

EMBRYO DESCRIPTIVE 2
Au centre de l’ovotide, les 2 pro-noyaux s’accolent étroitement. Autour, les fibres d’un fuseau achromatique s’organisent à partir du diaster, dont l’origine est probablement le spermaster.
L’origine du diaster n’est pas certaine dans l’espèce humaine. Les mécanismes de constitution de ce fuseau sont variables selon les espèces étudiées. Chez les non-Mammifères et chez la brebis, le spermaster se divise pour donner un diaster. Par contre, chez la souris, il existe des centrosomes ovocytaires qui donnent de multiples asters.
Les chromosomes se condensent progressivement.
L’enveloppe des pronucléus se résorbe et les chromosomes condensés se disposent sur un plan au sein d’un fuseau achromatique dépourvu de centrioles.
Ainsi on passe d’emblée à la métaphase de la première mitose de segmentation de l’oeuf vers 20 à 24h après la fusion gamétique.
LES CONSEQUENCES DE LA FECONDATION
* - Le retour à deux cellules diploïdes après une phase transitoire 4n ADN.
* - La création d’un nouvel individu génétiquement original par mélange des génomes parentaux. Il y a transmission des caractères héréditaires, mais aussi brassage des gènes au cours de la méiose. Ceci est le but de la reproduction sexuée. Le brassage des gènes concerne l’hérédité nucléaire. L’hérédité mitochondriale est uniquement maternelle.
* - La détermination de son sexe génétique par le gonosome apporté par le spermatozoïde.Les génomes paternel et maternel ne sont pas totalement équivalents. L’empreinte génomique désigne une expression différentielle de ces deux génomes. Ce phénomène est responsable, par exemple, de la différence phénotypique entre le mulet (hybride de cheval et d’ânesse) et le bardot (hybride d’âne et de jument) : le bardot ressemble plus au cheval que le mulet.
La période du développement fait suite à la fécondation. Les multiplications cellulaires se succèdent, tandis que l’expression du génome de l’oeuf entraîne une différenciation progressive en feuillets, en tissus, et en organes, suivant un plan propre à chaque espèce.
Chez les vertébrés terrestres, le développement de l’embryon s’accompagne de la formation de tissus annexes, nécessaires à sa protection et à sa nutrition : amnios, allantoïde, lécithocèle secondaire (ou vésicule vitelline).
Chez les Mammifères placentaires, l’embryon s’implante dans l’utérus maternel. Une annexe spécialisée supplémentaire, le placenta, assure sa nutrition à partir de l’organisme maternel.
Durant sa croissance, l’embryon, puis le foetus, doit s’adapter en permanence à son environnement. Il existe à tout moment un équilibre fragile entre ses besoins (nutrition, oxygénation, ...) et les possibilités d’apport de l’organisme maternel.
Le développement embryonnaire comprend 2 étapes :
L’embryogenèse
C’est la formation de l’embryon avec mise en place des premières ébauches des organes.
Elle comprend 3 phases :
- Une phase de pré-morphogenèse. Elle correspond à la segmentation de l’oeuf qui devient une morula,une blastula correspondant au blastocyste pour les Mammifères . L’aspect extérieur change peu (l’oeuf reste une masse arrondie).
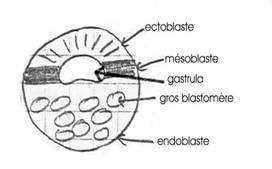
- Une phase de morphogenèse primordiale, durant laquelle se forment les feuillets embryonnaires pour aboutir à la gastrula. Chez les Mammifères, cette période se décompose en :
- Une phase de pré-gastrulation, durant laquelle s’individualisent 2 feuillets provisoires, l’épiblaste et l’hypoblaste (ou entoblaste primitif).
- Une phase de gastrulation proprement dite au cours de laquelle se forme les 3 feuillet définitifs (ectoblaste, mésoblaste et entoblaste).
- Une phase de morphogenèse secondaire, durant laquelle se placent les ébauches des futurs organes. Elle s’accompagne de la fermeture ventrale de l’embryon.
L’organogenèse
C’est la formation des organes.
