
Biologie cellulaire 6
II.3. APPAREIL DE GOLGI
II.3.1 Structure
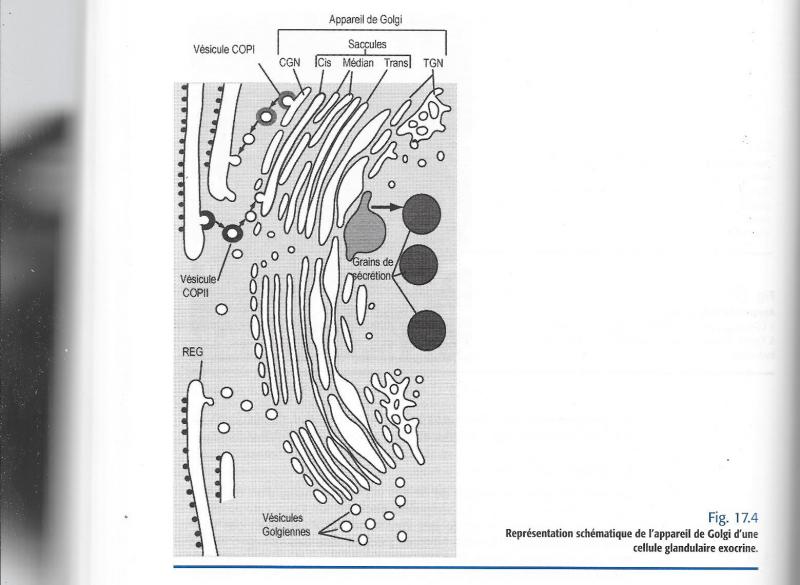
L’unité de structure est le dictyosome (constitué de citernes).Le dictyosome est formé d’un empilement de saccules entourées et bordées de vésicules.
Les saccules se divisent en trois compartiments distincts se différenciant par leur bagage enzymatique :
Le compartiment cis, côté réticulum.
Le compartiment médian
Le compartiment trans L'appareil de Golgi est formé d'un ensemble de sous-structures appelées "dictyosomes".
L'illustration ci-après présente un dictyosome. Il est formé d'un empilement de saccules, délimités par une membrane du même type que la membrane du réticulum endoplasmique.
Ces saccules sont incurvés; c'est ce qui donne au dictyosome sa forme caractéristique.
L'extrémité de chaque saccule tend à "s'effilocher" sous la forme de petites vésicules qui s'éloignent du dictyosome.
Le nombre, la forme et la dispostion des dictyosomes dépend :
- Du type de cellule
- De l’état fonctionnel de la cellule
II.3.2 La membrane golgienne
II.3.2.1 Rapport L/P : 35/65
II.3.2.2 Lipides :
forte similitude avec la membrane du réticulum
II.3.2.3 Protéines :
- beaucoup de glycosyltransférases ( fin de glycosylation ).
- beaucoup de sulfotransférases.
- des phosphatases ( récupération du phosphore).
- une chaîne de transport d’électrons
II.3.2.4 Différences avec la membrane du réticulum :
- pas de cytochrome P450
- pas de glucose 6 phosphatase
- pas d’enzymes impliquées dans la synthèse des lipides.
II.3.2.5 Remarque
Malgré ces différences, il n’y a pas d’enzymes marqueurs caractéristiques de ces membranes golgiennes .
Les enzymes sont pour la plupart des protéines intégrales.
Les membranes sont également asymétriques.
Les portions glucidiques des lipides et des protéines sont située en regard de la lumière de la cavité.
II.3.3 Le contenu des cavités
Le compartiment golgien et le compartiment réticulum contiennent les mêmes constituants dans des proportions et concentrations souvent différentes. Souvent elle contient plus de polysaccharides.
II.3.4 Les rôles physiologiques
II.3.4.1 place et rôle de l’appareil de golgi dans le processus de sécrétion
Schéma général
Cheminement d’un polypeptide synthétisé par les ribosomes fixés sur le réticulum :
- Emission par le RE de vésicules de transition contenant la molécule.
