
EMBRYO DESCRIPTIVE 1
Dans ce cours, les aspect purement moléculaires du développement ne sont pas encore abordés ils feront l’objet d’une étude dans la rubrique biologie du développement.
Chapitre 1
L’ ORIGINE DE L’OEUF
1. Spermatogénèse-spermiogénèse
Voir biologie générale
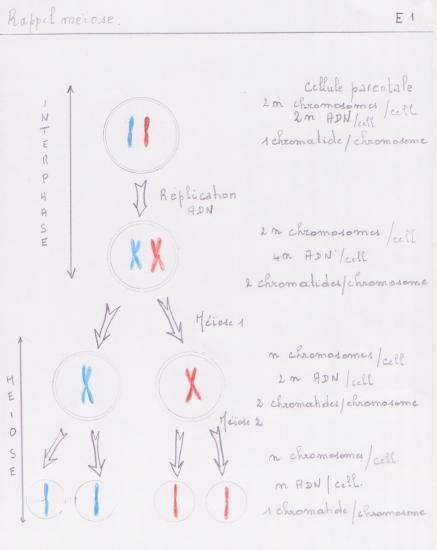
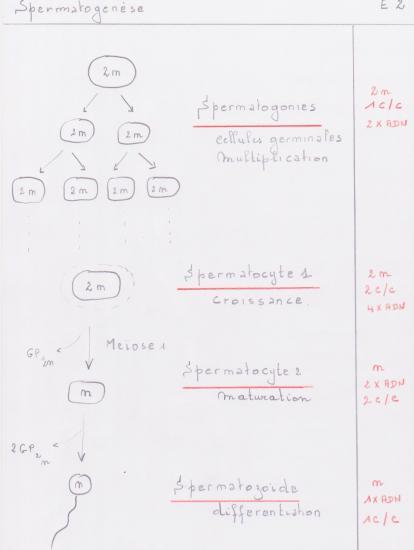
La spermatogenèse est le processus de production des spermatozoïdes, qui a lieu dans les gonades mâles ( tubes séminifères des testicules chez les Vertébrés ). Elle englobe les phénomènes qui, des spermatogonies, cellules souches diploïdes (2n chromosomes), aboutissent aux spermatozoïdes, gamètes mâles haploïdes (n chromosomes).
Schéma général-rappel

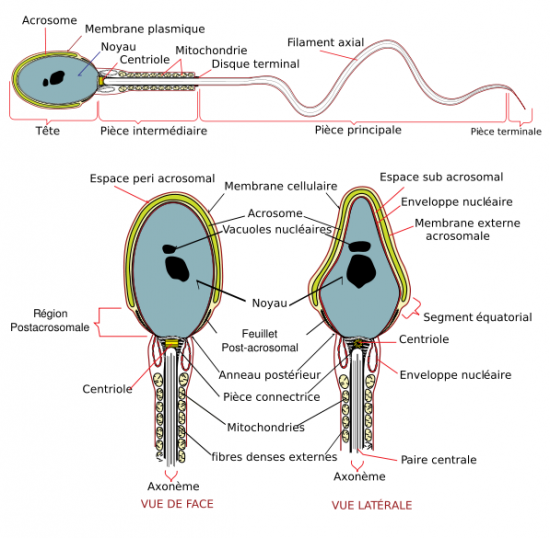
Structure d’un spermatozoïde de Vertébré aprés différentiation.
2.Ovogénèse
2.1 Phase de multiplication
Partant des cellules germinales, formation des ovogonies
2.2 Phase de croissance
Formation des ovocytes I ( 2n-4ADN-2 chromatides/chromosome)
Deux stades :
- a) La provitellogénèse
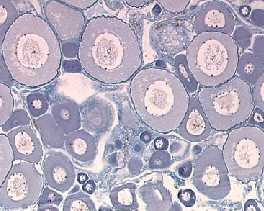
Ovocytes humains en pro-vitellogénèse
- Synthèse de beaucoup d’ARNr ( nucléole )
- Synthèses enzymatiques nombreuses
- Multiplication des mitochondries
La machinerie se met en place pour le développement futur
- b) La vitellogénèse
- Accumulation de substances nutritives : le "vitellus"
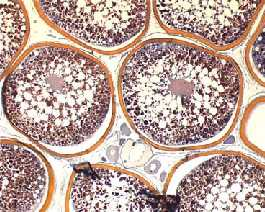
Ovocytes humains en pleine vitellogénèse

Follicule atrétique dans un ovaire, en pleine vitellogénèse
Le vitellus : protéines ( la phosphitine ), glucides, lipides.