Elle débute à la 4e semaine du développement et se poursuit pendant le 2e mois.
A partir du début du 3e mois, débute la période foetale, ou période d’histogenèse, aucours de laquelle se précise la structure histologique des organes qui, pour la plupart, deviennent fonctionnels.
DEFINITIONS
- Le terme d’oeuf désigne le zygote,
- Le terme de pré-embryon désigne l’organisme durant la segmentation. A ce stade, les cellules de l’embryon ne sont pas distinctes de celles qui donneront les annexes.
- Le terme d’embryon désigne l’ensemble des cellules à l’origine de l’individu, à l’exclusion des annexes.
- Le terme de foetus désigne l’embryon à partir du moment où il a pris les caractéristiques morphologiques de l’espèce, c’est à dire à la fin du 2e mois.
- Le terme de conceptus a un sens beaucoup plus large et désigne tous les stades.
Parmi les mécanismes du développement :
- La détermination : C’est l’engagement d’une cellule dans une lignée cellulaire définie avant la différenciation.
- La différenciation : C’est l’acquisition, généralement irréversible, de caractères cellulaires propres. Au cours du développement, les premières cellules sont pluripotentes. Puis, lors de la construction de l’individu, les cellules se différencient progressivement, constituant des lignées cellulaires dont la destinée sera de plus en plus restreinte à une fonction ou a un tissu précis.
2° La segmentation méroblastique ou partielle
Pour les oeufs télolécithes, centrolécithes, alécithes
Les oeufs renfermant beaucoup de vitellus ( télolécithes, centrolécithes) ou pas de vitellus ( alécithes ) subissent une segmentation méroblastique: seulement une portion du cytoplasme se segmente car l’axe de division ne pénètre pas ou ne pénètre qu’incomplètement le vitellus. La segmentation méroblastique peut être:
discoïde chez les oeufs télolécithes: vitellus abondant au pôle végétatif
superficielle chez les oeufs centrolécithes: vitellus abondant central

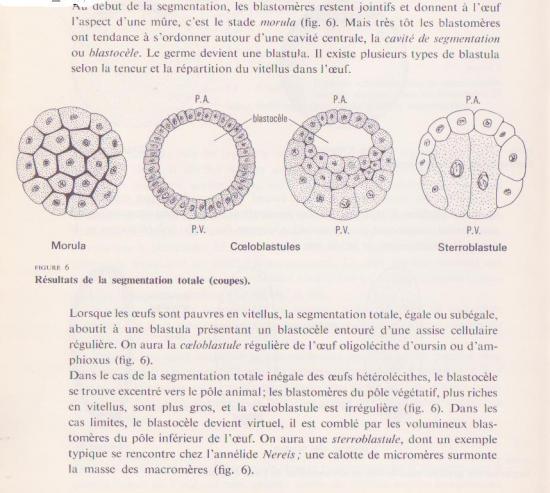
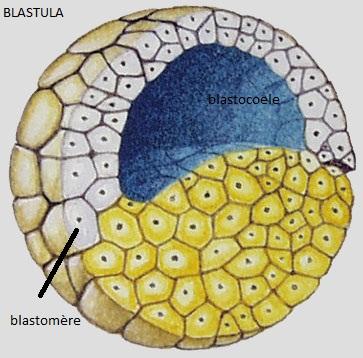
Le résultat de la segmentation : formation d’un germe pluricellulaire creusé d’une cavité : la blastula , les cellules constituant la blastula sont les blastomères
La cavité centrale : le blastocoele
exemple de Blastula
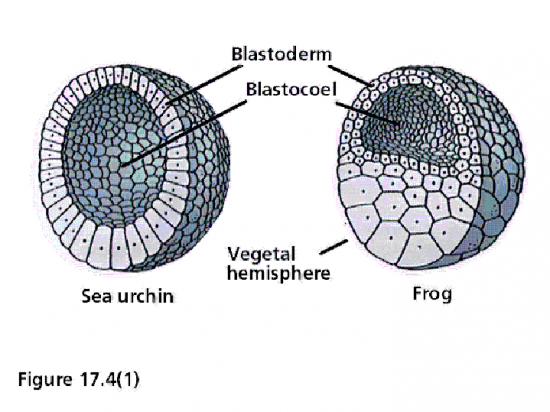 Blastula d’oursin et de Grenouille.
