

Classification générale cycles 4
2.6 CLASSE DES BASIDIOMYCETES
2.6.1 Caractères généraux
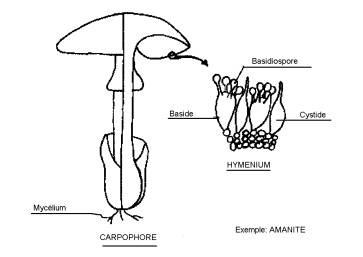
Ce sont généralement les "champignons" du langage courant : hyphes cloisonnés de structure articulaire ; fructifications ou carpophores contenant des basides. ( chez les Ascomycètes, les carpophores contenaient des asques )
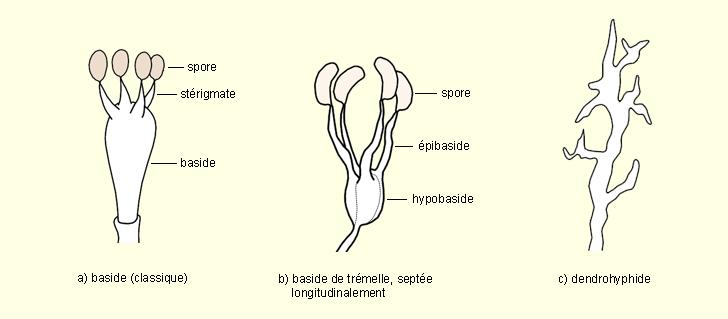
La baside et les basidiospores se présentent généralement comme suit : Baside = élément binucléé ( deux noyaux n ) dont les élément fusionnent à un moment donné ( noyau 2n = zygote ) à partir du dicaryon de la baside. Viennent ensuite les cinèses réductionnelles, donnant 4 noyaux n . Chaque noyau s'isole extérieurement à la baside, au sommet d'un pédicelle et devient une basidiospore qui est donc une exospore.
Comme chez les Ascomycètes, le mycélium est constitué au départ par deux filaments n ( il y a généralement hétérothallie ) . Ces filaments vont fusionner et deviennent des hyphes à dicaryons . La phase à dicaryons est beaucoup plus longue chez les Basidios que chez les Ascos : elle constitue l'essentiel du mycelium et du carpophore ( pied, chapeau avec son hymenium, anneau, volve, voile ).
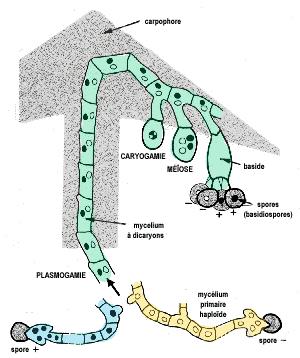
Chez les basidiomycètes, comme les coprins par exemple, la génération gamétophytique est représentée par des filaments mycéliens haploïdes. Ces filaments (= le mycélium primaire) réalisent la reproduction sexuée en s'unissant deux à deux, c'est une somatogamie ou périttogamie.
Dans un premier temps, seuls les cytoplasmes des cellules fusionnent, on dit qu'il y a plasmogamie. Ceci donne naissance à un mycélium dicaryotique (=mycélium secondaire), c'est à dire constitué de cellules à deux noyaux haploïdes. Cette phase dicaryotique spécifique des champignons constitue la génération sporophytique. Le mycélium dicaryotique s'organise en pseudotissus et forme le carpophore.
Au niveau des sporocystes, a lieu la fusion de deux noyaux haploïdes, ou caryogamie, bientôt suivie d'une méiose qui génèrera quatre spores méiotiques exogènes. Les sporocyte sont des basides et les spores méiotiques des basidiospores. Ces spores seront à l'origine du mycélium gamétophytique haploïde et le cycle est bouclé.
Notons encore que si le reproduction sexuée ( par le carpophore ) permet d'établir le cycle complet des champignons , il existe également une multiplication végétative par bourgeonnement ; cette dernière serait la plus fréquente.
C’est l’ensemble dans lequel nous allons trouver la grande majorité des champignons qui intéressent les mycophages et les mycologues amateurs et qui sont les mieux connus. Cependant pour ces derniers qui ont déjà acquis de bonnes connaissances de leur systématique il va falloir réviser quelques certitudes !
En premier lieu parce que il y a de multiples convergences de formes, de sorte que les termes classiques, Hyménomycètes, Aphyllophorales, Gastéromycètes tout comme « Clavaire », « Hydne », « Polypore », ..., n’ont plus qu’une valeur physionomique. De même que les distinctions entre Phragmobasidiés, Hétérobasidiés, Homobasidiés sont moins strictes que ce que l’on pensait . Comme si, dès le début de leur évolution, les Basidiomycètes ancestraux possédaient en puissance toutes ces possibilités morphologiques qui ont pu se révéler à plusieurs reprises et de façon indépendante au cours de leur différenciation.
Et tout d’abord où se situe l’origine des Basidiomycètes ? La question ne paraît pas résolue. La plupart des analyses phylogéniques montrent un enracinement commun avec les Ascomycètes quelque part dans les Zygomycètes. La seule chose qui paraisse certaine est leur monophylie, c’est à dire une origine commune aux deux embranchements actuels. Ce qui sépare en premier lieu les deux groupes est le type de cellule qui produit les spores reproductives, ici c’est donc la baside qui produit des basidiospores externes, 4 presque toujours. A la différence de l’asque qui ne montre que des variations de forme réduite, la baside présente des types morphologiques très variables.
