

Biologie cellulaire 4
I.7.2.4 Reconnaissance des cellules entre elles
Toutes les cellules d’un même organisme portent sur leur membrane plasmique des protéines appelées antigéniques ou marques du soi . Ces protéines présentent un très grand polymorphisme car leur synthèse est placée sous le contrôle de gènes comportant de très nombreux allèles.
Il s’agit du complexe majeur d’histocompatibilité (CMH) qui chez l’homme porte le nom spécifique de système HLA.
Le rôle biologique actuellement proposé est la présentation de l’antigène ( classe I) au cours de la reconnaissance spécifique par les lymphocytes T ( classe II). Le grand degré de diversité explique pourquoi il est très difficile de trouver deux personnes non apparentées HLA identiques.
Les protéines du CMH constituent 1% des protéines membranaires.Grossièrement, les molécules du CMH humain ou HLA sont groupées en molécules de classe I ( toutes les cellules de l’organisme ) et de classe II ( macrophages, lymphocytes B). Ces deux types de molécules émergent à la surface des membranes cellulaires et n’induisent pas de réaction de la part des lymphocytes.
Par contre, ces sortes d’antennes présentent en quelque sorte toute protéine étrangère ou issue de cellules vieillissantes de manière à alerter le système immunitaire.
Structure des CMH ( de classes I et II )
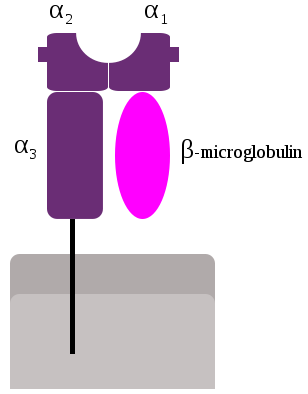
CMH de classe I : deux chaînes polypeptidiques α et β qui présentent toutes deux des domaines « immunoglobuline-like » et qui sont associées de manière non covalente
la chaîne α est polymorphe
la chaîne β est non polymorphe et elle est codée par un géne non présent dans le CMH
Les molécules du CMH I sont constituées de quatre parties caractéristiques :
une région de liaison au peptide antigénique
une région immunoglobuline-like est formée par les domaines β2m et α3 et est la région qui fixe le CD8
une région transmembranaire qui est unique, la chaîne β2m ne présentant pas de segment transmembranaire
une région intra-cytoplasmique qui également unique pour les mêmes raisons que pour la région transmembranaire
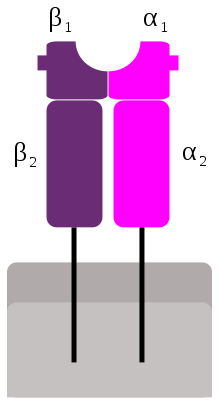
CMH de classe II : idem CMH de classe I
la chaîne α est polymorphe
la chaîne β est non polymorphe et codée par un gène présent dans le CMH (contrairement à la chaine β du CMH de classe I)
Les molécules du CMH II sont constituées de quatre parties caractéristiques :
une région de liaison au peptide antigénique est formée par les domaines α1 et β1 qui forment une cavité dans laquelle ira se loger le peptide antigénique
une région immunoglobuline like est formée par les domaines α2 et β2 est la région qui fixe le CD4
une région transmembranaire constituée de deux segments, un provenant de la chaîne α et l’autre de la chaîne β
une région intra-cytoplasmique est également constituée de deux segments pour les même raisons que la région transmembranaire
I.7.3 Les jonctions entre cellules
1.7.3.1 Les espaces intercellulaires
Les cellules d’un organisme sont séparées d’entre elles par des espaces intercellulaires interrompus par places par des complexes jonctionnels, elles s’engrènent par des interdigitations.
Les cellules n’adhèrent pas les unes aux autres sur toute leur longueur. Dans un épithélium prismatique simple, des fentes ou des fentes intercellulaires séparent les cellules. Ils sont obturés au niveau du pôle apical par une formation, le cadre cellulaire , structure destinée à empêcher la sortie du liquide intercellulaire et le passage de la surface de l’épithélium vers la fente.
L’espace intercellulaire a des dimensions relativement constantes. Il intervient dans la cohésion des cellules grâce à la présence d’un gel hydraté renfermant des protéines ( fibronectine, élastine, collagène).La présence de Mg++ et de Ca ++ est indispensable pour assurer cette cohésion.
I.7.3.2 Morphologie des zones de jonction intercellulaires.
