

Biologie cellulaire 5
II.2. RETICULUM ENDOPLASMIQUE
II.2.1 Définition
Le réticulum endoplasmique est un système de saccules ou de canalicules limité par des membranes de nature lipoprotéique et comprenant deux compartiments qui communiquent l’un avec l’autre, mais qui diffèrent par leur constitution et leur fonction.
Ces deux compartiments sont :
-Le réticulum endoplasmique granulaire ( REG)
-Le réticulum endoplasmique lisse ( REL).
Ces deux compartiments intercommunicants ont un constituant commun de type membranaire , d’une épaisseur de 5 à 6 nanomètres, dont la structure est comparable aux autres cytomembranes.
Ils diffèrent par la forme des cavités limitées par ces membranes et par la présence sur la face externe, dans le cas du REG, de granulations : les ribosomes qui y sont accolés.
II.2.2 Structure
Le réticulum endoplasmique morcelle le volume cellulaire en deux compartiments :
- Le noyau qui sera entouré par la membrane nucléaire.
- Le cytoplasme.
Il ménage dans le cytoplasme des cavités de formes diverses
On distingue deux grandes parties dans le réticulum :
- Le réticulum granulaire, les granules étant des ribosomes nombreux fixés sur la membrane du réticulum
- Le réticulum lisse ne portant pas de ribosomes.
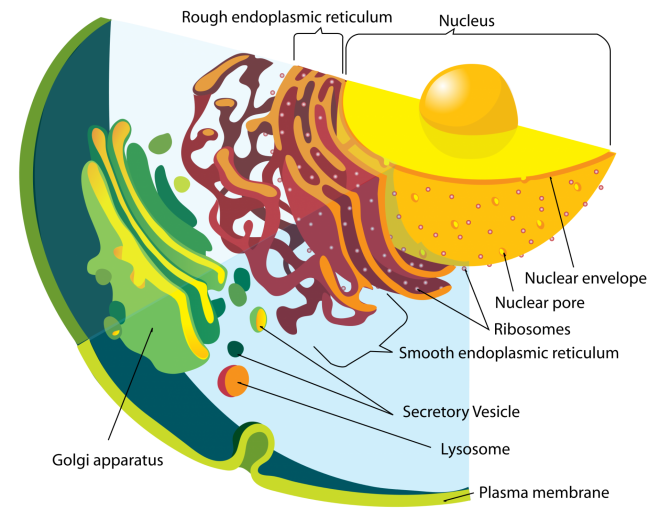
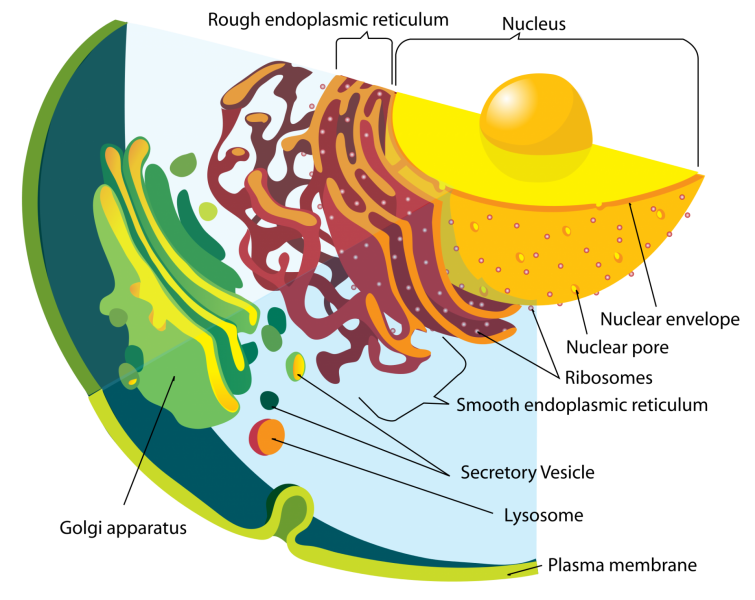
L’importance et la dispersion spatiale du réticulum varie en fonction du type de cellule et aussi en fonction de ses activités métaboliques.
II.2.3 Rôles
REG : Synthèse de protéines et de glycoprotéines qui seront finalement excrétées.