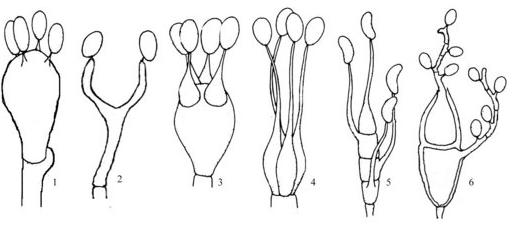 Différents types de basides. 1 : Holobaside d’Agaricale. 2 : Holobaside réduite de Calocera. 3 : Hétérobaside de Tulasnella avec épibasides. 4 : Hétérobaside de Tremella incomplétement divisée. 5 : Phragmobaside d’Auricularia. 6 : Phragmobasides de Puccinia issues d’une téliospore (probaside)
Différents types de basides. 1 : Holobaside d’Agaricale. 2 : Holobaside réduite de Calocera. 3 : Hétérobaside de Tulasnella avec épibasides. 4 : Hétérobaside de Tremella incomplétement divisée. 5 : Phragmobaside d’Auricularia. 6 : Phragmobasides de Puccinia issues d’une téliospore (probaside)
2.6.2 CLASSIFICATION GENERALE DES BASIDIOMYCETES
2.6.2.1 Classification moderne
Tout comme pour les ascomycètes, une classification moderne, basée sur la phylogénie s'ébauche, en voiçi la structure :
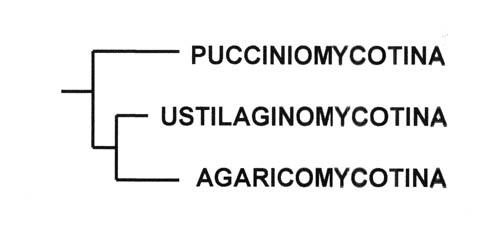
Voir le site de l'association mycologique de Toulouse : http://www.associationmycologiquetoulouse.ups-tlse.fr/spip.php?article33
Pucciniomycotina
(Uredinomycotina)
Plus de 8000 espèces décrites, dont la majeure partie appartiennent à l’ordre des Pucciniales (les Rouilles des végétaux). En dehors des phytoparasites, il existe aussi des mycoparasites, des parasites d’insectes et des saprotrophes. Certaines espèces sont levuriformes, la présence de basidiocarpes est exceptionnelle.
PUCCINIOMYCETES (Urédinomycètes)
Ce sont tous des parasites, ils possèdent une phragmobaside le plus souvent issue d’une téliospore.
Pucciniales (Urédinales).
Les Rouilles des végétaux ont un cycle complexe dans lequel, en plus de la forme parfaite (téliospores), on peut observer des organes sexués (spermogonies) et des anamorphes de multiplication, successivement : écies (écidies) donnant les éciospores puis urédo produisant les urédospores. La baside naît d’une téliospore qui joue souvent un rôle de conservation (voir figure basides n°6). Beaucoup sont hétéroxènes, c’est à dire qu’elles accomplissent leur cycle sur 2 plantes appartenant à des familles différentes. L’exemple classique est celui de la rouille noire du blé, Puccinia graminis dont la phase écidienne est hébergée par des Berberis et la phase urédotélienne par des Graminées.
Ce cycle peut être plus ou moins réduit par disparition d’une ou plusieurs phases jusqu’à ne comporter que des téliospores comme chez Puccinia malvacearum. Les rouilles ont une grande importance économique en raison des dégâts qu’elles provoquent sur les cultures, céréales (Puccinia), légumineuses (Uromyces), florales (Phragmidium sur rosiers), caféier (Hemileia)...voir cycle dans classification traditionnelle
Trois autres ordres sont à citer pour leur curieuse biologie.
Helicobasidiales Ont d’abord une phase haploïde parasites dans les écies de Pucciniales (Tuberculina) et après fécondation. sur les racines de plantes pendant leur dicaryophase.
Septobasidiales Elles vivent en symbiose avec des cochenilles. Elles forment des croûtes à la surface des écorces, abritant ainsi des colonies de cochenilles dont elles parasitent un certain nombre d’individus.
Platygloelales A noter encore avec Eocronartium et Jola parasites des mousses aux minuscules fructifications clavarioïdes.
CYSTOBASIDIOMYCETES, AGARICOSTILBOMYCETES
Ces classes réunissent essentiellement des champignons totalement ou partiellement levuriformes.
MICROBOTRYOMYCETES
Principalement représentés par les Microbotryales, phytoparasites dont beaucoup d’espèces se localisent dans les anthères des fleurs. Etaient classées avec les Ustilaginales (les Charbons) en raison de leurs téliospores sphériques, plus ou moins ornementées ressemblant à celles de ces champignons. Leurs basidiospores germent en donnant une forme levure. Exemple Microbotryum violaceum = Ustilago violaca dans les anthères de certaines Caryophyllacées.
MIXIOMYCETES
Une seule espèce, Mixia osmundae, qui mérite d’être citée car elle a d’abord été décrite comme un Ascomycète.
Ustilaginomycotina
Réunissent près de 1500 espèces de parasites des végétaux. Beaucoup possèdent des téliospores et une phase haploïde levuriforme.
USTILAGINOMYCETES
Ce sont les « Charbons » dont l’importance économique est loin d’être négligeable, charbon du blé (Ustilago tritici), du maïs (Ustilago maydis ) par exemple. Leur sporulation constitue des amas (sores) de téliospores de couleur foncée. Ces spores sont libérées par éclatement des tissus de l’hôte. Leur germination donne une phragmobaside. Deux ordres :
Ustilaginales surtout parasites sur Graminées et Cypéracées.
Urocystales sur diverses Mono et Dicotylédones (Urocystis ficariae sur Ranunculus ficaria).
EXOBASIDIOMYCETES
Se différencie des précédents par la prèsence d’une holobaside. On y trouve des champignons produisant ou non des téliospores.
