
EMBRYO DESCRIPTIVE 5
2.4.5 Gastrulation des Mammifères
Généralités
Oeuf alécithe; segmentation holoblastique rotationnelle.
Bien que les mouvements morphogénétiques de la gastrulation des reptiles et oiseaux aient évolué en adaptation à des oeufs riches en vitellus, ils ont été retenus chez les mammifères.
L’épiblaste embryonnaire de la blastula mammalienne est similaire à l’épiblaste aviaire. À son bout postérieur un épaississement produira la ligne primitive et le noeud primitif, qui se creuseront en sillon primitif et gouttière primitive au travers lesquels migreront les précurseurs endodermiques et mésodermiques. Ce processus est semblable à celui retrouvé chez les oiseaux.
Revenons brièvement à ces notions, nous limitant aux Vertébrés. On réalise que le premier axe à se manifester est l’axe dorso-ventral. Cet axe se trouve à définir quelle est la face dorsale de l’embryon et quelle est sa face ventrale. Une ligne coupant cet axe sépare l’embryon en 2 moitiés, gauche-droite, et établit la symétrie bilatérale. Toutefois, quand on examine le développement rétrospectivement, on réalise que l’axe dorso-ventral, la symétrie bilatérale, était mis en évidence plus tôt; il a été déterminé plus tôt qu’il n’apparaît. En effet, le point d’entrée du spermatozoïde a amené un réaménagement cytoplasmique important. Ce point correspond au pôle ventral de l’embryon et celui diamétralement opposé au pôle dorsal. Cependant, le spermatozoïde agissait sur un cytoplasme déjà polarisé, qui imposait des contraintes à son entrée, et la polarité de l’oeuf est ultimement à l’origine de cet axe. Le pôle animal sera la face dorsale de l’embryon et le pôle végétatif la face ventrale. Nous avons vu que dans les oeufs non vitellins d’autres indices dévoilent une polarité intrinsèque. Les bases moléculaires et génétiques de cette polarité font actuellement l’objet de nombreuses investigations.
En considérant le développement rétrospectivement, l’axe antéro-postérieur se manifeste dès le début de la gastrulation. Le point de départ de la gastrulation: blastopore ou noeud primitif, correspond à l’emplacement du futur anus (le pôle postérieur); l’autre bout de l’embryon est inévitablement la future tête. Ici aussi la polarité a été déterminée plus tôt qu’elle n’apparaît. On le sait particulièrement des amphibiens chez qui la localisation du croissant gris, apparaissant à la fécondation, signale le point de départ de la gastrulation. Les bases moléculaires et génétiques de cette polarité font également l’objet de recherches intensives. Elle implique probablement un gradient de molécules cytoplasmiques dans l’oeuf, avant ou après la fécondation.
Il faut réaliser que les deux polarités sont dissociables. L’existence d’un axe, que ce soit le dorso-ventral ou l’antéro-postérieur, n’amène pas celle de l’autre.
Une autre forme de polarité ou d’axe existe; il s’agit de la latéralisation. L’axe dorso-ventral, ou la symétrie bilatérale, a amené la création de 2 moitiés dont on ne sait toutefois pas encore laquelle est la gauche et laquelle la droite tant que l’axe antéro-postérieur n’est pas établi. Ceci accompli, les moitiés gauche et droite sont en principe identiques, sauf qu’une est l’image miroir de l’autre. Cependant, on sait pertinemment, avant même d’étudier l’organogenèse, que tout dans le vertébré n’est pas pair. Qu’est-ce qui détermine que des organes uniques se localisent à gauche ou à droite? Il semble que la matrice extracellulaire joue un rôle dans cette détermination.
Modalités chez les Mammifères
I\ Protothériens ou Monotrèmes (mammifères inférieurs).
Exemple : l’ornithorynque.
Il y a des ovocytes riches en réserves, s’entourant d’albumen au niveau de l’oviducte puis d’une coque cornée contenant des nodules calcaires. Seule l’ovaire gauche est fonctionnel. L’œuf est pondu par la femelle dans un terrier.
Chez les échinides (ils ressemblent à des petits hérissons et sont proches des marsupiaux), il y a incubation des œufs dans une poche ventrale abdominale dans laquelle ils sont reçus au sortir du cloaque. Les deux ovaires sont fonctionnels et l’œuf pondu correspond à un embryon de poulet à 40 heures d’incubation.
On distingue trois périodes chez ces espèces :
- Une période intra-utérine (brève).
- Une période d’incubation.
- Une période qui suit l’éclosion et où commence la lactation.
II\ Les métathériens (les marsupiaux).