REL : Synthèse de lipides et lieu de détoxification de la cellule.
II.2.4 La membrane du réticulum
Par comparaison avec la membrane plasmique nous noterons les particularités suivantes :
Rapport L/P = 30/70
Lipides : Peu de cholestérol,peut de glycolipides
Protéines :
des protéines pompes et canaux,
des enzymes ( ex :cytochrome P450) catalysant des réactions d’hydroxylation,
deux chaînes de transporteurs d’électrons.
des hydrolases ( glucose 6 phosphatase)
nucléoside phosphatases,
glycosil transférases.
II.2.5 Le contenu des cavités
Une solution aqueuse d’holoprotéines de glycoprotéines et de lipoprotéines. Ce contenu est variable en fonction du type cellulaire, de l’état physiologique et de l’espèce.
II.2.6 Les rôles physiologiques
II.2.6.1 Le transfert de chaînes polypeptidiques ( synthétisées au niveau des ribosomes).
Le tout début de la synthèse d’une protéine destinée à être exportée du cytoplasme se produit sur un ribosome libre, puis la synthèse se poursuivra sur le même ribosome devenu lié à la membrane du réticulum dans le cas des eucaryotes. Le fait qu’un ribosome soit libre ou lié dépend seulement de la nature de la protéine qu’il est entrain de synthétiser.
Schéma : Transfert des chaînes protéiques
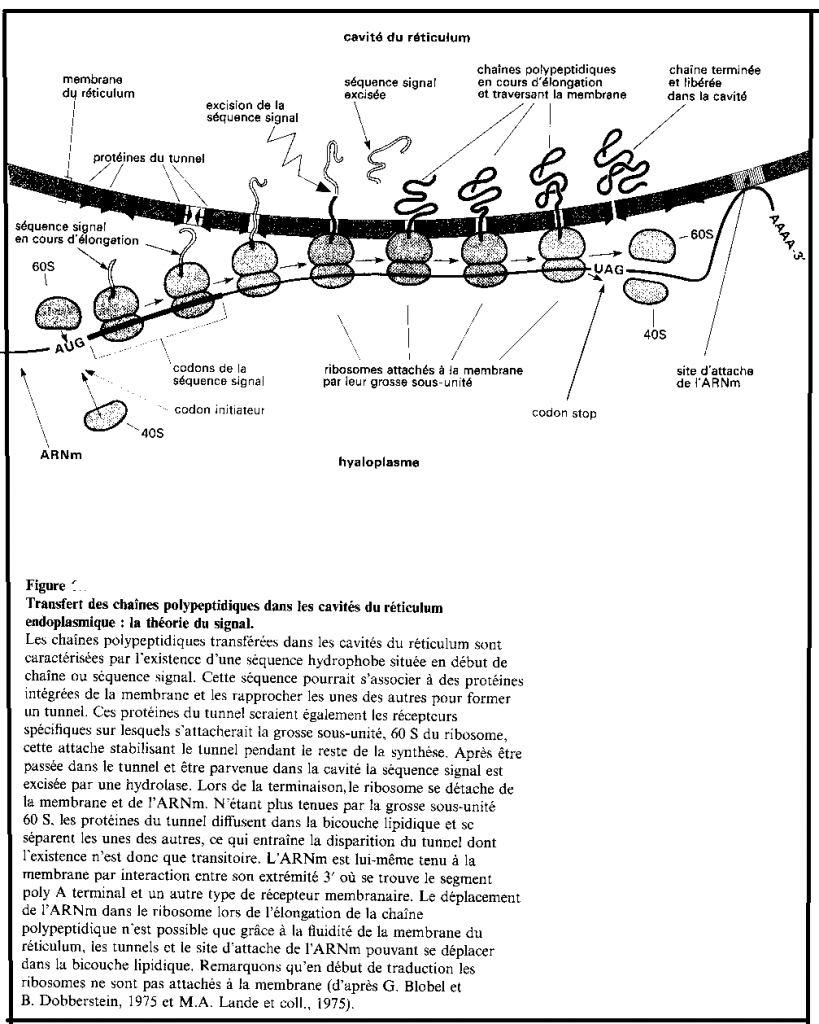
II.2.6.2 Au niveau membranaire
Les membranes du réticulum contiennent des systèmes enzymatiques qui catalysent de nombreuses réactions importantes du métabolisme de divers lipides : acides gras, phopholipides, cholestérol et ses dérivés.