Tilletiales Avec des téliospores de couleur sombre qui, pour la plupart, se développent dans les ovaires de Graminées (Tilletia caries, la carie du blé).
Entylomatales, les téliospores sont peu ou pas colorées et dispersées dans les parenchymes foliaire de l’hôte. Exemple le très commun Entyloma ficariae.
Doassansiales Elles se distinguent principalement des précédentes par l’agglomération des téliospores en masse. Elles parasitent surtout des plantes aquatiques.
Exobasidiales Il n’y a pas de téliospores et les basides sont produites le plus souvent à la surface des organes parasités, feuilles principalement, qui sont plus ou moins profondément déformés, l’exemple type est Exobasidium rhododendri qui produit de grosses galles vivement colorées sur le feuillage des Rhododendrons.
Microstromatales et Georgesfischeriales ne regroupent qu’un petit nombre d’espèces.
ENTORRHIZOMYCETES
Groupe très réduit (un seul genre, Entorrhiza) qui se distingue par des téliospores se différenciant à l’intérieur des cellules vivantes dans les tissus des racines de l’hôte . Ces téliospores germent sans donner de baside extérieure mais en se cloisonnant et seuls 4 stérigmates émergent. Leur rattachement traditionnel aux Ustilaginomycotina, basé sur leur parasitisme, est douteux, certaines analyses les font apparaître à la base de l’ensemble des Basidiomycètes.
Agaricomycotina
Ils comprennent le tiers des espèces décrites de champignons et l’énorme majorité de celles qui intéressent le plus les mycologues amateurs. A part quelques unités, c’est parmi eux que l’on trouve les espèces comestibles, et aussi des plus toxiques. De plus leur importance écologique et économique est considérable. En tant que mycorhiziens ils contribuent à la croissance et au maintien d’une grande partie des forêts mondiales et par là à la production de biomasse, bois en particulier. Bois que bon nombre d’autres se chargent ensuite de détruire. Quelques font l’objet d’une culture et d’un commerce intensifs. Leur classification traditionnelle basée sur la structure de la baside (hétéro ou holobaside), le type de basidiocarpe, d’hyménium... a été profondément bouleversée. Les recherches phylogénétiques basées aujourd’hui non plus sur un seul mais sur plusieurs fragment de génome donnent des résultats relativement solides. Il semble donc que l’on puisse accepter trois ensembles (classes) d’importances inégales, Agaricomycètes, Dacrymycètes et Trémellomycètes.
TREMELLOMYCETES
C’est la classe qui paraît la plus proche de la souche commune des Basidiomycota. Les basides sont tétracellulaires avec des cloisons plus ou moins complètes disposées longitudinalement ou obliquement (voir figure basides n°4). Les basidiocarpes (quand ils sont présents) sont gélatineux. La phase haploïde est levuriforme.
Trémellales sont typiquement représentées par l’espèce commune Tremella mesenterica, saprotrophe sur bois mort. Beaucoup d’espèces bien moins spectaculaires sont des mycoparasites d’autres Basidiomycètes.
Filobasidiales ne produisent pas de basidiocarpe. Ces champignons sont surtout connus par leurs formes levures : Cryptococcus. L’une d’elle, C. neoformans, est un pathogène humain, très dangereux pour les immunodéficients.
DACRYMYCETES
Petit ensemble de d’espèces à basidiocarpes gélatineux de faible taille. La baside est unicellulaire prolongée de deux longs srérigmates qui lui donnent un aspect fourchu (voir figure basides n°2), holobaside, hétérobaside ? C’est comme l’on voudra ! Ce sont des décomposeurs de cellulose (pourritures brunes).
Un seul ordre, Dacrymycétales, exemples : Dacrymyces, Calocera.
AGARICOMYCETES
Parmi tout une série d’ordres dont les relations ne sont pas encore bien clairement définies se détachent deux ensembles auxquels on donne le rang de sous-classes, les Agaricomycétidées et les Phallomycétidées. Restent un certain nombre d’ordres pour lesquels il est encore difficile d’assigner une place exacte dans une phylogénie générale. Un ensemble se trouve près de la racine des Agaricomycètes. Ce sont d’une part des hétérobasidiés, Sébacinales et Auriculariales et d’autre part des holobasidiées, Cantharellales.
Sébacinales aux basidiomes réduits ou nuls avec basides renflées, subdivisées longitudinalement et surmontées de 4 épibasides allongées.. Elles jouent un rôle, longtemps méconnu mais important, dans la mycorhization de nombreuses plantes comme les Orchidées ou les Ericacées.
Auriculariales, elles possèdent des basidiocarpes gélatineux, leurs basides sont typiquement cloisonnées transversalement Les genres Auricularia et Exidia sont bien connus.
Cantharellales, homobasidiées, comprennent les Cantharellus, Craterellus et aussi Hydnum, Clavulina. Le sort des Tulasnellales, aux basides de forme particulière (figure basides n°3), n’est pas résolu, pour les uns cela reste un ordre distinct alors que certaines analyses les incluent dans les Cantharellales.
Le deuxième ensemble est constitué de champignons homobasidiés, très souvent lignicoles aux basidiomes de consistance coriace, parfois réduits « croutes ». On peut citer :
Corticiales
Polyporales : Polyporus, Fomitopsis, Laetiporus
Hyménochaetales : Coltricia, Inonotus, Phellinus
Téléphorales : Telephora
PHALLOMYCETIDEES
Elles ne correspondent pas exactement aux ex Gastéromycètes car ce regroupement concerne aussi des champignons qui n’y étaient pas inclus et en exclu au contraire d’autres.
Phallales. Les basidiospores baignent dans une gléba mucilagineuse à odeur puissante (dispersion entomophile) (Phallus, Mutinus, Clathrus...)