La gestation est de courte durée, l’embryon se développe dans l’utérus où il effectue son organogenèse. Il y a formation d’un placenta très rudimentaire. Quand le jeune est mis au monde, il gagne la poche marsupiale où il achèvera sa croissance.
III\ Les euthériens.
La gestation va atteindre son maximum de compétence. L’œuf est dépourvu de réserve (alécithes). Le développement va dépendre de substances nutritives apportées par la mère grâce à un organe d’origine fœto-maternelle qu’est le placenta.
IV\ Physiologie de la gestation.
L’œuf ne peut s’implanter dans la muqueuse utérine que si celle-ci est préalablement préparée par l’action synergique des œstrogènes puis de la progestérone.
L’ovariectomie précoce empêche la nidation. Or, quand la gestation est installée, on distingue deux types d’espèces :
- Espèce où l’ovaire est indispensable jusqu’au terme (exemple : le lapin, le hamster).
- Espèce où l’ovaire n’est plus nécessaire dans la deuxième partie de la grossesse (femme, cobaye, chien, chat) : le placenta prend le relais hormonal et sécrète une gonadotrophine agissant sur le corps jaune cyclique qui devient ainsi gestatif. La grossesse se résume par un équilibre entre œstrogènes et progestérone, sans oublier les interactions hypophyse, hypothalamus.
V\ L’ovoimplantation.
Les œufs alécithes (de 100 à 150 µm de diamètre).
Dans les douze heures qui suivent la ponte, l’ovocyte II doit être fécondé, sinon il dégénère. La première mitose a lieu vers la trentième heure. La segmentation est asynchrone. Au bout de quatre jours, on obtient un stade morula, formé de douze à seize cellules. Le transit tubaire est favorisé par un flux péritonéo-utérin en péristaltisme de l’oviducte et de l’action des cils.
La morula arrive dans l’utérus où elle atteint un stade blastocyste (vers le 6ème jour). Ce blastocyste est formé de deux parties :
• des cellules aplaties à la périphérie
• un amas cellulaire à l’un des pôles constituant le bouton embryonnaire.
Le blastocyste peut demeurer libre dans la cavité utérine pendant un temps variable (jusqu’à six jours chez la femme).
VI\ Les différents types de nidation.
A\ L’implantation centrale.
Dans ce cas, la muqueuse utérine est peu transformée. Le blastocyste reste dans la lumière de l’utérus et s’y développe. Ses parois viennent alors s’accoler à celles de l’utérus. L’endomètre est peu transformé. C’est un phénomène que l’on observe chez les carnivores, les ruminants, les suidés et les chéiroptères.
B\ L’implantation excentrée.
Le blastocyste s’insinue entre les plis de la muqueuse utérine qui est fortement transformée et qui ressemble à une véritable dentelle. La transformation de l’endomètre se fera grâce à des hormones. Le blastocyste détruit partiellement la muqueuse utérine, se refermant sur lui-même pour former une chambre close, sans communication avec la lumière utérine. Cette implantation a lieu chez le lapin, le rat et la souris.
C\ L’implantation interstitielle.
Le blastocyste pénètre dans l’épaisseur de l’endomètre et s’y développe. C’est le cas des primates, de l’homme. La grossesse ectopique (extra-utérine) est fréquente chez la femelle, dans la trompe, l’abdomen ou sur un ovaire.
La gastrulation chez l’homme
La gastrulation se déroule en deux temps, il y a différenciation rapide des annexes embryonnaires ( pas de réserves vitellines )
Partons du blastocyste au moment où il entre en contact avec l’endomètre utérin.
1ère étape
1) F + 8 JOURS
Formation de l’entoblaste et de l’ectoblaste
Au niveau du bouton embryonnaire, l’ectoblaste se creuse en une cavité, la cavité amniotique

Le toit de cette cavité est tapissé par les amnioblastes.
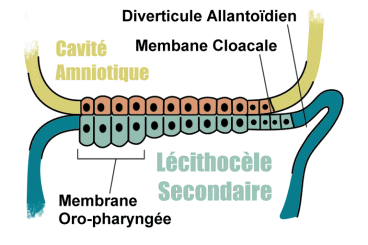
Entre les deux cavités ainsi formées, le disque embryonnaire est constitué par deux couches de cellules: l’entoblaste, vers la cavité du blastocyste ou lécithocèle primaire, et l’ectoblaste vers la cavité amniotique.
