A et ADN – B ( forme biologique la plus importante).
Les différences au niveau de ces deux formes d’ADN se situent au niveau :
- diamètre de l’hélice
- pas de l’hélice
- orientation des paires de bases.
ADN-B
- Diamètre : 2,37 nanomètres
- Pas de l’hélice : 3, 40 nanomètres
- Distance entre paires de bases : 0,33 nanomètres.
- Paires de bases perpendiculaires à l’axe de l’hélice.
- 10 paires de bases par tour.
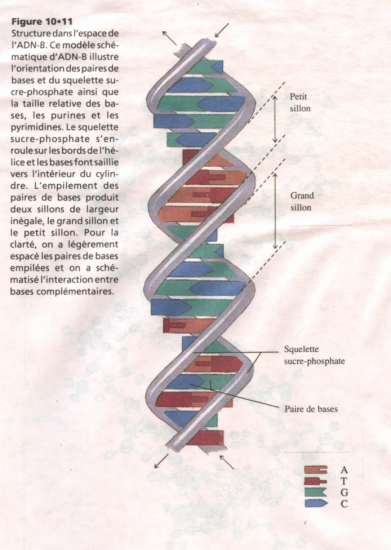
Structure schématique
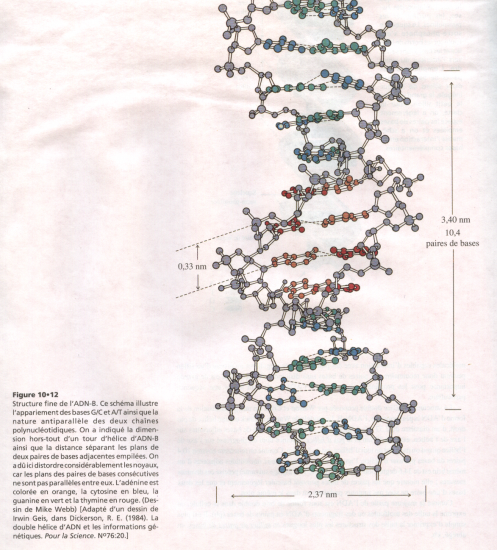
Structure détaillée
Remarques :
- Unité de longueur de l’ADN, le kilo-bases(Kb) = 1000 paires de bases.
- Nombre de paires de bases par génome haploïde :
Virus : +/- 10.000
Bactéries : 4,6 .106
Insectes : 1 . 109
Hommes : 2,8 . 109
Chez certains autres Vertébrés : 1011
L’ADN peut être circulaire ou linéaire.
d) hélicoïdale, double hélice gauche ADN – Z
Dans les conditions physiologiques, il a été possible d’obtenir l’ADN-Z en méthylant des séquences au niveau des cytosines (voir note) ou en opérant au niveau de certains polyamines comme la spermine.
L’ADN – Z n’est pas l’image en miroir de l’ADN classique . Sa conformation est différente :
Squelette P – S – P en zig zag
Hélice plus svelte, plus torsadée que le B – ADN : 12 pdb/tour de spire, le pas de l’hélice est de 4,46 nanomètres et le diamètre de 1,8 nanomètres.
Les bases sont plus exposées dans ce type d’ADN , l’ADN peut passer de façon réversible d’une forme à l’autre. L’enseignement que l’on peut tirer de cette constatation est que l’ADN est une structure dynamique qui est capable de subir :
- Des déformation au sein de l’ossature de chaque chaîne
- Des mouvements plus lents au niveau des bases qui s’ouvrent et se ferment
On dit que : « l’ADN respire »
Il est évident que toutes ces modifications de structure sont perçues par les molécules interagissant avec l’ADN.
Exemples
- voir méthylation des gènes
- Certaines protéines sont capables de se lier spécifiquement avec ADN-Z mais non avec B-ADN.
- Ces ADN-Z binding protéines doivent jouer un rôle important dans la régulation de l’expression génique.
Cet ADN Z est rencontré dans un milieu naturel lorsqu'il y a une abondance de séquences G-C. Les bases restent bien centrées à l'axe de la double hélice et il y en a 12 par tour.
Les molécules d’acide désoxyribonucléiques sont formées de deux chaînes dont les nucléotides ont hybridés deux à deux sur toute la longueur.• Les deux chaînes sont antiparallèles, c’est à dire que l’extrémité 5’ de l’une est du côté de l’extrémité3’ de l’autre.
• Pour que tous les nucléotides puissent s’hybrider ; il faut que l’ordre dans lequel ils sont liés ensemble soit complémentaire de la chaîne opposée.
• Les bases azotées liées par les liaisons hydrogènes sont tournées vers l’intérieur, tandis que les riboses et les acides phosphoriques, hydrophiles sont tournés vers l’extérieur.
• La chaleur peut dissocier les deux chaînes : c’est la fusion du DNA. Cette fusion est réversible : les deux chaînes peuvent s’hybrider à nouveau.
La structure secondaire du DNA est telle que les deux brins sont enroulés l’un autour de l’autre. Chacun des deux brins est orienté (5’→3’) dans le sens opposé à celui de l’autre brin (3’→5’). On dit qu’ils sont antiparallèles.
• Les bases azotées sont tournées vers l’intérieur de la double hélice de façon à ce que chacune s’hybride avec une base de l’autre brin (A avec T, C avec G, etc..). On dit que les bases successives de chacun des brins sont complémentaires.
• La double hélice a un « pas » de 3,4 nm c’est à dire qu’il y a environ 10 paires de nucléotides pour chaque tour d’hélice.

Une vue perspective de la double hélice montre bien comment les bases azotées sont parallèles entre elles, leurs noyaux empilés comme des assiettes au centre de la double hélice.
Date de dernière mise à jour : 17/06/2020
Ajouter un commentaire