Elongation et désaturation des acides gras.
L’acide palmitique qui est synthétisé dans les membranes par le système plurienzymatique -acide gras synthétase- est allongé par réactions successives avec le malonyl-coenzymeA. A l’état de thioester du coenzyme A, les acides gras sont désaturés. La formation de la double liaison est catalysée par une oxydase à fonction mixte qui utilise une molécule d’oxygène et une molécule de NADH. Le transfert des électrons à l’oxygène se fait par l’intermédiaire d’une chaîne dont l’un des constituants est sans doute le cytochrome b5 et il y a probablement formation d’un dérivé hydroxylé intermédiaire.Par exemple la déshydrogénation de l’acide stéarique en acide oleique se fait selon les réactions suivantes.
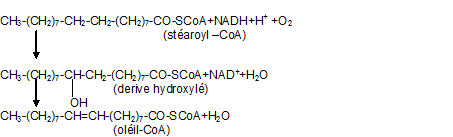
Ces réactions d’élongation et de désaturation sont particulièrement importantes dans les membranes du réticulum des hépatocytes et des adipocytes. Cette réaction fait intervenir différents enzymes et est en fait complexe ( voir biochimie )
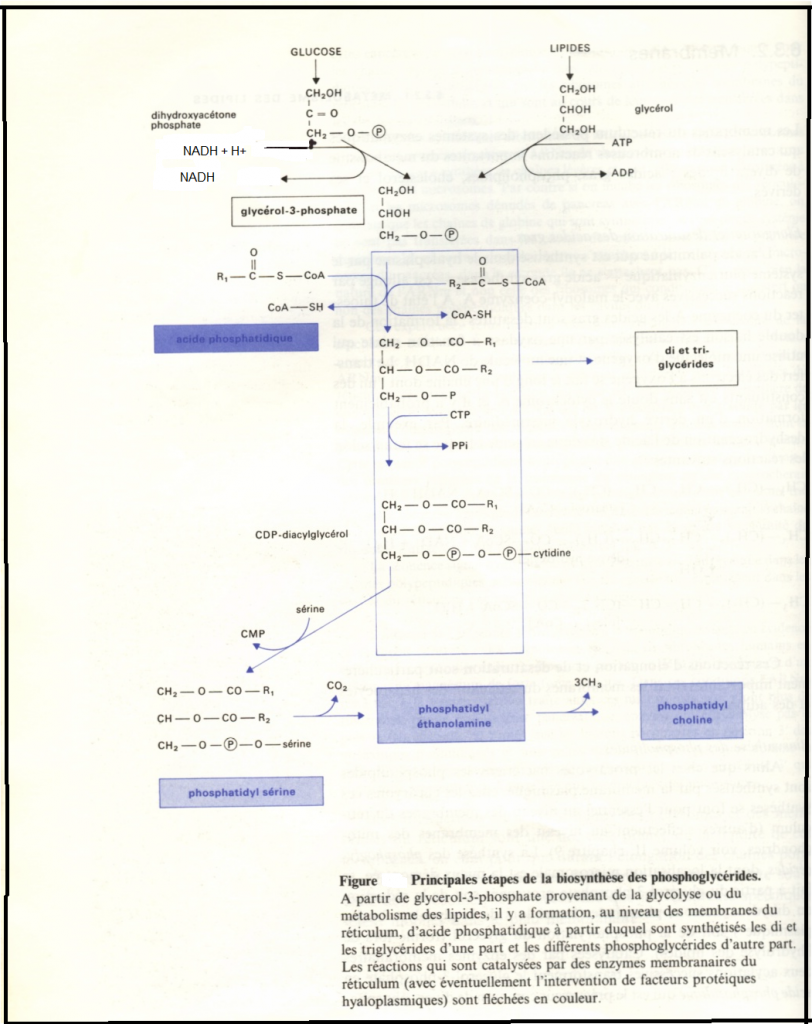
Biosynthèse des phospholipides
Chez les eucaryotes,ces synthèses se font essentiellement au niveau des membranes du réticulum, c’est la synthèse des phosphoglycérides qui y est la mieux démontrée.