Le mésenchyme primaire, premier état du mésoblaste extra-embryonnaire, apparaît dans le blastocyste. Il est constitué de cellules qui proviennent du cytotrophoblaste. Ces cellules sont étoilées, anastomosées les unes aux autres. Les premières cellules formées s’organisent en une nappe délicate, la membrane de Heuser qui tapisse la surface interne de la cavité du blastocyste, hormis la région occupée par l’entoblaste. Progressivement, l’ensemble constitué par l’amnios, la plaque embryonnaire et la membrane de Heuser se décolle du cytotrophoblaste en formant un espace. Cet espace est occupé d’emblée par un réseau lâche de cellules du mésenchyme primaire.
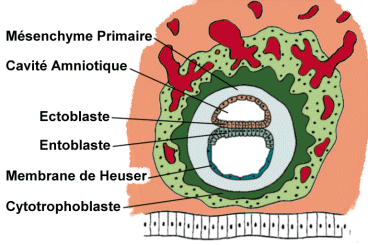
Le développement de l’entoblaste conduit à la constitution du lécithocèle secondaire, avec un phénomène de striction du lécithocèle primaire. Le développement du mésenchyme extra-embryonnaire produit la formation du coelome externe ( extra-embryonnaire ). Ce qui persiste du lécithocèle primaire, bordé par la membrane de Heuser forme un reliquat: le kyste exocoelomique.
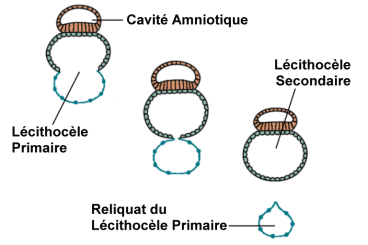
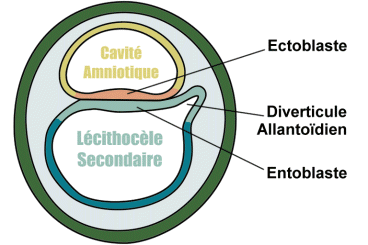
Dans la région caudale du lécithocèle secondaire apparaît une évagination dans le pédicule embryonnaire: le diverticule allantoïdien, de nature endoblastique.
Pendant ce temps, le trophoblaste se divise en deux assises cellulaires :
1) Le syncitiotrophoblaste
Au contact de la muqueuse, les cellules trophoblastiques émettent des villosités constituées de cellules plurinucléées qui envahissent les tissus maternel.
Au fur et à mesure de l’infiltration, les vaisseaux sanguins maternels sont érodés , et il se forme des lacunes remplies de sang ( action de protéases, d’enzymes lytiques)

3. Le cytotrophoblaste
Prolifère et émet des villosités qui finalement pénètrent dans les lacunes sanguines du syncitiotrophoblaste.
Ce sont ces microvillosités qui plus tard seront irriguées par les vaisseaux sanguins du fœtus.
Voir la suite dans le chapitre sur les annexes embryonnaires.
2) F + 10 JOURS
Le mésenchyme primaire ou extra-embryonnaire, qui est la forme primaire du mésoblaste extra-embryonnaire apparaît.
Ce mésenchyme proviendrait du cytotrophoblaste.
Croissance des villosités du trophoblaste
Décollement de la cavité amniotique par croissance du trophoblaste
Position de l’entoblaste dans le blastocoele.
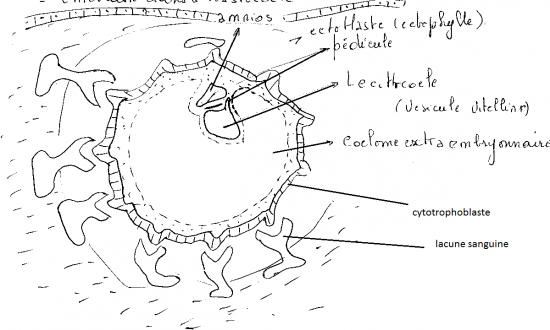
3) F + 14 JOURS
Fermeture de l’entoblaste complète : formation du lecithocoele encore appelé vésicule vitelline.
Délamination du mésenchyme primaire : apparition du coelome extra-embryonnaire ; formation du pédicule embryonnaire
2ème étape gastrulation proprement dite
Mise en place du cordomésoblaste
La gastrulation constitue l’évènement majeur de la troisième semaine.
Il s’agit d’un mécanisme complexe, programmé dans le temps et dans l’espace, associant prolifération et migration cellulaires à partir de l’épiblaste.
La gastrulation détermine l’orientation future de l’embryon et sa symétrisation ; elle met en place les 3 feuillets fondamentaux (ectoblaste, entoblaste et mesoblaste) de l’embryon qui devient alors tridermique. En outre la gastrulation se poursuit par la mise en place de la chorde et par la neurulation. Simultanément, et dès la mise en place de la chorde, la mésoblaste intra-embryonnaire va se segmenter dans le sens cranio caudal par un processus de métamérisation.