Gomphales. Réunissent avec les Gomphus, des types aux morphologies très diverses : coraloïde (Ramaria), en massue (Clavariadelphus), hypogée (Gautiera)...
De plus deux nouveaux ordres sont crées :
Géastrales pour les familles des Géastracées, Sclerogastracées et Sphaerobolaceae
Hystérangiales, essentiellement hypogées.
AGARICOMYCETIDEES
Nous voici au cœur de l’essentiel de la matière des atlas de terrain, de ce qui intéresse le plus l’amateur de champignons.
Russulales. Suivant les auteurs cet ordre est souvent considéré comme un groupe frère des autres Agaricomycetidées. La famille des Russulacées est évidemment bien connue : Russula, Lactarius constituent une part importante de la flore de nos sous bois. Oui mais, certaines espèces tropicales sont annelées ou ressemblent à des pleurotes tandis que des formes souterraines (Gymnomyces, Macowanites, Zelleromyces) viennent se loger au sein même de ces deux genres qui sont eux mêmes paraphylétiques. En effet les Russules du sous-genre Compacta sont plus proches des Lactaires blancs (L. piperatus ...) que des autres Russules. Et puis la biologie moléculaire a révélé des parentés à priori peu évidentes avec d’autres familles : Peniophoracées, Auriscalpiacées, Hericiacées, Stéréacées.... Ainsi sont inclus dans les Russulales des genres comme Albatrellus, Amylostereum, Artomyces, Auriscalpium, Hericium, Heterobasidion, Peniophora, Stereum. A noter que ces regroupements ne correspondent pas uniquement à des données biomoléculaires, on trouve de façon presque générale chez ces champignons des spores amyloïdes et des lacticifères.
Agaricales. C’est l’essentiel des champignons lamellés mais pas seulement.car il y a aussi des formes gastroïdes ou clavarioïdes.. On peut y distinguer 5 clades (rameaux) principaux : Plutéoïdes : Plutéacées, Amanitacées, Pleurotacées. Hygrophoroïdes : Hygrophoracées, Ptérulacées, Typhulacées. Marasmioïde : Marasmiacées, Omphalotacées, Schizophyllacées... Agaricoïdes : Agaricacées (inclus Lycoperdon), Strophariacées, Inocybacées Cortinariacées... et d’autres formes gastroîdes : Nidulariacées, Hymenogastracées. Tricholomatoïdes : Tricholomatacées, Lyophyllacées, Entolomatacées, Mycenacées...
Bolétales. On sait déjà qu’ils ne s’agit pas uniquement de formes charnues à hymenium poré. Ici aussi la biologie moléculaire apporte des précisions mais qui méritent encore d’être approfondies. On distingue un certain nombre de lignées bien individualisées (sous-ordres ou familles) : Boletinées, autour des genres Boletus, Xerocomus, Porphyrellus, Chalciporus, Phylloporus... Suillinées : Suillus, Gomphidius Rhizopogon... Paxillinées : Paxillus, Melanogaster... Sclerodermatinées : Gyroporus, Scleroderma, Calostoma, Pisolithus. Hygrophoropsidacées : Hygrophoropsis. Serpulaceae : Serpula Coniophoracées : Coniophora.
2.6.2.2 Classification traditionnelle
Comme argumenté dans le chapitre relatif aux ascomycètes, cette classification se révèle toujours très utile, surtout à des fins pratiques.
I. Absence de carpophore
Basides cloisonnées. Champignons parasites
----------------> Hémibasidiés (ou Hémibasidiomycètes)
II. Carpophores généralement présents
a) Carpophores gélatineux et basides cloisonnées, Champignons saprophytes
----------------> Protobasidés
b) Carpophores non gélatineux et basides unicellulaires
----------------> Autobasidiés
2.6.2.2.1 LA SOUS CLASSE DES HEMIBASIDIES
Ces champignons généralement parasites sont considérés comme de redoutales ennemis de nombreuses plantes cultivées
Du point de vue agronomique, cette sous-classe comporte deux Ordres importants :
- Ordre des Ustilaginales
- Ordre des Uredinales
A) O. Ustilaginales
Parasites des céréales, détruisant entièrement les organes floraux
Les ustilaginomycètes regroupent environ 1300 espèces qui sont des parasites de plantes à fleurs avec un seul hôte et sont donc comme les rouilles (pucciniomycètes) des pathogènes d'importance économique. D'autant plus qu'ils attaquent les fleurs et empêchent donc la formation des graines. A la fin de l'infection, ils forment souvent des amas de spores noires d'où le nom de charbon qui leur a été donné. Ils attaquent leur hôte de manière particulière en secrétant via des vésicules des dépôts à l'intérieur des cellules. Ils alternent souvent entre une phase mycélienne infectieuse et une phase levure saprophyte. Leurs pores septaux sont plus simples que chez les agaricomycotina et ne sont pas des dolipores caractéristiques. Deux grands groupes sont reconnus sur la base des données moléculaires.
- Genre Ustillago
exemple U.Tritici
C'est le "charbon" du Froment. Des spores noires germent avec les grains de pollen sur stigmates des fleurs : le mycélium envahit les ovules, puis ultérieurement les graines qui en sont issues sans leur causer de dommages.
Ce mycélium dormant se réveille avec la germination du grain au printemps suivant et envahit toute la plante issue du grain, détruisant les épis qui sont alors remplacés par des amas de spores.
L'attaque se produit donc en seconde généralement.
Les agriculteurs produisent de moins en moins leur semences, la maladie se raréfie donc. Traditionnellement on luttait contre cette maladie en traitant les semences à l'eau chaude sans en altérer les propriétés germinatives.