- Fusion de ces vésicules avec la première saccule cis.
- Passage d’une saccule à l’autre par des vésicules latérales.
- Emission de vésicules de sécrétion par le dernier compartiment trans.
- Migration,éventuelle de ces vésicules vers la membrane plasmique.
- Fusion avec la membrane plasmique et exocytose.
Les vésicules de sécrétion, fusionnent en général, avec des grains de sécrétions plus volumineux.
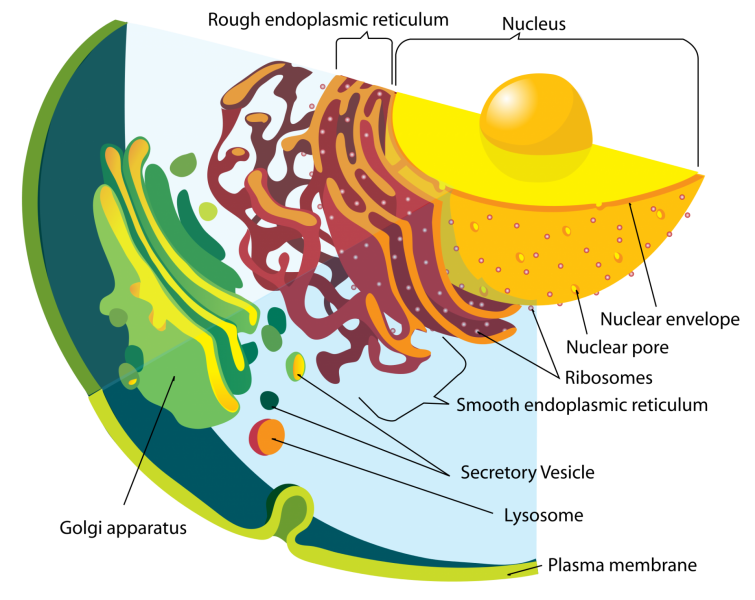
APPAREIL DE GOLGI
II.3.4.2 La membrane
- Sert à emballer les produits de sécrétion ( vésicules et grains de sécrétion).
- Les glycosylations
Celles-ci sont catalysées par les transférases des membranes et correspondent à l’élongation et à la terminaison des chaînes polysaccharidiques.Les glycolysations ont été initiées dans le RE ( fixation de N acétyl galactosamine) et sont terminées dans l’appareil de golgy, par l’addition des résidus glucidiques les plus périphériques ( fructose, mannose, acide sialique )
la glycolysation sera différente selon la nature des molécules à emballer (vésicules de sécrétions ou lysosomes). Les destinations des glycoprotéines sont les suivantes : 1) glycoprotéines intracellulaires ( enzymes des lysosomes), 2) les glycoprotéines extracellulaires qui se divisent en glycoprotéines de surface dont on connaît les propriétés antigéniques spécifiques et en glycoprotéines excrétées.