F + 16 JOURS
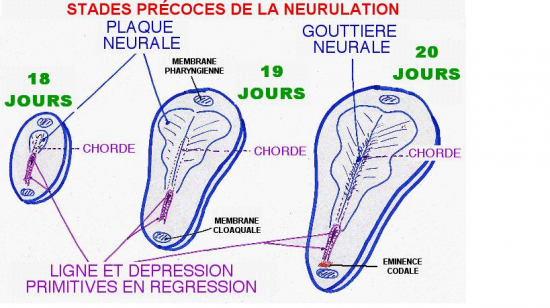
Processus analogue à celui vu pour l’œuf d’oiseau : Apparition d’une ligne primitive (LP) se terminant par le nœud de Hensen
Dès le 15ème jour, une structure linéaire, la future ligne primitive, apparait sur le versant épiblastique du disque embryonnaire dans la zone qui constituera ultérieurement la partie caudale de l’embryon. Son orientation prédétermine déjà l’axe crânio-caudal de l’embryon.
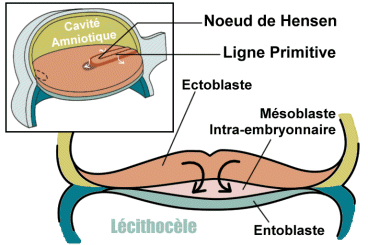
La ligne primitive va rapidement se creuser pour former une dépression dont l’extrémité proche du centre du disque embryonnaire se renfle circulairement et se creuse en son centre pour former le noeud de Hensen (ou noeud primitif ou dépression primitive).
La ligne primitive est alors le siège d’importantes migrations cellulaires : les cellules épiblastiques avoisinantes vont migrer, glisser dans la dépression primitive pour venir s’enfouir sous l’épiblaste.
Mais le déterminisme de ces migrations cellulaires obéit à une programmation génétiquement régulée.
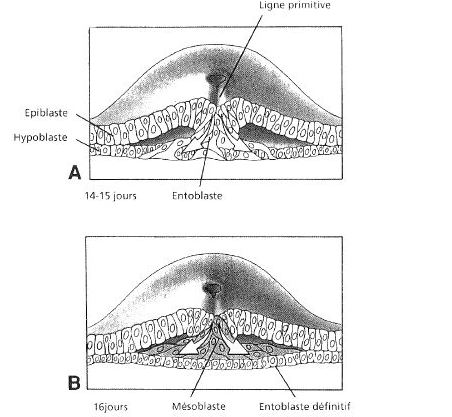
- Une première poussée cellulaire au travers de la ligne primitive se déplace jusqu’à l’hypoblaste. Les cellules hypoblastiques situées dans la zone du disque embryonnaire sont alors repoussées par ce nouveau contingent d’origine épiblastique. Ainsi se crée le feuillet entoblastique définitif qui remplace l’hypoblaste (appelé par certains entoblaste primaire) dans la zone du disque embryonnaire.
- Une deuxième composante cellulaire proliférant à partir de la ligne primitive aboutit à la formation de cellules qui migrent et s’insinuent entre épiblaste et entoblaste, latéralement à la ligne primitive mais aussi cranialement et caudalement, pour former un 3ème feuillet : le mésoblaste intra-embryonnaire.
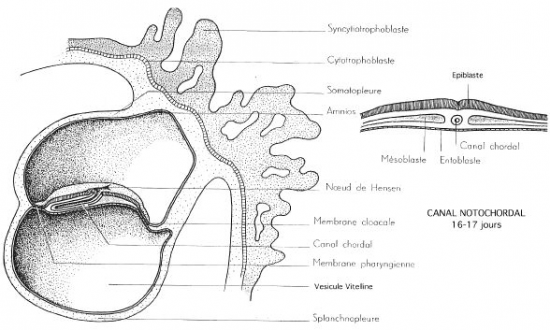
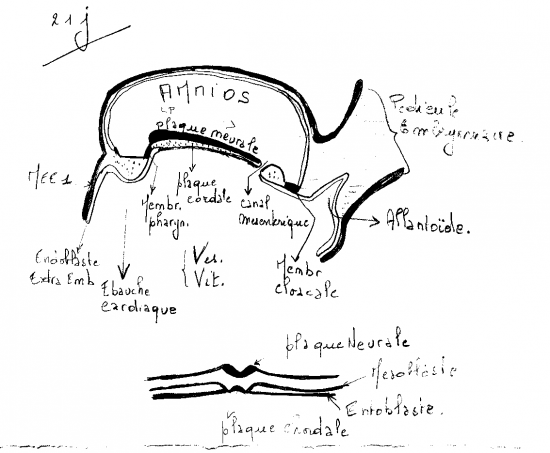
Dans deux zones strictement limitées, à l’extrémité craniale et à l’extrémité caudale du disque embryonnaire, l’épiblaste reste intimement accolé à l’entoblaste. Ces deux zones correspondent respectivement à la membrane pharyngienne et à la membrane cloacale.