Blastula d’oursin et de Grenouille.1° Segmentation de l’œuf oligolécithe d’Echinoderme
Le Concombre de mer
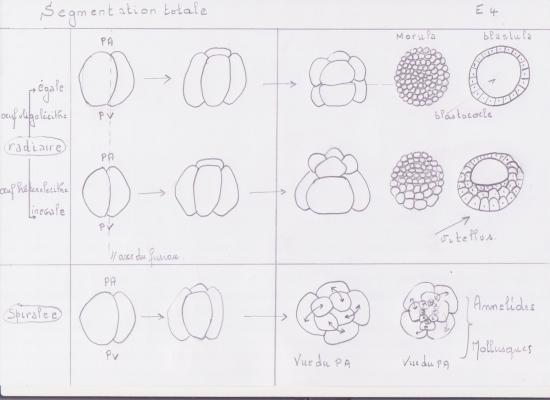
Forme la plus simple de segmentation. L’oeuf des échinodermes est oligolécithe, il renferme peu de réserves vitellines et celles-ci sont réparties uniformément dans le cytoplasme. Après l’union des deux pronoyaux, l’axe du premier fuseau mitotique se forme perpendiculaire à l’axe A-V de l’oeuf fécondé; donc le premier clivage passe directement au travers des pôles animal et végétatif. Comme le plan de clivage coupe ces deux pôles, on dit qu’il est méridional. Comme les deux blastomères obtenus sont de taille égale, c’est un clivage égal. Les fuseaux mitotiques de la seconde division sont également perpendiculaires à l’axe A-V, mais à angle droit par rapport au premier fuseau. La division coupe les pôles A-V, se produit simultanément dans les deux blastomères et résulte en la formation de 4 blastomères égaux. Elle est méridionale et égale. Le troisième clivage ne passe plus par les pôles A-V de l’embryon mais, pour chacun des 4 blastomères, est perpendiculaire à cet axe car les fuseaux mitotiques sont dans l’axe A-V. On dit que le clivage est équatorial. Il sépare l’embryon en 8 blastomères égaux.
La quatrième division est méridionale et produit 16 blastomères égaux.
Les divisions successives se font en alternance selon les plans équatorial et méridional, jusqu’au stade 256 cellules.
Le résultat de la segmentation est une petite boule, la blastula, dont le volume est sensiblement égal à celui de l’oeuf initial et dont les petits blastomères ont tous la même taille. Au cours des divisions, les contacts intercellulaires se sont relâchés de sorte que les blastomères ne s’agrègent pas en une masse compacte, mais décrivent une sphère d’une seule assise cellulaire autour d’une cavité remplie de fluide, le blastocoele. Les cellules qui se jouxtent établissent des jonctions entre elles. La blastula est entourée de l’enveloppe de fertilisation.
Les blastomères sécrètent des protéines dans le blastocoele; le fluide qu’il renferme devient sirupeux et a la propension de retenir beaucoup d’eau. Ainsi, au cours de la segmentation, de l’eau pénètre dans le blastocoele par osmose, à partir du milieu aqueux externe, et fait enfler la blastula. Cette pression exercée sur les blastomères pourrait aider à les orienter de sorte à ce qu’ils décrivent une sphère d’une seule assise cellulaire autour du blastocoele.
Les cellules possèdent des cils à leur surface externe. Leur battement impute à la blastula un mouvement de rotation à l’intérieur de l’enveloppe de fertilisation. Peu après la fin de la segmentation, les blastomères sécrètent une enzyme, l’enzyme d’éclosion, qui digère l’enveloppe de fertilisation. L’embryon devient une larve nageant librement.
À n’importe quel moment pendant la segmentation, un embryon coupé en deux selon n’importe quel méridien produira deux moitiés qui sont identiques. Ce type de symétrie est caractéristique d’une sphère ou d’un cylindre, il s’agit de la symétrie radiaire.