ou encore U. maydis
L'espèce la plus connue de ce groupe actuellement est Ustilago maydis qui attaque le maïs provoquant des galles (c'est à dire des tumeurs où le tissu de l'hôte est hypertrophié et mélangé avec le mycélium) qui remplace les grains dans l'épi. Le génome de ce champignon est maintenant complètement séquencé. Son analyse devrait permettre de mieux comprendre sa biologie. Le cycle est le suivant :
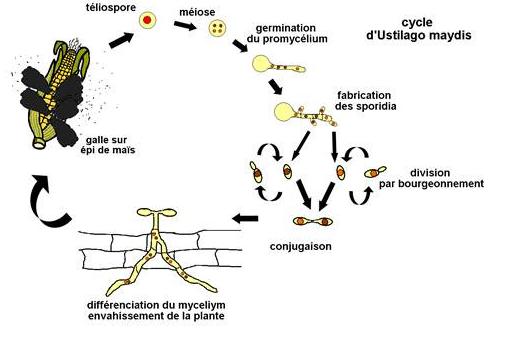
Le cycle commence par la germination de la spore diploïde (téliospore) au printemps qui s'accompagne de la méiose. Ceci donne naissance à un promycélium de 4 cellules haploïdes. Chacune de ces cellules produit au moins un bourgeon (le sporidium) qui est l'équivalent de la basidiospore. Ce processus peut se répéter plusieurs fois. Si des cellules compatibles se rencontrent, il y a fusion cellulaire et formation du dicaryon. Ceci entraîne la formation du mycélium et l'envahissement de la plante (qui ne se produit donc que s'il y a fusion sexuelle). Lorsqu'il atteint la surface de la plante, le mycélium peut aussi produire des spores dicaryotiques qui assurent une propagation asexuée. La fusion nucléaire intervient dans les téliospores en formation pendant l'automne dans des galles qui occupent la place des grains dans l'épi, terminant le cycle et assurant la production d'une forme de résistance pendant l'hiver;
- Genre Tilletia :
exemple T. tritici
C'est la "carie du froment".

Une basidiospore adhérant au grain infecte la toute jeune plante. A la floraison, l'ovaire est dévoré par le champignon et est remplacé par un amas de spores binucléées à membrane cellulosique très résistante et appelées chlamydospores. Ces spores tombent au sol. A l'automne, fusion des deux noyaux et formation du zygote. Au printemps suivant interviennent des cinèses réductionnelles. Les quatre petites cellules n obtenues forment de petits diverticules dans lesquelles s'engage chaque noyau = quatre basidiospores.
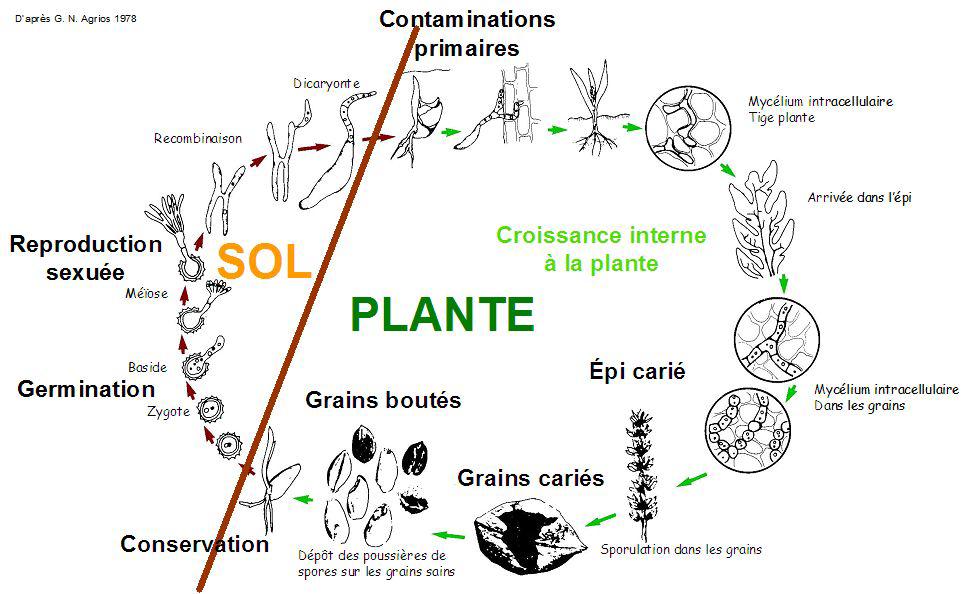
B) O. Uredinales
Parasites responsables des maladies connues sous le nom de " rouilles " : voir ci-après. Attaquent des milliers d'espèces de Phanérogames et surtout des graminées sociales ( exemples : céréales ) et surtout des graminées sociales ( exemple : céréales ).
En conditions d'années humides, les dégâts peuvent être énormes, par action sur les feuilles et développement de plusieurs types de spores dont un type de teinte jaune orange qui leur vaut leur nom.
Le cycle biologique est complexe, passant par un hôte intermédiaire.