Les types de jonctions intercellulaires sont nombreux. Les jonctions intercellulaires diffèrent par :
Leur forme
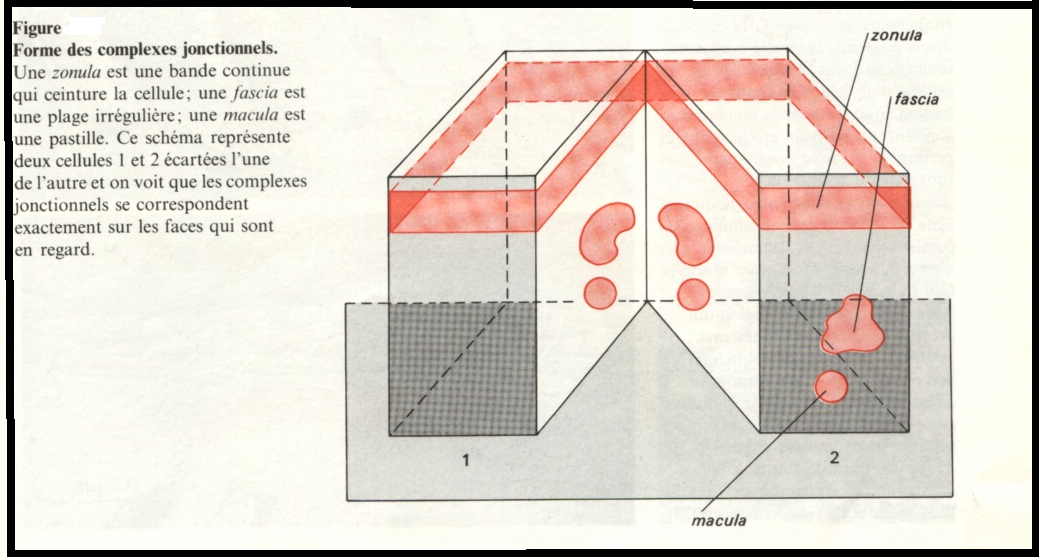
Le type zonula, correspondant à une jonction qui encercle complètement la cellule, s’observe essentiellement dans les épithéliums et plus particulièrement dans la zone apicale des épithéliums prismatiques simples.
Le type fascia , s’applique aux jonctions plus ou moins étendues à contour irrégulier.
Le type macula ou maculaire décrit des jonctions circulaires ou ovalaires.
Leur structure :
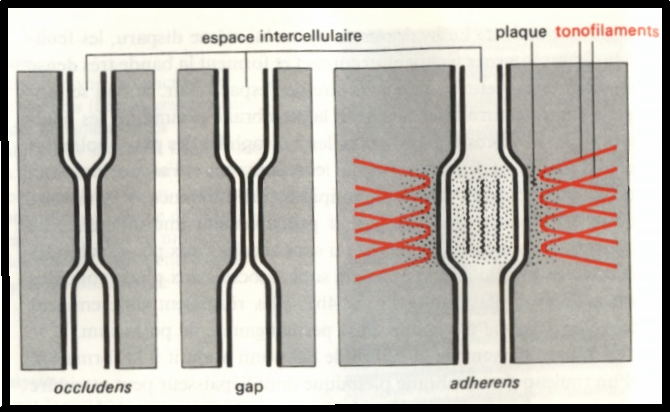
Type occludens, tigh junction , jonction serrée : dans ce type de jonction il y a disparition de l’espace intercellulaire par fusion des feuillets externes des membranes des deux cellules adjacentes.
Gap junctions, jonctions communiquantes. : L’espace intercellulaire y est très réduit ( 2 nanomètres ) .
Type adherens, desmosomes : L’espace intercellulaire est plus large ( 20 nanomètres) et rempli d’un matériel qui diffuse les électrons.

I.7.3.3 Ultrastructure des zones de jonction intercellulaires.