Les protéines destinées à utiliser les voies de biosynthèse/sécrétion intègrent de façon co-traductionnelle la lumière du réticulum endoplasmique (RE), compartiment dans lequel la protéine va subir les premières étapes du repliement et certaines modifications telles que la glycosylation et la formation de ponts disulfures. Les protéines naissantes seront aidées dans ces différentes étapes de maturation par des protéines résidant dans le RE telles que les protéines chaperons qui assurent le bon déroulement du repliement et évitent les interactions intermoléculaires indésirables, les enzymes de glycosylation (glycosidases) et enfin la protéine disulfide isomérase qui permet aux ponts disulfures de se former correctement. Les protéines, qui peuvent s'assembler en oligomères ou rester monomérique, subissent un contrôle qualité avant d'être regroupées dans des régions spécialisées du RE, aussi appelées sites de sortie (ERES pour Endoplasmic Reticulum Exit Sites). En cas de défaut, les protéines sont orientées vers la voie de dégradation cytosolique par le protéasome. Les protéines présentes dans les ERES sont ensuite acheminées vers le compartiment intermédiaire (ERGIC, pour Endoplasmic Reticulum/Golgi Intermediate Compartment), compartiment de triage entre les protéines qui retournent vers le RE et celles qui continuent leur route vers le Golgi. Traversant les différentes citernes du Golgi (dans l'ordre du transport antérograde, cis-,médian et trans-Golgi), les ramifications oligosaccharidiques des protéines vont être modifiées
de façon séquentielle par les glycosidases ou glycosyl-transférases rencontrées. Certaines protéines retourneront vers le RE par transport rétrograde à partir des saccules du Golgi mais la plupart atteindront à ce stade le réseau trans-golgien (RTG ou TGN pour Trans-Golgi Network). De nouvelles modifications post-traductionnelles peuvent encore intervenir, telles que la sialylation ou la sulfatation des sucres ou encore le clivage protéolytique par des protéases comme la furine. Le TGN permet le triage des protéines à un stade tardif du transport et est impliqué dans la spécification de l'adressage des protéines vers la voie lysosomale ou vers des domaines spécialisés de la membrane plasmique. En fonction des voies empruntées à partir du TGN, les protéines seront sécrétées de façon constitutive ou régulée, et pourront être envoyées vers les régions apicales ou basales distinctement dans le cas des cellules polarisées. Ce compartiment est aussi particulièrement important car il fait le lien entre les voies d'exocytose et d'endocytose.
Les protéines provenant du milieu extracellulaire pénètre dans la cellule cible soit par un mécanisme non spécifique de pinocytose, soit par recrutement par des récepteurs membranaires. L'internalisation des molécules se fait par l'intermédiaire d'endosomes précoces qui font le tri entre les protéines à recycler vers la membrane plasmique (c'est en général le cas pour les récepteurs libérés de leur ligand) et celles qui migreront via les endosomes tardifs vers les lysosomes. A partir des structures endosomales, il existe également une voie d'adressage des protéines vers le TGN. Les lysosomes constituent le principal site de dégradation des molécules (protéines, acides nucléiques, sucres et phospholipides) mais aussi de structures plus importantes (fragments d'organelles, corps étrangers...). L'hydrolyse des substrats se fait à pH acide sous l'action d'enzymes spécialisées et les produits de dégradation sont alors reconduits vers le cytosol pour être réutilisés ou relargués dans le milieu extracellulaire.
Les sulfatations
Elles consistent en la fixation d’un ou plusieurs radicaux sulfate sur de glycoprotéines ou des protéoglycanes, à l’aide des sulfotransférases.
Obtention : Chondroïtines sulfates, molécules des cornées et des cartilages, héparannes sulfates se logent dans les membranes basales, kératannes sulfate dans les cartilages…
Remarque : l’appareil de golgi aurait également une fonction protéolytique de type : Proprotéine -----> protéine active
II.3.4.3 Production de membrane pour la surface cellulaire.
Cette production est nécessaire pour deux raisons :
- Remplacement suite au départ de membrane par émission de vésicules d’endocytose et d’exocytose
- Renouvellement et recyclage des constituants membranaires.
L’appareil de golgi contribue à la biogénèse de la membrane plasmique , non seulement par rapport de substance membranaire nouvelle mais aussi par le rôle qu’il joue dans la synthèse des polysaccharides.
II.3.4.4 La cavité
Trois principaux rôles :
- Ségrégation et stockage de molécules endogènes très variées, la ségrégation de matériel d’origine exogène est plus rare .
- Transformation de molécules.
- Transit de molécules.
II.3.5 La biogénèse
L’appareil de golgi est une structure en équilibre dynamique
- Côté réticulum : face de formation ( arrivée de vésicules de transition )
- Côté opposé : face de maturation ( départ des vésicules de sécrétion et autres).
II.3.6 Résumé après l’étude de ces trois membranes
Lipides :
Memb du RE------>Memb. golgi --------> Memb. Plasmique
Augmentation de la teneur en lipides
Augmentation de la teneur en glycolipides et cholestérol
Augmentation de la longueur des chaînes des acides gras
Diminution de la saturation des chaînes d’acides gras.