Les plaquettes vitellines viendront soit des cellules nourricières de l’ovocyte I ( follicules primaire ) soit des cellules hépatiques.
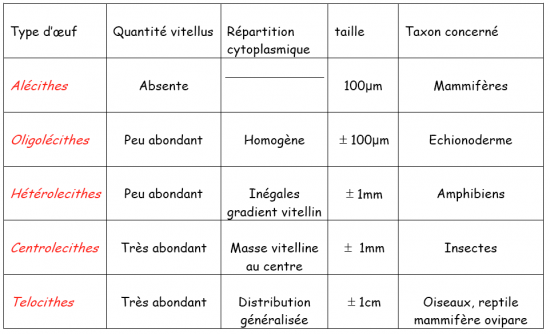
Suivant l’importance de la charge en vitellus, on classera les ovules de la façon suivante :

2.3 Phase de maturation
Par méïose, passage à l’ovocyte II puis à l’ovule
Au point de vue métabolique
Synthèse importante de protéines enzymatiques migrant vers le noyau qui gonfle, ces protéines serviront lors des segmentations de l’oeuf, mise en oeuvre future d’une quantité importante d’ADN.
Rq : - a) la progestérone intervient :
- dans le déclenchement de la division réductionnelle
- dans les processus métaboliques
- b) Chez les Vertébrés, la phase de maturation s’arrête à la métaphase de la mitose équationnelle ( entre ovocyte II et ovule ).
- c) Acquisition du pouvoir fécondant par le spermatozoïde
- au niveau épididyme et canal déférent : acquisition de la mobilité
- dépôt sur la membrane du spermatozoïde de glycoprotéines ( renforcement de la protection et de la stabilité de la membrane ).
- remaniement de l’acrosome , forme et structure ; anti-enzymes qui empêchent les enzymes acrosomiales d’exercer leurs activités à l’égard des cellules bordantes des voies génitales mâles et femelles . Ex : anti trypsine.
- émission d’un plasme séminal de protection phénomène de DECAPACITATION , masquage des marques du soi..
- au niveau des trompes de Fallope : phénomène de CAPACITATION , ou aptitude à féconder. des substances émises par les trompes vont provoquer un changement général des propriétés de membrane du spermatozoïde. La fusion des membranes de l’ovule et du spermatozoïde est dès lors possible.
3. La fécondation
Phénomène suivi sur oeuf d’oursin

3.1 L’activation
3.1.1 Les phénomènes morphologiques
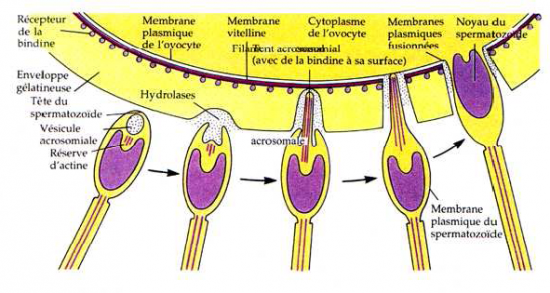
a) Spermatozoïde en contact avec la gangue gélatineuse de l’ovule
Digestion de cette gangue par les hydrolases de l’acrosome
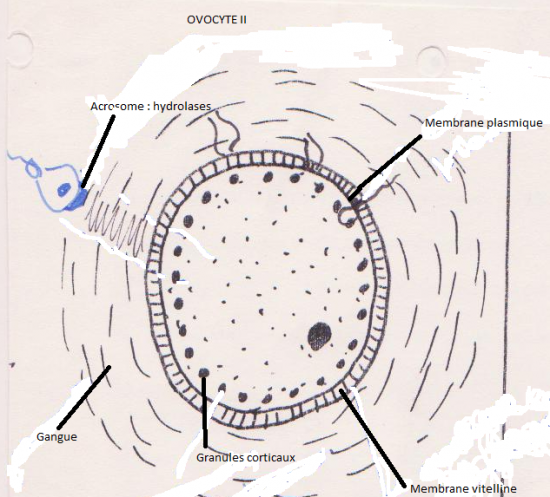
b) Spermatozoïde en contact avec les membranes vitelline et plasmique.