Les cellules dans un tissus sont donc séparées par un espace intercellulaire plus ou moins large , dans lequel circule un liquide interstitiel transportant les éléments nutritifs, les produits de sécrétion et les déchets. Dans certaines régions, les membranes cellulaires adhèrent l’une à l’autre ; ces zones de jonction intercellulaire
comprennent principalement :
Les tigh junctions
Les gap junctions
Les desmosomes
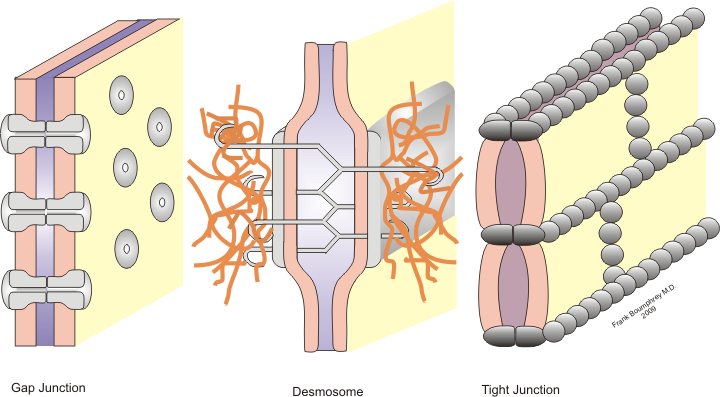
Les tigh junctions
a) Définition : Ce sont des régions spécialisées de la membrane où les feuillets externes établissent un contact si étroit qu’ils obturent complètement l’espace intercellulaire et empêchent le passage de toute substance .
b) Localisation : Ce type de jonction s’observe dans de nombreux tissus . Il unit les cellules endothéliales les unes aux autres, les entérocytes, les hépatocytes qui bordent un canalicule biliaire.Elles constituent une bande continue , ceinturant les cellules ( zonula
occludens au niveau de entérocytes.).
c) Structure
Sur une coupe perpendiculaire à leur plan, les tigh junctions montrent cinq feuillets :
Deux feuillets sombres, correspondant aux feuillets internes des membranes cellulaires .
Deux feuillets clairs situés de part et d’autre d’un feuillet médian, ces feuillets d’aspect clair sont les résidus accolés des feuillets externes de la membrane plasmique.
Un feuillet médian correspondant à la zone de fusion des feuillets externes des membranes cytoplasmiques. Cette fusion n’est pas continue, les techniques de cryodécapage démontrent qu’elle n’a lieu qu’au niveau de particules protéiques alignées irrégulièrement, formant une image en réseau de ces contacts cellulaires. Les points de fusion résultent d’une interpénétration des protéines globulaires qui se font face.
d) Fonctions
Cette jonction est occlusive, l’espace intercellulaire virtuel s’oppose au passage du liquide interstitiel.
Les échanges se font donc obligatoirement à travers les cellules et non par les espaces intercellulaires.Les zonula occludens sont constituées de plusieurs protéines transmembranaires dont les deux principaux représentants sont l'occludine et les membres de la famille des claudines. Ces protéines transmembranaires sont associées à d'autres protéines comme la ZO-1, la ZO-2, la ZO-3. La ZO-1 interagit avec la spectrine, elle-même reliée aux microfilaments d'actine du cytosquelette.
Les gap junctions ( jonctions communicantes ).
a) Définition
Les gap junctions sont des régions spécialisées des membranes de deux cellules adjacentes, qui se caractérisent essentiellement par un rapprochement des deux membranes.
b) Structure
Sur une coupe perpendiculaire à leur plan, les gap junctions sont formées de sept feuillets ( dans la mesure ou l’espace intercellulaire peut être assimilé à un feuillet) :
Les 3 feuillets de la membrane d’une des deux cellules ( un sombre, un clair ).
L’espace intercellulaire ( 2 à 3 nanomètres ).
Les trois feuillets de la membrane de la cellule voisine.
Les gap comprennent donc :
Quatre feuillets osmiophiles.
Trois feuillets osmiophobes.
Très souvent les deux feuillets externes sont si rapprochés qu’il est difficile en microscopie de faire la différence entre une gap et une tigh junction. Des techniques utilisant des traceurs des milieux intercellulaires sont alors requises.
c) Organisation moléculaire
Chacune des membranes cellulaires qui fait partie de la jonction gap ( membrane jonctionnelle) possède dans son épaisseur des protéines connues sous le nom de connexines. Les molécules de connexine s’associent en connexon ( hexamère de connexine).