Exemples :
- Puccinia graminis sur froment et épine vinette ( hôte secondaire )
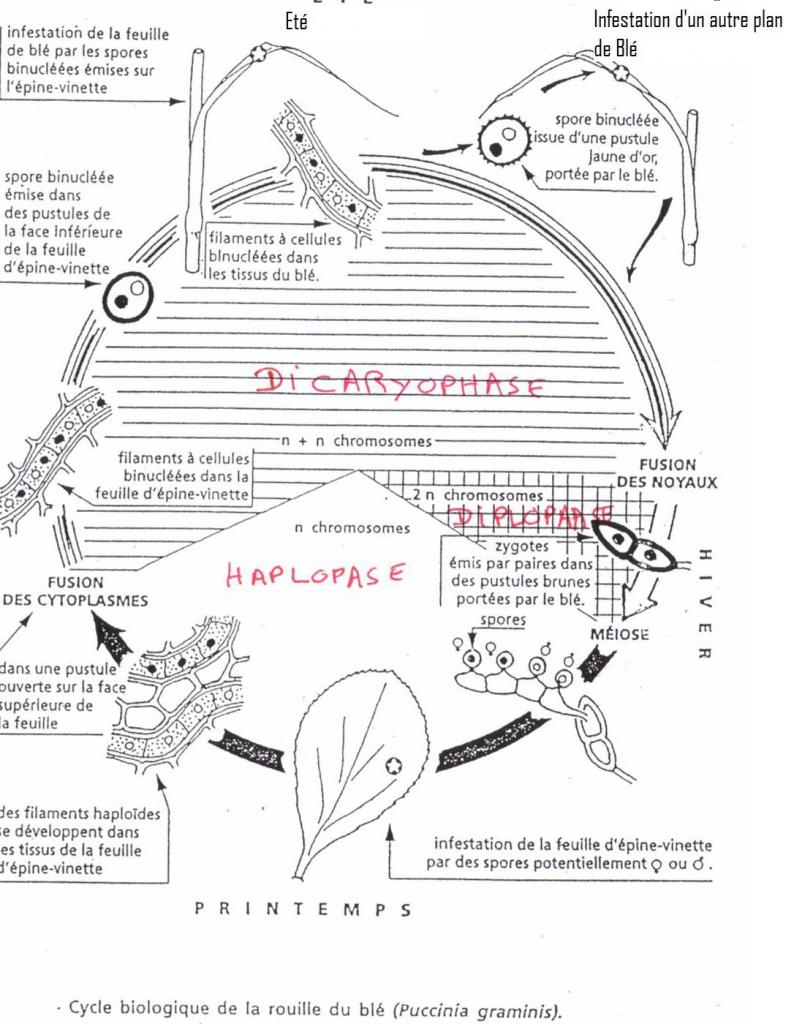
Pustules sur feuilles d'épine vinette
- Filament n *Picnides ---------> picnospores , face supérieure--------> cycle végétatif sur épine vinette------>picnides*
- Filament n*Ascidies ----------> ecidiospores , face inférieure( 2 noyaux)---------> infestation du plant de froment par les feuilles--------> **
Pustules jaune or sur feuilles de froment
**------> dicaryons dans tissus froment---> pustules jaune or( conidies- urédospores) - infestation d'autres plants de froment pas autre type de spore binucléée (conidies --urédospores)----------> dicaryons------>Téleutospores ( noires de survie )--------> caryogamie ------->zygote ----> méiose----->basides-->basidiospores-------------> retour sur feuilles épine vinette -------- filament n
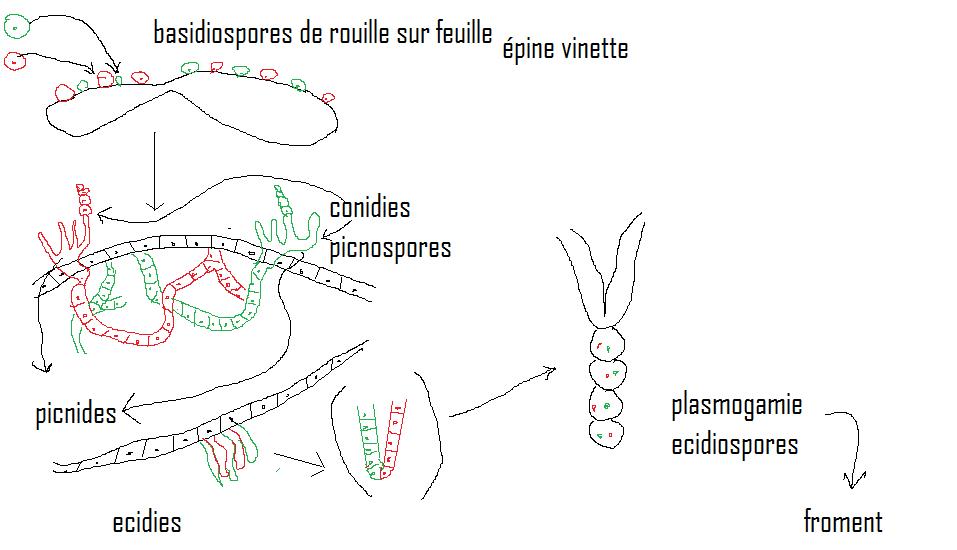
- Melampsora larici-populina sur peuplier et mélèze
- Melampsora allii-populina sur peuplier et Allium
- Autres Melampsoras avec d'autres hôtes
- Gymnosporangium sabine = rouille grillagée du poirier ( Téleutospores sur Genévrier )
-Autres rouilles : du Caféier, de la Betterave ( autoïque ), des Graminées de prairies; des Légumineuses; rouille " jaune " du froment, de l'orge, du seigle; rouille "couronnée" de l'avoine; des Malvacées, du rosier, etc...
2.6.2.2.2 SOUS-CLASSE DES PROTOBASIDIES
Champignons xylophages, exemples : Tremelles ( oreilles de judas ), Hirnéoles

Trémelles sur bois
Caractéristique de l'"oreille de Judas"
Fructification: sessile ou à pied rudimentaire, 2-8 cm de large et jusqu'à 4 cm de projection, en forme de bonnet, de coquille ou d'oreille, fixée latéralement au substrat ou par le sommet; à chair mince, gélatineuse-caoutchouteuse au frais, devenant cornée et dure en séchant, pouvant reprendre sa consistance initiale en présence d'eau, à odeur faible et à saveur agréable.