- tube acrosomial entre en contact avec la membrane vitelline qui est perforée ( hydrolases).
- fusion des membranes plasmique et formation d’un pont qui va en s’élargissant.
- sites spécifiques d’interaction ( protéines réceptrices ) : démasquage des antigènes par capacitation.

c) Monospermie
Règle : en condition de saturation, jusqu’à plus ou moins 1500 spermatozoïdes sur ovule ( redondance nécessaire ). Si pénétration de plusieurs spermatozoïdes, il y a polyspermie ( le nombre de chromosomes dépasserait la normale... ) , avortement précoce. Pour parer à cette éventualité : système de régulation de la pénétration.
d) Système de la régulation de la pénétration.
Il se compose de deux obstacles à la polyspermie
1° Mécanisme de blocage rapide mais incomplet
Quelques secondes après le contact entre gamètes, entrée de Na+ dans la cellule → variation de potentiel électrique comparable à la variation d’un potentiel d’action nerveuse.
2° Mécanisme de blocage lent mais complet
Plus ou moins 20 secondes.
Il s’agit de remaniements structuraux.
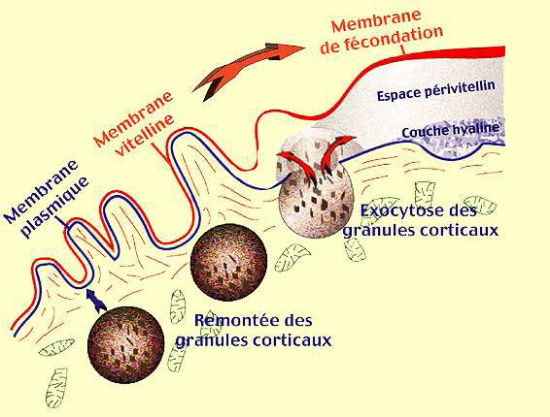
- les granules corticaux, au site d’interaction, vont fusionner avec la membrane plasmique ovocytaire; cette fusion va s’étendre à toute la surface de l’oeuf.
Les molécules contenues dans les granules vont jouer un rôle spécifique :
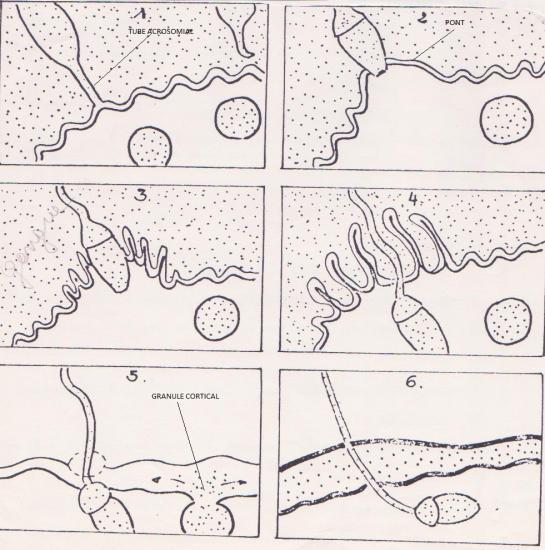
- Les deux enzymes présentes :
- Une modifie spécifiquement les protéines réceptrices de la membrane vitelline; résultat : les spermatozoïdes en surnombre se détachent .