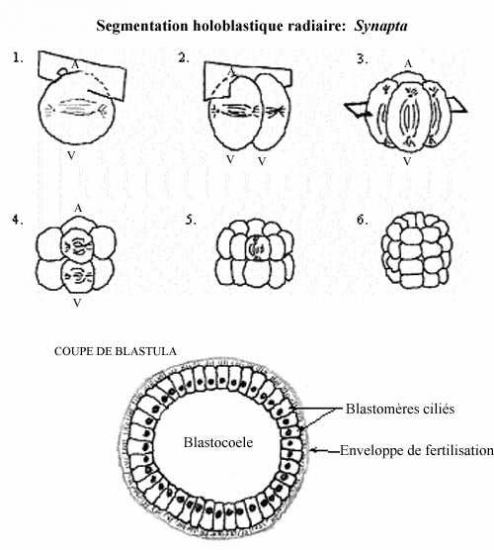
L’oursin
L’oursin possède lui aussi un oeuf oligolécithe.
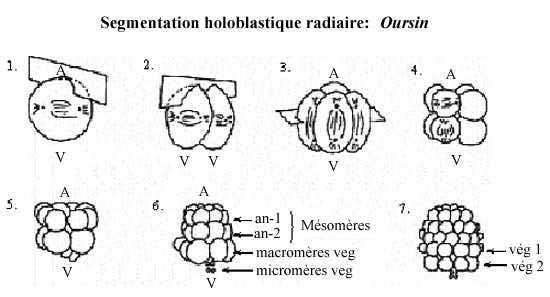
Ses 3 premiers clivages ressemblent à ceux du concombre de mer Synapta. Au 4e clivage, les 4 blastomères de l’hémisphère animal se divisent selon l’axe méridional et donnent une assise de 8 blastomères de taille égale appelés mésomères; les blastomères de l’hémisphère végétatif se divisent selon l’axe équatorial mais inégalement, en réalité sous l’équateur, et donnent 4 grosses cellules appelées macromères, sous les mésomères, et 4 petits micromères, sous les macromères.
Au clivage suivant, les 8 mésomères se divisent équatorialement (également), donnant deux assises animales, l’une au dessus de l’autre: an-1 et an-2. Les macromères se divisent méridionalement, donnant une assise de 8 macromères; les micromères, en se divisant, forment un petit amas sous les macromères.
Tous les plans de clivage de la 6e division sont équatoriaux; les macromères forment donc deux assises végétatives, l’une au dessus de l’autre: vég-1 et vég-2. Les 7e divisions sont méridionales. Il en résulte une blastula de 128 cellules, creusée d’un blastocoele rempli de fluide, et entourée de l’enveloppe de fertilisation. Une larve se forme comme chez le Concombre de mer.
Point de départ
Chez les Anoures, la fécondation est externe. Le mâle va chevaucher la femelle en répandant son sperme au moment où les œufs sont pondus.
La fécondation doit avoir lieu dès l’émission des ovocytes par la femelle. La gangue, hydrophile, gonfle au contact de l’eau et devient rapidement un obstacle à la pénétration des spermatozoïdes (fécondation).
Chez les Urodèles, la fécondation a lieu au niveau du cloaque de la femelle ; il n’y a pas d’organe copulateur. Le mâle émet un spermatophore (formation mucilagineuse) au niveau duquel sont agglutinés les spermatozoïdes. Ce spermatophore va adhérer au substrat (caillou).
La fécondation est dite interne et se produit au niveau du cloaque (canal mixte urogénital et dont les lèvres viennent enserrer le spermatophore).
La pénétration des spermatozoïdes a lieu généralement au niveau du pôle animal car c’est là où il y a le moins d’encombrement.
Il y a monospermie chez les anoures (un seul spermatozoïde rentre) mais polyspermie chez les urodèles (5 ou 6 spermatozoïdes rentrent mais un seul va féconder le noyau). La fécondation va entraîner une activation dans l’ovocyte : deux types d’activation (interne et externe).