F + 20 JOURS
Fin de gastrulation, voir coupes sagittales et transversales
Le nœud de Hensen recule .
Immédiatement en avant de la membrane cloacale, le mésoderme reste plus compact et forme l’éminence caudale. Il en est de même immédiatement en arrière de la membrane pharyngienne où on observe une masse mésoblastique compacte, la plaque préchordale .
La gastrulation (correspondant aux migration cellulaires que nous venons de décrire à partir de la ligne primitive) est une étape essentielle de l’embryogénèse dont les conséquences sont multiples :
a) Les 3 feuillets définitifs de l’embryon, ectoblaste, mésoblaste, entoblaste dérivent tous d’un même feuillet primaire : l’épiblaste.
b) La ligne primitive détermine l’orientation axiale cranio-caudale, de l’embryon et sa symétrie bilatérale.
c) Le déterminisme de la gastrulation est antérieur à la mise en place de la ligne primitive. Avant même la gastrulation, il existe des territoires présomptifs sur l’épiblaste correspondant à des groupes de cellules qui vont spécifiquement fournir le mésoblaste, l’entoblaste, la chorde après migration dans la ligne primitive ou le noeud de Hensen. Le territoire présomptif du neurectoderme est lui-même déjà prédéterminé.
Par inductions successives, les sous-groupes cellulaires prédéterminés dans les territoires présomptifs seront les précurseurs de l’ensemble des tissus et ébauches de l’organisme (lignage cellulaire).
C’est dire l’importance que revêt le mécanisme de la gastrulation et sa régulation.
Complément : détails sur la mise en place de la corde
Vers la fin de la deuxième semaine le germe dermique est compris entre deux cavités : la cavité amniotique du coté dorsale et la cavité du lecithocèle secondaire du coté ventrale.
La gastrulation se déroule entre les 16ème et les 22ème jours du développement embryonnaire Elle correspond à la mise en place d’un germe tridermique(ectoblaste, chordo-mésoblaste, endoblaste dit aussi entoblaste) à partir d’un germe dermique
Au début de la troisième semaine le disque embryonnaire se caractérise comme suit :
une augmentation de taille (2 mm environ ).
un changement de forme pour devenir ovalaire avec une région céphalique plus large que la région caudale.
un épaississement de l’éctophylle dans sa région caudale.
I- Formation de la ligne primitive et du nœud de Hensen
Vers le 16ème jour se dessine dans la région caudale de l’éctophylle, un sillon longitudinal médian : c’est la ligne primitive qui croit en direction du centre du disque. Sa croissance s’achève vers le 17ème jour par la mise en place du nœud de Hensen du coté craniale du sillon.
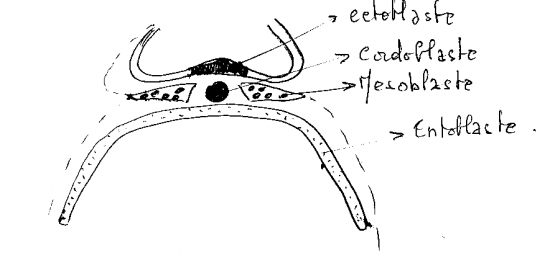
II- Mise en place du chordo-mésoblaste
A fin d’éviter la complication de l’exposé, les mise en place du mésoblaste embryonnaire et du matériel chordale seront traitées séparément malgré leur concomitance.
A- Mise en place du mésoblaste intra-embryonnaire
entre le 17ème et le 18ème jour toutes les cellules éctophylliques à potentialité mésoblastique pénètrent en profondeur, a travers la ligne primitive, pour s’insinuer en nappe entre l’éctophylle et l’entophylle excepté deux régions ou les deux feuillets suscités demeure accolée :
l’une céphalique qui constitue la membrane pharyngienne(futur bouche).
l’autre caudale qui donne la membrane cloacale(futur anus).