Cette synthèse se fait à partir du glycérol-3-P provenant du dihydroxyacétone phosphate ( formé lors de la glycolyse aloplasmique) ou de la phopsphorylation du glycérol (issu de l’hydrolyse des lipides). Catalysée par des enzymes membranaires, deux acylations successives transforment le glycérol 3 phosphate en acide phosphatidique qui est le précurseur commun des divers phosphoglycérides ( et également des di et des triglycérides qui sont aussi synthétisés dans les membranes du réticulum.
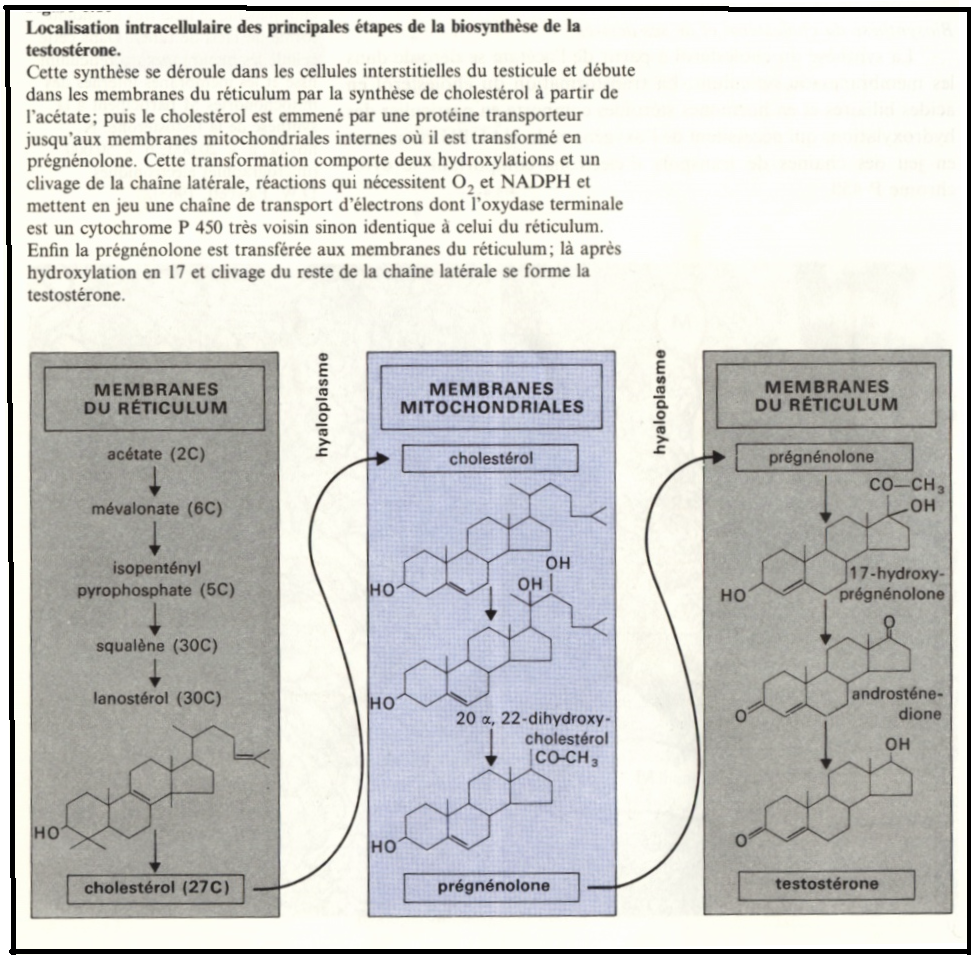
Biosynthèse du cholestérol
La synthèse du cholestérol à partir de l’acétate se déroule dans les membranes du réticulum. La transformation du cholestérol en acides biliaires et en hormones stéroïdes comporte en particulier des hydroxylations qui nécessitent de l’oxygéne et du NADPH et
mettent en jeu des chaînes de transporteurs d’électrons comportant le cytochrome P 450.
Dans les cellules hépatiques où est synthétisée la plus grande part du cholestérol des vertébrés les réactions qui conduisent à l’acide cholique et aux autres sels biliaires ont lieu au niveau des membranes du réticulum.