Face supérieure: stérile, habituellement veinée et côtelée, veloutée-tomenteuse, brun pâle, brune ou brun rougeâtre, noircissant en séchant.
Face inférieure: fertile, lisse à légèrement plissée, brun jaunâtre, brun rougeâtre, brun grisâtre ou brun pourpré.
Spores: allantoïdes, lisses, hyalines, guttulées, 12-16 x 4-7 µm. Sporée blanche.
Habitat: Habituellement en groupes sur les troncs morts de feuillus et de conifères, en été et en automne. Les individus qui ont poussé tard à l'automne peuvent se réhydrater après la fonte des neiges printanière et persister ainsi quelques temps avant de se décomposer. Commun
Comestibilité: Comestible.
2.6.2.2.3 SOUS-CLASSE DES AUTOBASIDIES
Ce sont les " champignons" du langage courant, mais dont le profane ne retient, en fait, que l'élément extérieur, c'est à dire le carpophore.
La plupart des autobasidiés sont saprophytes, ou parasites des parties ligneuses des plantes. Beaucoup d'entre eux vivent en symbiose avec les arbres ( et les végétaux supérieurs ) : leur mycelium peut constituer des des associations appelées mycorhises avec les racines. Ces associations sont à bénéfice réciproque ( voir la rubrique pédologie ).
une majorité de nos plantes forestières forment des mycorhises avec certaines espèces de champignons; Beaucoup de plantes herbacées liées aux mycorhyses, certaines le sont obligatoirement ( les Orchidées doivent être associées aux champignons du genre Rhizoctonia ). On admet de plus en plus que les associations nutritives plantes-microflore apportent à l'hôte, non seulement des matières alimentaires, mais aussi des facteurs de croissance et de résistance au parasitisme animal et végétal. En d'autres termes une mycorhisation abondante serait un moyen de lutte préventif à l'égard de certaines maladies.
Parmi les Autobasidiés, on distingue :
- les Hymnomycètes : champignons à hyménium nu, étalé à l'air libre.
Exemple : Agaric, champignon blanc de prairie, le plus connu et le plus consommé.
- les Gastéromycètes : l'hyménium est enfermé dans le carpophore ( Exemple : la Vesse de loup ou Lycoperdum).
A . Les Hyménomycètes
Souvent saprophytes, vivant sur matière organique abondante ( litière, praiie, bois mort...)
Au printemps une basidiospore germe ---> mycelium qui prolifère tout l'été; vers l'automne apparaît un carpophore sur le feutrage mycélien qui provient en réalité de deux basidiospores n de polarité sexuelle différente ( = hétérothallie )
La plus grande partie du mycelium est à dicaryons (plasmogamie avec deux noyaux : n+ et n- ) . La fusion des cytoplasmes et la formation de dicaryons se réalise très tôt ; le mycélium des Basidiomycètes est constitué dans sa quasi-entièreté par des dicaryons ( ce qui le distingue des Ascomycètes ou la dicaryophase se limite à l'intérieur du carpophore ).
Un fait important à signaler dans le développement du mycélium de Basidiomycètes est la division des articles binucléés par voie d'anses d'anastomoses.
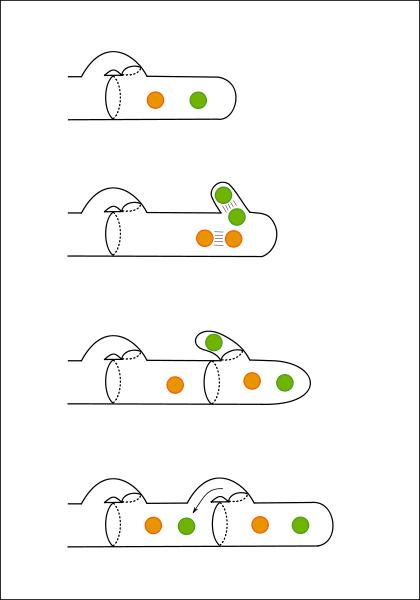
figure Wikipedia : anses d'anastomose
Le carpophore se compose d'un pied ou stipe, d'un chapeau, d'un anneau, parfois d'un voile.