B- Mise en place du matériel chordal
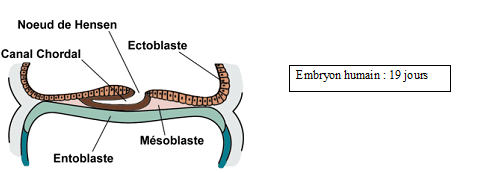
a) Stade canal chordal
Entre les 17ème jour et 18ème jours les cellules éctophylliques à potentialité chordale pénètrent à travers le nœud de Hensen obliquement mais axialement vers l’avant : c’est le prolongement céphalique pour former le canal chordale.
b) Stade canal chordal fissuré
vers le 19ème jour, un processus de fissuration longitudinal survient en plusieurs points de la ligne de soudure entre le toit de lecithocèle secondaire (endoblaste) et le plancher du canal chordale Suit à ce phénomène la cavité amniotique communique avec le lecithocèle secondaire par l’intermédiaire du nœud de Hensen.
c) Stade gouttière chordal renversée
vers le 20ème jour, la fissuration intéressant à la fois le toit du lecithocèle secondaire et le plancher du canal chordale fissurée s’est étendu à toute la largeur de ce dernier de sorte que le matériel chordale a pris temporairement la forme d’une gouttière renversée.
A ce stade l’invagination gastruliènne est terminée, la ligne primitive commence à régresser et le nœud de Hensen a reculé pour former le canal de Liberkuhn.
A mesure que recule le nœud de Hensen, la gouttière chordale s’enfonce de plus en plus d’avant en arrière.
d) Stade plaque chordal
Vers le 21ème jour, la gouttière chordale s’étale sous forme d’une plaque allongée : c’est la plaque chordale, elle occupe la region médiane du toit du lecithocele secondaire tout en maitenant sa continuité avec l’endoblaste.
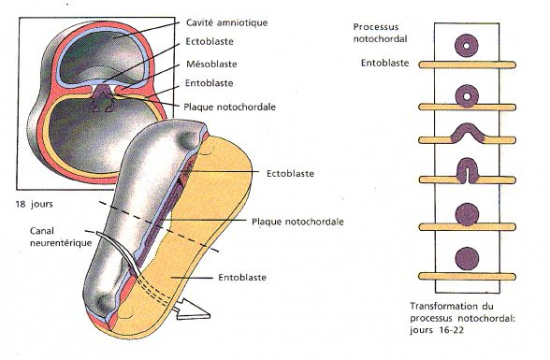
e) Stade tige plaine (chorde dorsale )
vers le 22ème jour, la plaque chordale se détache de l’endoblaste et s’enroule sur elle-même autour d’un axe longitudinal pour former la corde dorsale. A mesure que se détache la chorde, l’endoblaste rétablie sa continuité.
A la fin de la gastrulation la tige dorsale occupe l’axe de l’embryon compris entre les deux membranes pharyngienne et cloacale. Son toit est en contacte avec ectoblaste et son plancher est posé sur l’endoblaste.
Remarque : entre l’extrémité craniale du canal chordale et la membrane pharyngienne se situe la plaque préchordale qui dérive du mésoblaste. A partir du 18ème jour l’ectophylle et l’entophylle sont appelés respectivement ectoblaste et endoblaste du fait de la mise en place du chordo-mésoblaste.
3.La neurulation
Ce mouvement morphogénétique est imbriqué avec le mouvement de gastrulation , la limite n’est en réalité pas nette ( voir séquences temporelles.)
C’est le moment de la formation du système nerveux. Alors qu’au stade précédent, les contacts qui s’établissent entre le mésoderme dorsal et l’ectoderme induisent le neurectoderme, au stade neurula, l’ébauche du système nerveux se forme dans la région dorsale de l’embryon.
1.LA MISE EN PLACE DE LA CHORDE ET LE DEBUT DE LA NEURULATION (oeuf d’oiseau)
Le processus de gastrulation se poursuit et va directement induire la neurulation.
Dès le 16ème jour, à partir du noeud de Hensen, des cellules vont proliférer et former un tube creux, le processus notochordal (ou tube notochordal), qui se dirige vers la future zone céphalique de l’embryon.
Plus en avant de la zone de prolifération du tube notochordal, on observe une densification mésoblastique : la zone préchordale (au même titre qu’il existe une zone dense près de la région postérieure : l’éminence caudale).
Le tube notochordal va alors subir des remaniements qui le transforment, vers le 22ème jour, en un tube cellulaire plein : la chorde dorsale.
En effet, le tube notochordal vient d’abord s’accoler avec l’entoblaste, pour ensuite fusionner avec lui. Le processus notochordal est alors intégré dans l’entoblaste selon l’axe médian de l’embryon et forme la plaque notochordale. Ultérieurement, la plaque notochordale se creuse à nouveau en gouttière ; elle se désolidarise de l’entoblaste et forme la structure longitudinale définitive : la chorde dorsale, cylindre solide dans l’axe céphalique du futur embryon.