C’est une phase de divisions cellulaires : on passe d’un œuf unicellulaire à une blastula (6 à 10000 cellules). Cette segmentation implique de nombreux facteurs qui vont orienter les fuseaux.
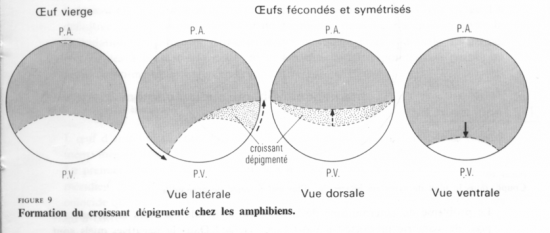
Le premier plan de division passe par l’axe pôle animal/pôle végétatif, dit méridien et donne 2 blastomères égaux. Ce plan peut couper, ou non, le croissant gris.
Le deuxième plan de clivage est parallèle au premier et donne 2 méridiens égaux.
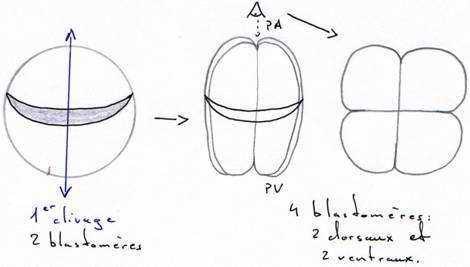
La segmentation est totale, et, pendant les deux premières divisions, elle est égale.
Au cours du troisième cycle cellulaire, la segmentation devient inégale (subéquatoriale) et donne 4 micromères au pôle animal et 4 macromères au pôle végétatif.
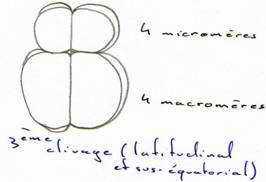
Les cycles cellulaires se poursuivent rapidement, sont synchrones jusqu’à la 10ème ou 12ème division puis, deviennent asynchrones. Ce cycle qui fait passer de la division synchrone à la division asynchrone est appelé transition blastuléenne.
Les cycles sont rapides jusqu’à la mi-blastula (pas de phase G1 ni G2).
Il y a une expression maternelle des gènes : toutes les protéines synthétisées le sont à partir d’ARN maternel stocké. Après la transition blastuléenne, les cycles s’allongent et apparaissent les phases G1 et G2 (G1, S, G2, M).
On a l’apparition d’un asynchronisme des divisions cellulaires. C’est une expression zygotique des gènes.
Au départ, on a un stade morula. Les cellules deviennent de plus en plus petites et vers la mi-segmentation, on aura le stade blastula.
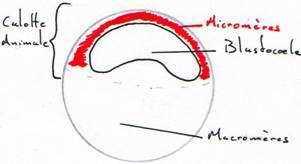
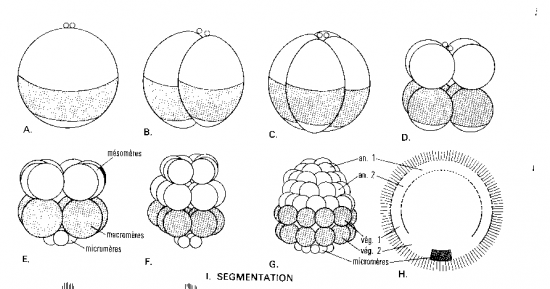
L’oeuf hétérolécithe d’amphibien possède plus de vitellus que celui des échinodermes et moins que celui d’oiseau, vitellus qui devient concentré au pôle végétatif après la fécondation (oeuf hétérolécithe), et le noyau migre au pôle animal.La 1ère division commence au pôle animal de l’oeuf et s’étend lentement vers le pôle végétatif; elle est méridionale et égale. Avant que cette division ne soit achevée, un second clivage méridional s’amorce au pôle animal, perpendiculaire au premier, lui aussi égal.