Protéines :
Memb. RE--------> Memb. Golgi ---------> Memb. Plasmique
Beaucoup plus de discontinuités
Protéines plus ou moins spécifiques dans chaque cas.
II.3.7 L’appareil de golgi de certains types de cellules.
Exemple : La cellule acineuse pancréatique a une importante fonction de sécrétion exocrine, elles sécrètent diverses enzymes ou proenzymes ayant un rôle digestif :Trypsinogène,chymotrypsinogène,carboxypeptidase, ribonucléase, désoxyribonucléase,
lipase,amylase…
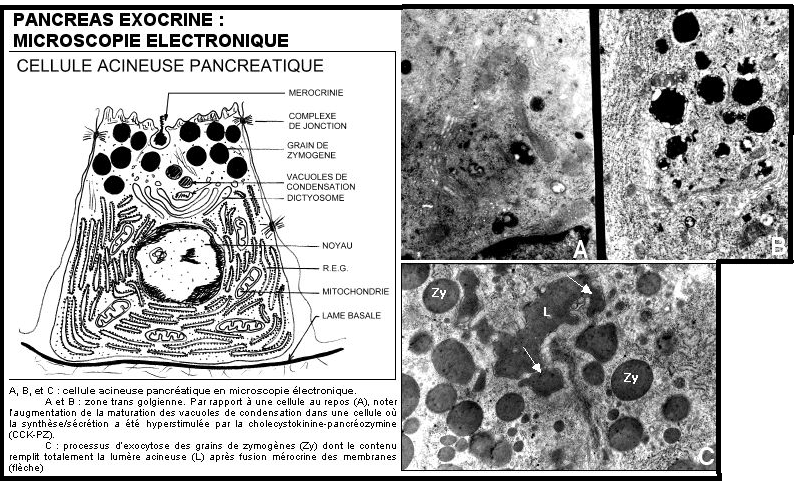
Les acides aminés pénètrent dans la cellule par la membrane basale et sont mis en contact avec le réticulum endoplasmique situé du côté basal et très volumineux. La synthèse protéique peut ainsi être initiée. Au dessus du noyau, l’appareil de golgi estégalement très développé.L’appareil de golgi émet des vésicules a contenu très dense ( densification par cheminement de saccule en saccule dans l’appareil de golgi) appélée zymogènes et contenant les enzymes digestives. Ces zymogènes seront excrété au niveau du pôle apical par exocytose
II.4. LE CYTOPLASME
Chez les procaryotes, le hyaloplasme est le milieu cellulaire où baignent tous les organites de la cellule. Chez les eucaryotes, c’est le milieu où baignent les organites cytoplasmiques ; il est limité par la membrane plasmique et séparé du noyau par la membrane nucléaire.
II.4.1 La structure.
Le cytoplasme ou cytosol apparaît comme un milieu homogène dans lequel on ne distingue aucune organisation structurale .Cependant, on peut y remarquer des différenciations morphologiques :
- Les globules lipidiques, présents dans de nombreux types de cellules , ils font une part importante du volume hyaloplasmique dans certaines cellules (adipocytes ).
- Ces structures ne sont pas figées dans l’espace et le temps ; des courantscytoplasmiques les entraînent d’une région à l’autre de la cellule et leur quantité varie au cours de la vie cellulaire.
II.4.2 La composition chimique
- Par ordre d’importance quantitative :
- L’eau = 85% en moyenne.
- Les protéines : beaucoup d’enzymes, des protéines de structures indispensables à l’édification des organites.
- Des ARN : ARNm, ARNt .
- Des sucres, des acides aminés, des nucléosides et des nucléotides.
- De très nombreux composés du métabolisme intermédiaire.
- Des ions variés.
II.4.3 Les rôles et activités physiologiques
II.4.3.1 Réserve de combustibles et de matériaux de construction.
II.4.3.1.1 Les réserves de combustibles
- Glucose.
- Globules lipidiques et particules de glycogène.
Ces deux derniers combustibles rassemblés en une phase hydrophobe, n’ont aucune influence sur la pression osmotique du hyaloplasme. Grâce à eux, la cellule peut stocker de grandes quantités de combustible.