Dans les cellules qui synthétisent des stéroïdes (testicules, cellules du corps jaune dans l’ovaire, cellules interstitielles du testicule) et dont le RE est très développé, la synthése du cholestérol a également lieu dans ces membranes,mais seules certaines étapes de la biosynthèse des hormones se font dans le réticulum, les autres s’effectuent dans les membranes des mitochondries.
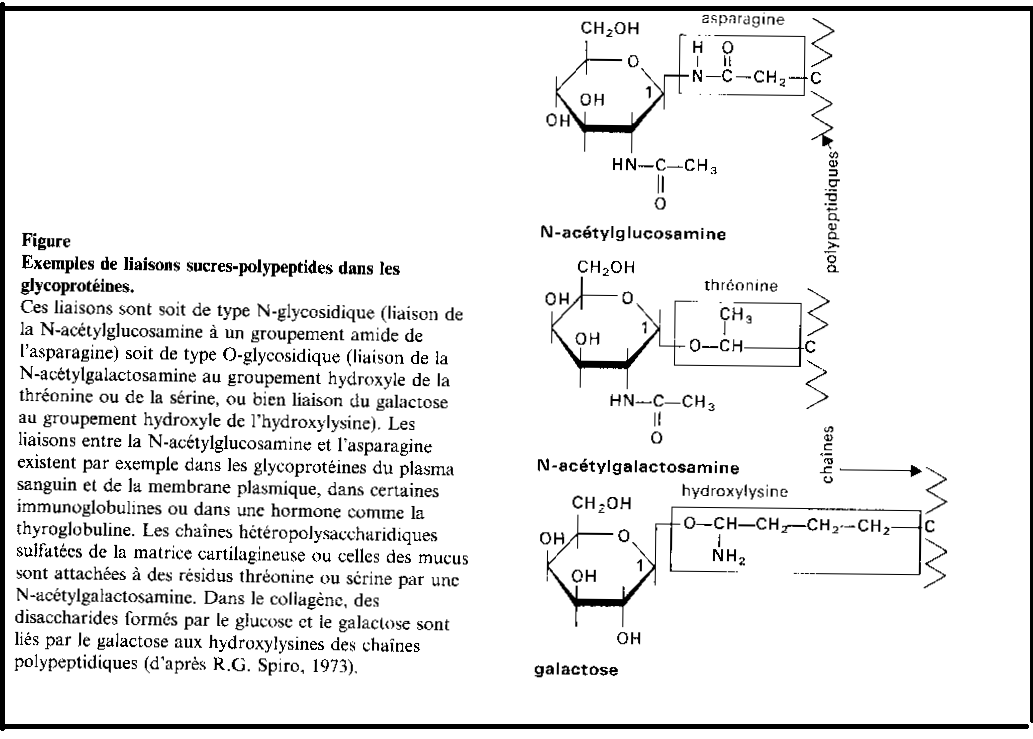
Les glycosylations
Les protéines synthétisées par le REG peuvent s’associer à des glucides. Cette synthèse de glycolipides porte le nom de glycosylation. Elle débute dans le REG et se termine dans l’appareil de Golgi.La glycosylation est initiée par une molécule intramembranaire du REG, le Dolichol. Les glucides ( liés à de acides nucléiques) traversent la phase lipidique de la membrane du REG et se combinent au Dolichol. Ces sucres sont la N-acétylglucosamine, le mannose hexosamine, le glucose. Ils se combinent au Dolichol ( en se liant avec la face en rapport avec la cavité du REG), dans l’ordre des énumérations précédentes, grâce a des glycosyltransférases spécifiques.
Pendant la synthèse d’une protéine, une glycosyl transférase transporte le polysaccharide complexé avec le Dolichol et le fixe sur la molécule protéique . Le polysaccharide en cours de synthèse, sous l’influence d’enzymes contenues dans la membrane du REG, perd la majorité de ses molécules de mannose tandis que d’autres sucres s’associent ainsi à la glycoprotéine assurant ainsi sa spécificité. Cette glycolysation s’achèvera dans les saccules de l’appareil de Golgi.
 - Détoxification.
- Détoxification.