- L’autre rompt les protéines de liaison qui relient les membranes vitelline et plasmique.
- Les substances colloïdales ( mucopolysaccharides ) :
Elles attirent par osmose de l’eau dans l’espace créé entre les deux feuillets membranaires. Le décollement s’accentue.
- Les protéines de structure :
Elles s’associent à la membrane vitelline qui est, de la sorte, renforcée . Cette enveloppe portera le nom de "membrane de fécondation".
Pendant ce temps, d’autres substances colloïdales provenant des granules corticaux recouvrent la surface externe de la membrane plasmique pour former la "membrane hyaline" de l’oeuf dont le rôle sera d’assurer ultérieurement la cohésion des cellules issues de l’oeuf.
Entre les deux membranes : le " liquide périvitellin ".
- Resserement des cellules périovocytaires
3.1.2 Les phénomènes physiologiques
a) avant la fécondation, l’ovule est au repos :
- fonctions réprimées : respiration, échanges cellulaires, synthèse de protéines, d’ARN, synthèse d’ADN bloquée.
b) au moment de la fécondation , reprise des activités.
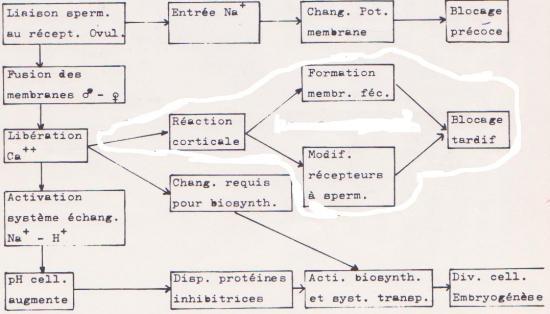
Le processus d’activation se divise en deux phases
1° Phase de changements précoces :
I ( interaction) + 3 ’’ : entrée de Na+ dans le cellule ( voir blocage précoce).
I + 20 ’’ : augmentation de la teneur en Ca++ à l’intérieur de la cellule ( réaction corticale commence ), entrée massive de Na+ . H+ libéré par l’ovocyte → pH augmente dans le cytoplasme.
I + 25 ’’ : activation d’une enzyme qui transforme la moitié des réserves cellulaires de NAD en NADP, ce qui implique que le métabolisme cellulaire devient plus réducteur , ce qui correspond à des synthèses accrues
I + 40 ’’ : augmentation de la consommation d’O2 par l’ovule. Glucose 6P déshydrogénase , en zyme requise dans le métabolisme des glucides ( glycolyse ) libérée dans le cytoplasme.
2° Phase de changements tardifs :
I + 5 ’ : réalisation des biosynthèses essentielles pour le développement de l’embryon-vitesse des synthèses de protéines augmentée-activation des systèmes de transport de la cellule apportant des acides aminés, des phosphates, des nucléosides.
I + 25 ’ : premier cycle de synthèse d’ADN.
Explications :
- Ca++ induit la fusion des granules corticaux avec la membrane plasmique ( flash calcique ).
- Entrée de Na+ et sortie de H+ : échange 1 Na+ pour 1 H+
Cette libération de protons par l’oeuf rendrait le cytoplasme plus alcalin et de ce fait, plus compatible avec une activation des réactions de biosynthèse dansq l’oeuf.
En effet, une augmentation de pH provoquerait une destruction d’inhibiteurs bloquant les enzymes et protéines ribosomiales.