L’alimentation de la cellule par le milieu extérieur ne doit donc pas nécessairement être continu.
La dégradation des combustibles se fait par des oxydations au niveau du hyaloplasme et des mitochondries. Elle produit l’énergie nécessaire à la régénération des nucléotises triphosphates. Principalement l’ATP.
Utilisation de l’ATP : Par hydrolyse de l’ATP, la cellule aura suffisamment d’énergie pour :
- activer certaines molécules
- réaliser de nombreuses biosynthèses
- réaliser les transports actifs d’ions et de molécules au travers des membranes.
- Réaliser les travaux mécaniques par les systèmes contractiles de la cellule.
II.4.3.1.2 Les matériaux de construction.
Matériaux de construction transformés et assemblés :
- Sur place dans le hyaloplasme :synthèse de divers oses, de mononucléotides, de certains acides aminés, de l’acide palmitique. Polymérisation du glycogène.
- Dans les ribosomes : Synthèse de chaînes polipeptidiques.
- Dans les membranes du réticulum et de l’appareil de golgi : allongement des chaînes polysaccharidiques des glycoprotéines et des glycolipides.
- Dans le noyau : synthèse des acides nucléiques
II.4.3.2 Carrefour des voies métaboliques.
Nombreuses réactions de dégradation ( catabolisme) et de synthèse (anabolisme) se déroulent dans le cytoplasme. Un seul exemple sera pris pour illustrer ce point : le métabolisme des glucides.
Les voies métaboliques du glucose 6 phosphate
-Le glucose introduit dans le cytoplasme est phosphorylé par une hexokinase, en glucose-6 phosphate et est en quelque sorte séquestré. Ce glucose 6 phosphate peut suivre trois voies métaboliques.:
II.4.3.2.1 la glycolyse
La glycolyse anaérobie est une chaîne métabolique qui permet à une cellule d'extraire de l'énergie directement du glucose. Cette chaîne, bien que peu rentable, est d'une importance fondamentale en biologie. Universelle, elle est partagée par tous les êtres vivants, des bactéries les plus primitives à l'homme. Les protéines qui les constituent sont les mêmes, les modes de régulation aussi.
Cette chaîne doit être aussi ancienne que la vie elle même et elle constitue certainement le plus ancien mode d'obtention de l'énergie encore existant aujourd'hui.
Du fait même de son ancienneté, et en dépit du peu d'énergie qu'elle peut extraire du glucose comparé à d'autres voies métabolique, elle est d'une efficacité incroyable en raison de son optimisation qui la conduit à une vitesse extrême, pouvant produire bien plus d'énergie par unité de temps que toute autre méthode. C'est par exemple elle qui alimente la contraction musculaire en cas d'effort intense.
Toutefois, son importance va beaucoup plus loin. Son ancienneté l'a placé à une position clef dans le métabolisme cellulaire. En effet de nombreuses voies métaboliques se greffent dessus, c'est à dire que le point de départ ou d'arrivée de certaines est un des métabolite de la chaîne. C'est par exemple le glycogène et l'amidon qui sont synthétisé à partir du glucose-6-phosphate (G6P), le second métabolite de la chaîne, de même que la voie des pentoses phosphates à l'origine des pentoses des acides nucléiques partage plusieurs métabolites avec cette chaîne et ne peut exister que grâce à elle. Les deux plus fréquentes sont toutefois celle qui se greffent sur le dernier métabolite de la chaîne, le pyruvate, qui sont les fermentations et le cycle de Krebs.
Le bilan brut de la chaîne, à partir d'une molécule de glucose, est :
C6H12O6 + 2 NAD+ + 2 ATP + 4 ADP + 4 Pi --> 2CH3CO COOH + 2NADH + 2H+ + 4 ATP + 2ADP + 2 Pi
Ce qui donne en bilan : C6H12O6 + 2 NAD+ + 2 ADP + 2 Pi --->2CH3CO COOH + 2NADH + 2H+ + 2 ATP
Date de dernière mise à jour : 16/06/2020
Ajouter un commentaire