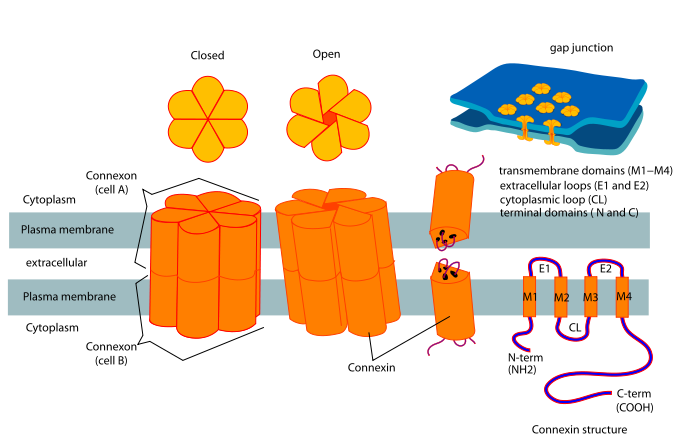
Chaque connexon d’une membrane est connecté à un connexon de la deuxième membrane jonctionnelle.Ces connexons ont un diamètre de 6 nanomètres et un canal central de deux nanomètres. Ils sont disposés selon un réseau à mailles hexagonales.
Les desmosomes maculaires
a) Définition
Ce sont les systèmes les plus complexes, les plus différentiés d’attache intercellulaire,distribués à des intervalles plus ou moins réguliers le long des limites cellulaires.
b) Forme
Ils sont ovalaires avec un diamètre de 400 à 500 nanomètres
c) Structure
Des coupes perpendiculaires au plan du desmosome révèlent :
que les membranes des cellules voisines ne fusionnent pas et n’établissent pas de contact direct,
que l’espace intercellulaire est à peu près de 20 nanomètres mais peut atteindre 50 nanomètres dans les structures les plus différentiées.
d) Les composants extracellulaires.
A) LE « COAT » ( glycocalyx contenant des oligosaccharides, des glycoprotéines et des glycolipides). Le coat à ce niveau est devenu plus compact, plus dense, il constitue le cément inter desmosomal.
B) LA LAMELLE CENTRALE OU LIGNE MEDIANE OU LIGNE DENSE CENTRALE.
v Localisation
Le centre de cet espace est occupé par une lamelle plus dense que la substance occupant cet espace ; elle est équidistante des membranes cellulaires . Elle est inconstante, lorsqu’elle existe elle peut apparaître comme une ligne continue ou comme une ligne interrompue à intervalles réguliers.
v Nature
Cette lame intercellulaire représenterait la région où les fibres et les particules du cell coat des deux cellules voisines s‘interpénètrent . Les particules de la ligne dense centrale sont reliées à des particules intramembranaires par un réseau de fins filaments ( filaments de lien transmembranaires).
C) LES COMPOSANTS CYTOPLASMIQUES.
v L’épaississement du feuillet interne. Les membranes cellulaires qui délimitent cet espace montrent un épaississement relativement important de leur feuillet interne.
v La plaque cytoplasmique.
Cette plaque dense et épaisse est séparée de la membrane par un espace clair peu important. Elle caractérise des desmosomes fortement différentiés. Il est à noter que cette plaque peut adhérer au feuillet interne. Elle a une structure lamellaire, les lames
étant parallèles à la membrane cellulaire, ces lamelles sont associées entre elles dans une matrice homogène de structure voisine à la kératohyaline ( la kératohyaline se présente sous forme de granules et est un précurseur de la kératine).La kératohyaline est un matériel protéique contenant beaucoup d’acides aminés soufrés.
v Les fibrilles intracellulaires ou tonofilaments
. Elles sont les constituants les plus importants des zones de jonctions très différentiées. Les tonofilaments d’un diamètre de 10 nanomètres traversent les plaques cytoplasmiques : ils se divisent en protofilaments qui s’insèrent sur des particules protéiques
volumineuses intramembranaires. Les tonofilaments sont des composants du cytosquelette classifiés parmi les filaments intermédiaires de cytokératine (voir plus loin ).
e) Localisation
Les desmosomes existent dans tous les épithélium, les endothélium et les mésothélium.