Le 3e clivage est équatorial; il ne s’effectue pas au centre de l’embryon mais plus près du pôle animal (donc, sus-équatorial), là où les plans de clivages des deux divisions précédentes ont achevé de traverser; il est donc inégal. Les plans de clivage méridionaux, du moins le second, n’ont pas encore achevé de traverser le cytoplasme de l’hémisphère végétatif quand a lieu ce 3e clivage sus-équatorial. Cette division sépare l’embryon en 4 petits blastomères animaux, les micromères, et 4 gros macromères végétatifs (qui éventuellement s’individualisent). Ce clivage holoblastique inégal établit deux régions embryonnaires majeures: une région de micromères qui se diviseront rapidement, dans l’hémisphère animal, et une région de macromères qui se diviseront lentement, dans l’hémisphère végétatif.
Les embryons amphibiens aux stades 16 à 64 cellules portent le nom de morula, à cause de leur aspect de mûre. Au stade 128 cellules l’embryon est considéré une blastula, creusée d’un blastocoele . Il occupe l’hémisphère animal; son plafond est constitué de plus d’une couche de micromères et son plancher de plusieurs assises de macromères.
Le blastocoele est une cavité qui permettra l’occurrence des migrations cellulaires pendant la gastrulation. Il empêche une interaction prématurée entre les cellules qu’il sépare. Par exemple, si les cellules du toit du blastocoele étaient apposées à celles du plancher, au lieu de se différencier en ectoderme comme elles le font normalement, elles se différencieraient en mésoderme. Les cellules du plancher du blastocoele, qui se différencieront en endoderme, induisent la différenciation des cellules qui leur sont voisines en mésoderme. Nous reviendrons sur ces notions. Donc, le blastocoele permet aux cellules de la blastula de conserver leur identité.
3 ° Segmentation de l’oeuf isolécithe d’Annélide ( holoblastique spiralée )
Se rencontre notamment chez les mollusques bivalves et les annélides, qui ont des oeufs isolécithes, pauvres en vitellus.
Au lieu de se produire parallèle ou perpendiculaire à l’axe A-V de l’oeuf, le clivage se produit à angle oblique.
En général, les deux premières divisions sont à peu près méridionales et égales.

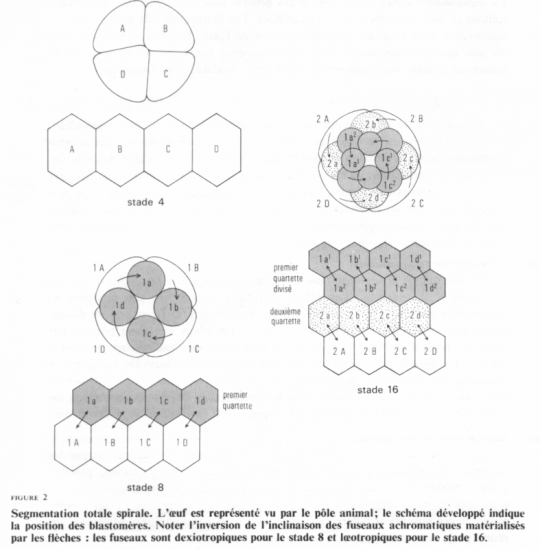
Cette blastula ne renferme pas de blastocoele et porte le nom de sterroblastula. Ses cellules sont en contact sur une plus grande surface que ne le sont les blastomères qui subissent un clivage radiaire et qui abritent un blastocoele. Ces embryons accomplissent un moins grand nombre de divisions avant d’entreprendre l’étape subséquente de développement.
voir schéma dans " types de segmentation "
4° Segmentation de l’oeuf télolécithe d’oiseau ( méroblastique discoïdale-oeuf télolécithe )


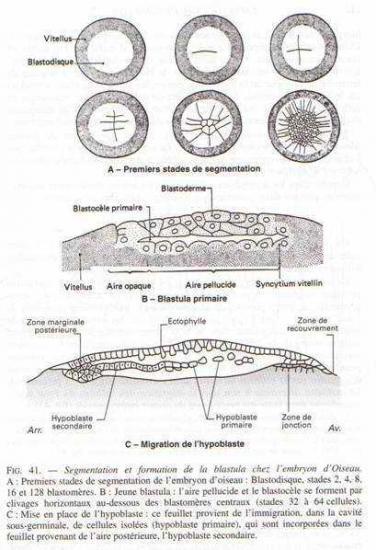
Un feuillet interne, l’entophylle ou hypoblaste, va doubler le feuillet externe, l’ectophylle ou épiblaste qui s’étend et s’amincit. Ces deux feuillets délimitent un blastocœle homologue du blastocœle des Amphibiens.