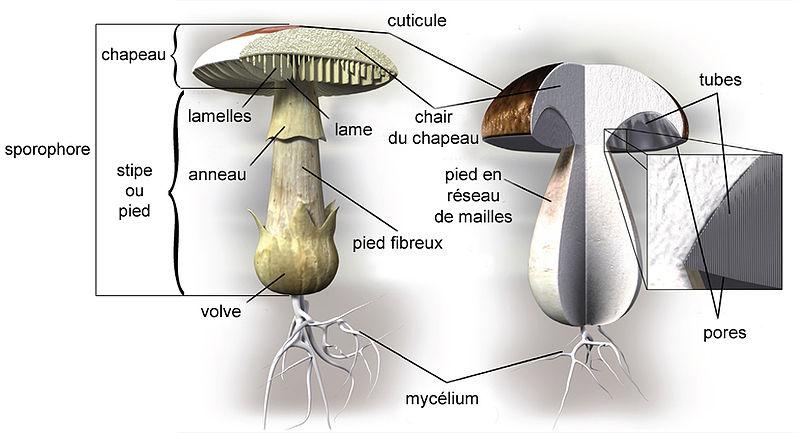
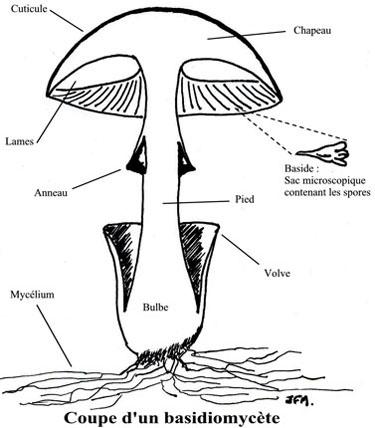
Pas de croissance polarisée par une initiale. Pourquoi se redressent-il ? C'est dans la baside que se réalise la fusion nucléaire ( zygote ) puis la réduction chromosomique
DIFFERENCES GENERALES ENTRE ASCOMYCETES ET BASIDIOMYCETES
| ASCOMYCETES | BASIDIOMYCETES |
|
présence d'organes sexuels ( anthéridies et oogones) à la base du carpophore |
Plus d'organes sexuels : une cellule indifférenciée d'un hyphe n fait office d'un hyphe + et d'un hyphe - |
| dicaryophase brève : réduite aux hyphes ascogènes ) |
la dicaryophase concerne la majeure partie du mycelium et tous les carpophores |
| Deux sexes ( spores + et spores - ) |
souvent 4 sexes (= tétrapolarité sexuelle) : les 4 basidiospores d'une même baside ne sont fécondables qu'avec les éléments d'une autre baside |
|
"anse de panier" au niveau de la cellule mère au niveau de la cellule-mère de l'asque |
anses d'anastomoses partout dans le mycelium |
| la sporée est extérieure au carpophore ( par projection ) | La sporée est intérieure au carpophore |
| les filaments mycéliens ne sont jamais binucléés | les filaments mycéliens sont binucléés dans leur immense majorité |
IMPORTANCE DE CE GROUPE DE CHAMPIGNONS
1) Nombre d'entre eux sont des champignons dont le carpophore est utilisé commealiment. Mais il faut distinguer les champignons comestibles (Agaric, Girolle), les champignons indigestes ( certaines Russules, Bolets, Lactaires ) et lesn champignons toxiques ( certaines Amanites, dont A. banche, A . phalloïde, A. vireuse sont mortelles ).
La mort est due à une destruction des globules rouges du sang .
2) Beaucoup de Basidiomycètes ( et d'Ascomycètes ) sont des saprophytes , ils débarrassent la surface du sol de nombeux déchets organiques qui s'y accumulent. A cet égard, les champignons jouent un rôle important dans les chaînes trophiques. De nombreux noms doivent être cités : Bolets, Fomes,Merulius ( le mérule des bois ).
3) Il existe aussi , parmi les Basidiomycètes, de nombreux parasites ( Rouilles, Charbons, caries, Armillaire, Polyphores, Pleurottes, Coprins ,... )
4 ) Il faut encore citer de nombreux basidiomycètes comme mycorhises .
5) A la limite, il est évident que certains champignons peuvent être classés comme parasites ou saprophytes.
B. Les gastéromycètes
Hyménium enfermé dans son carpophore.
Exemple, Lycoperdum ou saprophytes .
LES FUNGI IMPERFECTI
Ce sont les champignons dits " imparfaits", encore appelé Adélomycètes, Deutéromycètes ou encore Hyphomycètes .
Champignons avec un mycélium cloisonné, mais dont les fructifications ne sont pas connues.C'est donc par ignorance qu'ils sont qualifiés d'imparfaits et beaucoup d'Ascomycètes et de Basidiomycètes étaient jadis rangés dans ce groupe résiduel.
Leur reproduction se fait par des conidies sur conididiophores. Ceux-çi sont soit associés dans des conceptacles globuleux appelés pyenides ou étalés en coussinets appelés acervules ou encore sont indépendants. Trés nombreux genres ( +/- 1000) et plusieurs milliers d'espèces.
Plusieurs classes : Mycelia Sterilia, Coelomycètes, Hyphomycètes, Blastomycètes
Le sol constitue un réservoir inépuisable de champignons imparfaits. Certaines espèces sont de véritables champignons saprophytes du sol, où ils accomplissent tout leur cycle de développement ; ils jouent un rôle important dans la décomposition des débris organiques. D'autres sont des parasites facultatifs des plantes supérieures, susceptibles de végéter pour un temps dans le sol ; ils constituent un foyer d'infection pour les jeunes pousses, et imposent la pratique de l'assolement. D'autres enfin sont des champignons des litières, localisés dans la couche humique du sol ; on y rencontre en particulier des espèces prédatrices (Arthrobotrys, Dactylaria), remarquablement adaptées à la capture des nématodes ou des rotifères. La mycoflore du sol est le siège de compétitions et d'antagonismes, manifestés par la sécrétion de toxines ou par le mycoparasitisme, qui tendent à l'éviction des espèces parasites et contribuent à l'équilibre de la biocénose.
Beaucoup de spores de champignons se retrouvent, à l'état viable, dans l'atmosphère ; elles peuvent être à l'origine de réactions allergiques.
Les champignons du sol et de l'air sont les principales sources de contamination par les moisissures ; les dégradations causées par ces micromycètes aux biens de consommation, produits alimentaires, matériaux et objets fabriqués font subir de graves préjudices à l'économie.
Beaucoup de champignons parasites des plantes cultivées appartiennent aux diverses classes de Fungi imperfecti : Sphæropsidales (septorioses, rot blanc de la vigne) ; Mélanconiales (anthracnoses ou pourritures sèches des fruits et légumes, maladie criblée des arbres fruitiers) ; Hyphomycétales (cercosporioses, alternarioses, fusarioses, et de nombreux parasites secondaires ou de faiblesse). Les parasites appartenant à d'autres groupes (Ascomycètes) ont souvent une phase conidienne très active qui assure la dispersion locale du champignon pendant la période de végétation.
Classification des Fungi Imperfecti :  adelomycetes03.pdf
adelomycetes03.pdf
Date de dernière mise à jour : 05/07/2021
Ajouter un commentaire