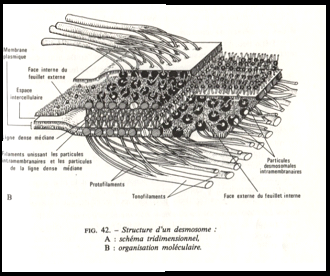
Le site d’accrochage intercellulaire nécessite la présence d’ions Ca++Les protéines qui assurent la jonction entre les deux membranes cellulaires adjacentes sont généralement du groupe des cadhérines, celles-ci s’unissent entre elles dans la zone intercellulaire et sont liées à des molécules d’actine par l’intermédiaire de protéines constituées de 3 sous unités, les caténines
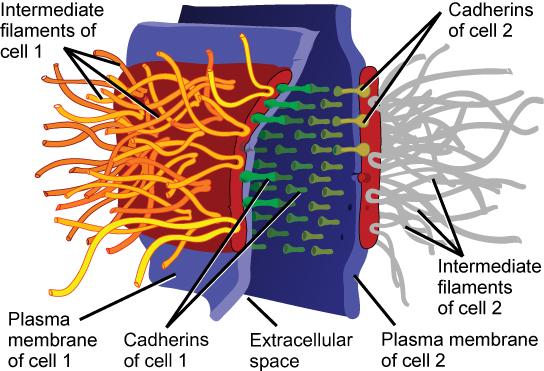
Desmosomes zonulaires
a) Définition
Les desmosomes zonulaires sont des jonctions continues qui circonscrivent le pôle apical des cellules prismatiques, juste au dessous des jonctions serrées.
b ) Structure
A . L’ESPACE INTERCELLULAIRE
Il a 15 à 20 nanomètres d’épaisseur, le cell coat n’y est pas condensé.
B. LES COMPOSANT INTRACELLULAIRES
De part et d’autre des membranes, existe une condensation cytoplasmique semblable aux plaques desmosomales, mais de nature chimique probablement différente.Des microfilaments d’actine des terminaisons de réseau microtrabéculaire s’insèrent sur ces plaques. L’interaction actine-protéines membranaires dépend d’une autre protéine, la vinculine (110000 daltons).
Les complexes de jonctions
a) Définition
Un complexe de jonction comprend les trois types de jonctions ( tigh, desmosome maculaire, desmosome zonulaire).
L’ensemble tigh-desmosome zonulaire forme le cadre visible en microscopie optique
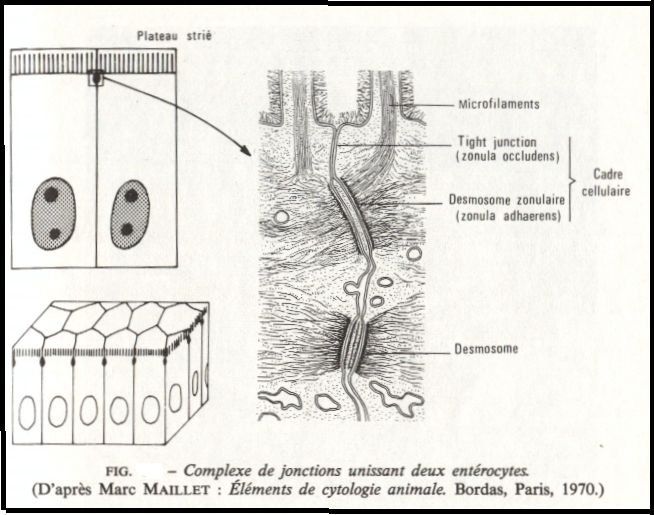
a) Localisation
Ces complexes de jonctions unissent le plus souvent les cellules prismatiques : Ils s’observent surtout dans l’épithélium intestinal , mais aussi dans d’autres types d’épithéliums comme les épithéliums glandulaires ( foie ).
Les hémidesmosomes
a) Définition
Ce sont des différentiations morphologiques assurant l’adhérence de l’épithélium à la lame basale et au chorion.
b) Structure
Celle-ci est variable selon le type d’épithélium considéré, mais quel qu’il soit, on peut, comme por les desmosomes ,distinguer :
- des composants cellulaires
- des composants extracellulaires.
La partie cellulaire comprend des structures sensiblement semblables à celles des desmosomes.
Les composants extra cellulaires se succèdent comme suit :
1) Une zone claire de 10 nanomètres de large occupée en son milieu par une ligne dense reliée à la membrane par un matériel filamenteux.
2) Une zone dense de 6 nanomètres d’épaisseur.
3) Une zone claire de 30 nanomètres d’épaisseur parcourue par de nombreux filaments ( microfibrilles de laminine).
4) La lame basale de 14 nanomètres d’épaisseur contenant de très nombreuses fibrilles qui prolongent celles venues de faisceaux du chorion. Ces faisceaux sont formés par des fibrilles dites d’ancrage, de 3 nanomètre de diamètre.
Ces hémidesmosomes sont de forme ovalaire, et de 200 nanomètres de diamètre.
Date de dernière mise à jour : 16/06/2020
Ajouter un commentaire