Simultanément à la croissance du processus notochordal et à la formation de la chorde, la ligne primitive régresse. In fine, elle ne représentera guère plus de 5 à 10 % de l’axe longitudinal du disque embryonnaire (contre plus de 50 % au 17/18ème jour).
L’ individualisation complète de la chorde est un processus qui déborde la 3ème semaine et ne s’achève que vers le 22-24ème jour. Entre-temps, l’intégration du processus notochordal à l’entoblaste avait ménagé localement un orifice au niveau du noeud de Hensen, orifice qui mettait transitoirement en communication la cavité amniotique avec la cavité vitelline : il s’agit du canal neurentérique.
La mise en place de la chorde vient induire l’ectoblaste sus-jacent dans le territoire présomptif qui génèrera le futur système nerveux (par le processus de neurulation). Dans ce territoire, l’ectoblaste s’épaissit et forme la plaque neurale qui s’agrandit antérieurement pour acquérir un aspect lobaire qui couvre la majorité de la zone craniale épiblastique, vers le 20ème jour.
La plaque neurale formera le cerveau et la moëlle épinière. C’est sa partie large et craniale qui produira le cerveau, alors que sa zone caudale (plus proche du noeud de Hensen) va assez rapidement se creuser en gouttière neurale qui sera à l’origine de la moëlle épinière.

2.LES PHASES MORPHOLOGIQUES DE LA NEURULATION
1° Epaississement de l’ectoblaste au dessus de l’ébauche cordale.
2° La plaque neurale se creuse en gouttière ; les cellules des bords de la plaque prolifèrent , et donc se surélèvent.
3° Rapprochement des bords : tube nerveux ou tube neural ou tube épithélial.
4° Le tube neural se détache de l’ectoblaste
5° Des cellules des bords de la gouttière se multiplient activement et s’en détachent : crêtes ganglionnaires ou bandelettes ganglionnaires.
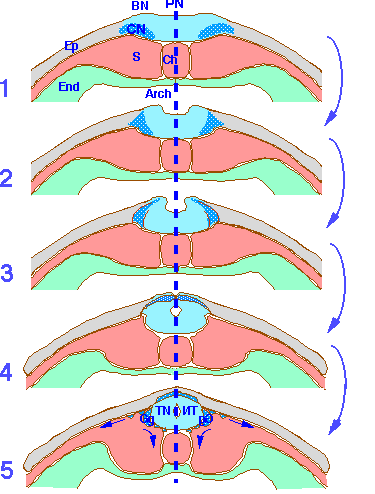
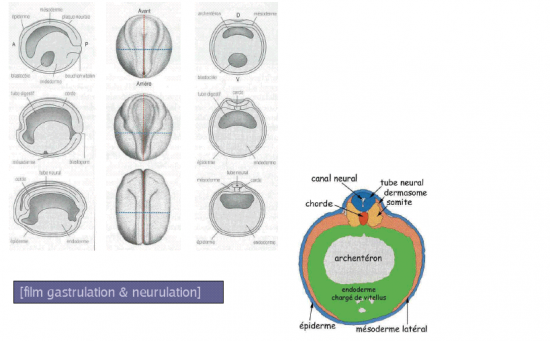
Schémas interprètatifs des mouvements du neurectoderme au cours de la neurulation sur des sections transversales d’embryons depuis le stade de la plaque neurale (PN)(1) jusqu’à la formation du tube neural (TN) (5). On retrouve l’épaississement du neurectoderme par rapport à l’épiderme limitant (1), le soulèvement des bourrelets neuraux (BN) (1,2), le rapprochement des bourrelets neuraux vers le plan médian (en tireté bleu) (3), l’affrontement des bourrelets neuraux (4) et la soudure des bourrelets neuraux dont l’aboutissement est la formation du tube neural qui se sépare de l’épiderme (Ep). On remarque que l’épiderme dorsal accompagne le neurectoderme dans un mouvement de recouvrement du tube neural. Le neurectoderme est composé de deux populations de cellules à vocation différente. En bleu clair, sont représentés les tissus à l’origine du tube neural sensu stricto. A la limite du neurectoderme et de l’épiderme, en bleu tramé, sont représentées les cellules de la crête neurale qui s’individualisent au moment de la soudure des bourrelets neuraux. Entre autres dérivés, elles sont à l’origine des ganglions rachidiens (Gg) qui se localisent à proximité du tube neural. Arch: Cavité de l’archentéron, Ch: chorde, End: endoderme, S: somites.