En résumé :
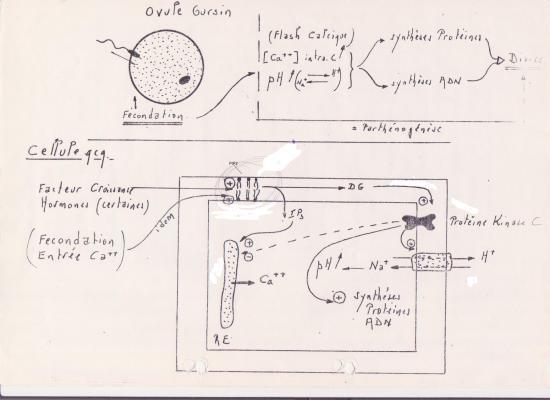
L’inositol triphosphate (1,4,5-triphosphate ou IP3) et les diglycérides sont le résultat de l’hydrolyse d’un phospholipide membranaire le phosphatidyl-inositol-diphosphate (PIP2),ce sont des seconds messagers membranaires, Cette réaction est commandée par plusieurs types de récepteurs types de récepteurs, liés aux phospholipases C. Les diglycérides membranaires agissent en activant les protéines kinases C. L’inositol triphosphate est un second messager, qui facilite la libération du Calcium contenu dans les citernes du reticulum endoplasmique.
3.1.3 Les phénomènes cytoplasmiques
L’activation se traduit également par des remaniements cytoplasmiques.
Modification dans la répartition des granules pigmentaires ayant pour résultat l’apparition du croissant gris à l’opposé du point d’entrée du spermatozoïde
Position du croissant gris :
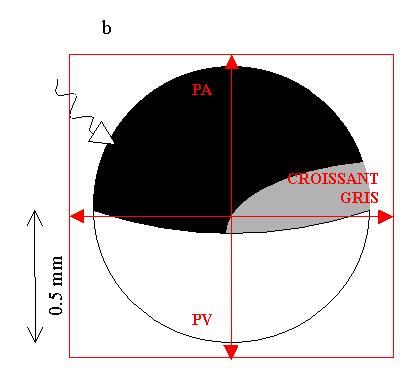
- Etablissement d’une polarité dorso-ventrale ( PA-PV) et d’une symétrie bilatérale.
Cette zone du croissant gris est importante car c’est à ce niveau qu’apparaîtront successivement :
1) Le blastopore
2) Les organes axiaux de l’embryon : somites, corde et tube nerveux.
3.2 L’amphimixie
L’amphimixie est la fusion des 2 lots haploïdes de chromosomes. Elle ne survient qu’après la formation de 2 pro-noyaux (pronucléus ou pronuclei). Elle va être immédiatement suivie de la première division cellulaire de l’oeuf, aboutissant à la formation des deux premiers blastomères.
La formation des pro-noyaux (pronucléus)
Les 2 pro-noyaux mâle et femelle apparaissent simultanément, environ 10 heures après la fusion des gamètes.
Les pronucléus sont de volumineuses structures sphériques, de 20 à 30 µm de diamètre.
* Le pro-noyau femelle
A la suite de la méiose, une enveloppe nucléaire se reforme autour du lot haploïde des chromosomes maternels. Les chromosomes se décondensent et des nucléoles apparaissent immédiatement.
* Le pro-noyau mâle
Au cours de la dégradation du spermatozoïde, le noyau spermatique perd temporairement son enveloppe.
La migration des pro-noyaux
A proximité du noyau du spermatozoïde se trouve le centriole proximal du spermatozoïde. Il induit la formation d’un appareil achromatique sans centriole (particularité des Mammifères). c’est un réseau étoilé de microtubules, appelé le spermaster.
Le spermaster tracte le pronucléus mâle vers le centre de l’ovotide (20 h de migration).
Le pro-noyau femelle se rapproche également du centre de l’ovotide par un processus analogue (le centriole femelle a été éliminé au cours de la deuxième division de méiose).
Durant cette période de migration :
- Les synthèse protéiques sont actives dans le cytoplasme
- Il y a réplication de l’ADN dans chacun des pro-noyaux, induite par des facteurs cytoplasmiques.
A la fin de la migration les chromosomes sont bichromatidiens. L’oeuf est donc temporairement pseudo-tétraploïde (avec 4n ADN) avant la 1ère division de segmentation, comme avant n’importe quelle mitose.
chromosomes paternels se décondensent. Durant cette étape, les protamines liées à l’A.D.N. sont remplacées par des histones synthétisées dans le cytoplasme ovocytaire. Des nucléoles réapparaissent.