Le mode de formation de l’hypoblaste est encore discuté. Il provient d’une première migration en profondeur (polyinvagination), dans la cavité sous-germinale, de petits groupes de cellules ou de cellules isolées provenant de l’aire pellucide, c’est l’hypoblaste primaire; elle est suivie d’une seconde migration plus importante, dans le sens postéro-antérieur, d’un feuillet de cellules issues de la partie postérieure de l’aire pellucide, c’est l’hypoblaste secondaire qui rejoint et englobe les îlots de cellules de l’hypoblaste primaire pour former l’hypoblaste (fig.précédente ). La direction de cette dernière migration est déterminée par la rotation de l’œuf dans l’oviducte , la présence de l’hypoblaste détermine à son tour la migration des cellules du futur endomésoderme dans la moitié postérieure du disque embryonnaire.
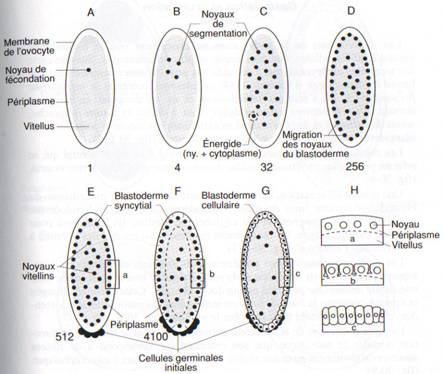
Rencontré chez les oeufs centrolécithes d’insectes, riches en vitellus central.
La segmentation offre ici aussi un exemple unique. Le noyau du zygote, en plein dans la masse vitelline, entreprend de nombreuses caryocinèses sans que ne se produisent de cytocinèses; ces noyaux s’appellent énergides. Ils migrent à la périphérie de l’oeuf, et les caryocinèses s’y poursuivent à un rythme prodigieux. L’embryon porte le nom de blastoderme syncitial: tous les noyaux du clivage sont contenus dans un même cytoplasme.
Des noyaux migrent au pôle postérieur de l’oeuf et s’enveloppent de membrane cytoplasmique; ce sont les cellules polaires de l’embryon. Elles donneront naissance aux cellules
germinales (reproductrices) de l’animal adulte. Ainsi, un des premiers événements de l’embryogenèse des insectes est la distinction des cellules germinales et somatiques.
6° Segmentation de l’oeuf alécithe de Mammifère.
- Fécondation interne, 1er tier de la trompe de Fallope
- cheminement dans la trompe : F + 30 h : 2 blastomères
F + 45 h : 4 blastomères
F + 60 h : 8 blastomères
F + 4 J : 16 blastomères
- dans l’utérus : F + 5 j : formation du blastocyste libre. Equivalent de la blastula .
Il se compose: - du bouton embryonnaire, massif cellulaire qui donnera l’embryon
- du trophoblaste qui participera à l’élaboration du placenta
F + 6 ou 7 J : implantation du blastocyste dans l’endomètre utérin.

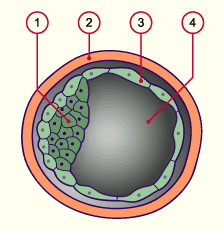
1 Embryoblaste
2 Zone pellucide
3 Trophoblaste
4 Blastocoele
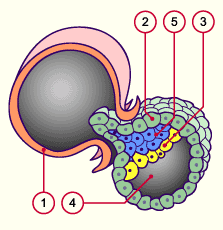
1 Zone pellucide
2 Trophoblaste
(masse cellulaire externe)
3 Hypoblaste (partie de la masse
cellulaire interne)
4 Blastocoele
5 Epiblaste (partie de la masse cellulaire interne)
Suite embryologie descriptive 3
Date de dernière mise à jour : 02/04/2018
Ajouter un commentaire