3.LE DEBUT DE LA METAMERISATION
Simultanément à l’apparition de la plaque neurale, le mésoblaste intra-embryonnaire se différencie.
D’abord dans la zone céphalique, à partir du 18/19ème jour, le mésoblaste para-axial se condense en blocs segmentaires de cellules qui se disposent en spirales et constituent les somitomères. Les 7 premièrs somitomères, à l’extrémité de la zone céphalique (proches de la membrane pharyngienne), vont dégénérer et tout au plus fourniront un contigent cellulaire pour la formation des muscles de la face, de la mâchoire et du pharynx.
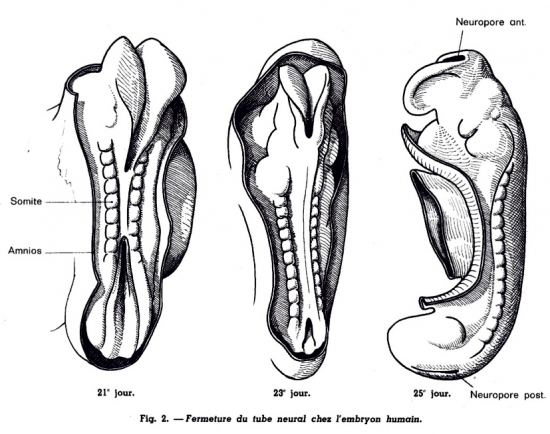
Les somitomères sous jacents vont poursuivre leur développement en blocs "métamérisés" dans le sens cranio caudal. La densification cellulaire s’accroit et aboutit à la formation de somites, structures cellulaires compactes de part et d’autre de la chorde dorsale.
A la fin de la 3ème semaine (21ème jour), une seule paire de somites s’est déjà constituée. Mais ultérieurement et au cours de la 4ème semaine, la somitisation (ou métamérisation) se poursuit dans le sens cranio caudal. Elle aboutira ultérieurement à la mise en place de 44 paires
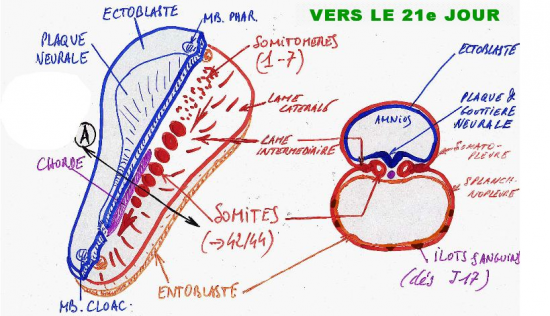
somites étagées de la zone cervicale jusqu’à la zone sacro-coccygienne.
La métamérisation est d’abord incomplète et les somites, même individualisés, restent reliés au mésoblaste par une zone compacte et plane, la lame intermédiaire (nous appelons somite primordial, un somite encore indifférencié et relié à la lame intermédiaire).
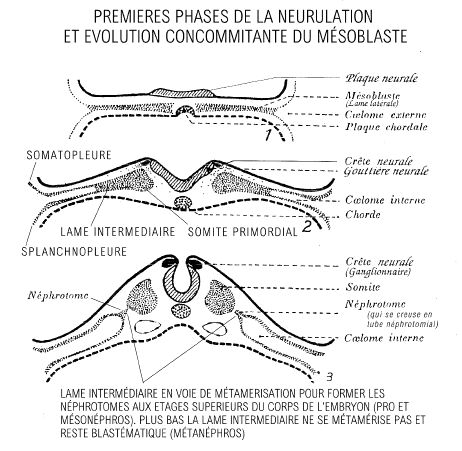
A la périphérie de la lame intermédiaire, le mésoblaste se clive en 2 feuillets distincts qui tapissent soit l’ectoblaste (ce feuillet porte alors le nom de somatopleure), soit l’entoblaste (ce feuillet porte alors le nom de splanchnopleure) du disque embryonnaire. La somatopleure et la splanchnopleure embryonnaires viennent fusionner avec les feuillets correspondants du mésoblaste extra-embryonnaire déja formé antérieurement et au cours de la deuxième semaine (voir chapitre précédent).
Date de dernière mise à jour : 02/04/2018
Commentaires
-
 tres bien explique, j'ai enfin pu comprendre.
tres bien explique, j'ai enfin pu comprendre. -
Super vos schémas m'ont bien éclairé là où j'étais un peu perdu :)
-
 Merçi bcp ^^
Merçi bcp ^^ -

 merci
merci
-
c' est une bonne production

Ajouter un commentaire