L’ensemble forme une masse volumineuse (500 fois le volume du noyau du spermatozoïde). Une enveloppe nucléaire se reforme alors en périphérie. L’aspect est alors le même que celui du pro-noyau femelle.

Suite embryologie descriptive 2
Sous-pages :
Date de dernière mise à jour : 02/04/2018
Commentaires
-
 D'accord, merci.
D'accord, merci. -
 Merci de votre réponse rapide! Cependant il y a toujours qqchose que je ne comprend pas. Quand vous dites"trace au niveau du génotype", voulez vous dire qu'une petite quantité de vitellus reste tout de meme présent au pole végétatif, définissant alors une polarisation précoce comme les lécithe, et expliquant qu'il y ait une vitellogénese chez les ovocytes humains (j'ai vu sur un autre site que le zygote humain serait microlécithe plutot qu'alécithe, (source non certaine)) ; ou alors que des protéines spéciales s'orienterait en définissant les deux poles selon un mécanisme différent du phénomene de gravité observé avec le vitellus?Voilà la dernière question que vous vous posez et la bonne et effectivement l'œuf humain est microlécithe comme vous le verrez sur la page suivante de ce même site ( microphotographie de la vitellogénèse humaine :http://www.vdsciences.com/pages/sciences-biologiques/biologie-animale/embryologie-descriptive/embryologie-descriptive.html
Merci de votre réponse rapide! Cependant il y a toujours qqchose que je ne comprend pas. Quand vous dites"trace au niveau du génotype", voulez vous dire qu'une petite quantité de vitellus reste tout de meme présent au pole végétatif, définissant alors une polarisation précoce comme les lécithe, et expliquant qu'il y ait une vitellogénese chez les ovocytes humains (j'ai vu sur un autre site que le zygote humain serait microlécithe plutot qu'alécithe, (source non certaine)) ; ou alors que des protéines spéciales s'orienterait en définissant les deux poles selon un mécanisme différent du phénomene de gravité observé avec le vitellus?Voilà la dernière question que vous vous posez et la bonne et effectivement l'œuf humain est microlécithe comme vous le verrez sur la page suivante de ce même site ( microphotographie de la vitellogénèse humaine :http://www.vdsciences.com/pages/sciences-biologiques/biologie-animale/embryologie-descriptive/embryologie-descriptive.html -
Bonjour Monsieur/Madame. Je tiens à vous remercier de votre cours détaillé et bien illustré. Comme vous avez l'air d'avoir une bonne connaissance du sujet, j'aimerais vous poser une question, si possible, sur le zygote humain.
Celle ci sera peut etre bete ou mal formulée mais je n'ai pas réussi à trouver jusqu'ici. En fait je ne sais pas dans quelle mesure la cellule oeuf humaine présente un pole animal et végétal car l'oeuf est allécithe donc il ne présente pas de vitellus. Hors le pole végétal se différencie du pole animal (uniquement?) par la présence de vitellus. Comment donc faire la différence entre les deux poles? Merci d'avance?C'est une très bonne question ! je vais vous donner quelques éléments de réflexion : Tenez compte du phénomène évolutif, les cellules vitellines se sont en quelques sortes vidées, mais il reste une trace au niveau du génotype qui fait qu'elles restent " végétatives " C'est l'amphimixie qui déclenche cette différentiation , mais les déterminants sont déjà présents au niveau de l'œuf non fécondé. Si vous regardez une "blastula" , "anse vers le haut " vous verrez que les cellules de la "anse" sont plus petites que les cellules sous le plancher du blastocoele. , l'axe horizontal de cette blastula n'est pas un axe de symétrie, l'orientation de cette blastula ou du moins des contenus cytoplasmiques cellulaires est déterminée par la gravité et la gravité est un déterminant de la différenciation pôle végétatif, pôle animal........., bonne réflexion ! étant donné que dès le départ l'œuf n'est pas homogène au niveau densité, ce qui implique l'existence de zones différentes impliquant des phénomènes épigénétiques différents
Ajouter un commentaire