Elle sur des molécules endogènes et exogènes ,captées par pinocytose, par hydrolysation, déméthylation, désamination et ensuite conjugaison ( intervention du cytochrome P 450 ). Les molécules liposolubles sont ainsi de venues hydrosolubles et sont éliminées par les reins via le sang.Cette détoxification se fait essentiellement au niveau du foie, mais aussi au niveau des reins, des intestins, des poumons et de la peau.
- 2.6.3 La cavité
Ségrégation et accumulations de molécules et ions ( Ca++) et Voie de cheminement intracellulaire.
Après cheminement, les molécules sont emballées dans des vésicules formées à partir de la membrane du réticulum.
Certaines vésicules contiendrons des catalases et seront appelées péroxysomes.
D’autres contiendrons des hydrolases et porteront le nom de lysosomes.
Enfin, certaines dites de transition gagneront l’appareil de Golgi.
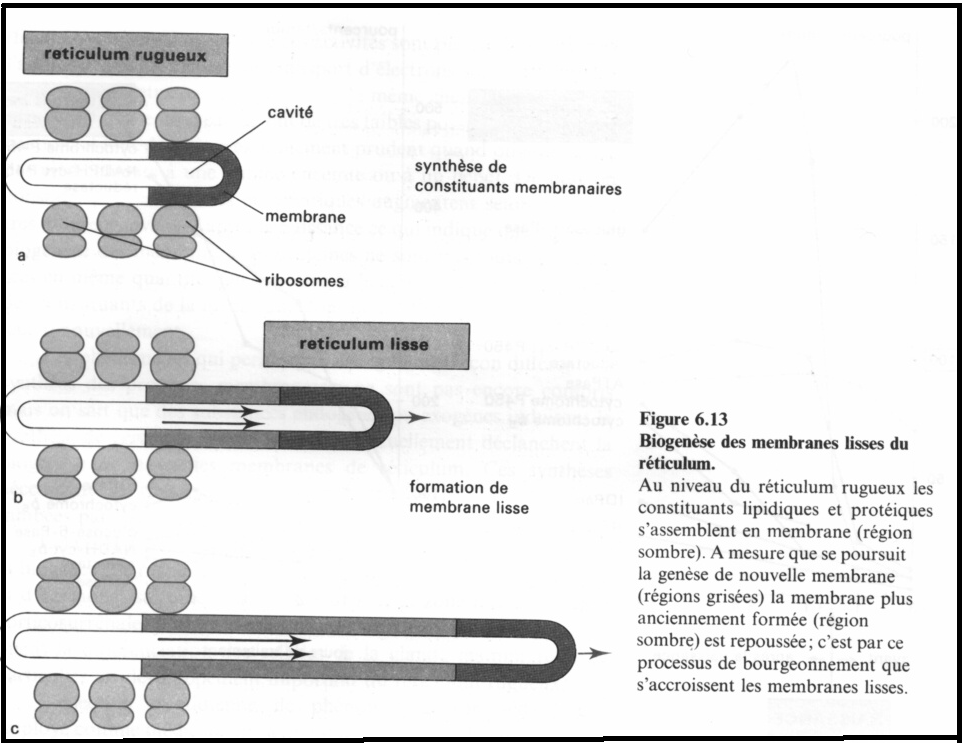
II.2.7 Biogénèse du réticulum endoplasmique et synthèse de membrane plasmique
Ce système de membranes hautement différencié , où se synthétisent les phospholipides et les protéines est à l’origine de la formation des membranes de la cellule.L’étude de la cytodifférenciation indique que l’apparition du REG précède celle du REL.Les
protéines et les phopholipides sont assemblés dans le REG puis transportés pour constituer le REL. La quantité de phospholipides synthétisés est réglée par la quantité de protéines capables de se fixer aux phospholipides.
Ce qui signifie que la quantité de nouvelles membranes synthétisées est déterminée par la quantité de protéines disponibles dans le REG où la liaison phospholipides – protéines s’effectue.La synthèse des membranes du REL pourrait aussi s’effectuer sur place sans qu’il y ait transfert de membranes .
Le document personnel suivant reprend et étend ce que nous avons vu jusqu'ici :
https://1drv.ms/p/s!Ao2vWE-NUWRIhA22cBZsDWMoGTP2?e=k28gEp
Date de dernière mise à jour : 16/06/2020
Ajouter un commentaire